










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.112 no.7-8 Pretoria 2016
http://dx.doi.org/10.17159/sajs.2016/20150480
RESEARCH ARTICLE
Diverse diets of the Mio-Pliocene carnivorans of Langebaanweg, South Africa
Adam Hartstone-RoseI, II; Katheryne N. BrownIII; Carissa L. LeischnerI, II; Ka'la D. DraytonIV
IDepartment of Cell Biology and Anatomy, University of South Carolina School of Medicine, University of South Carolina, Columbia, South Carolina, USA
IIDepartment of Anthropology, University of South Carolina, Columbia, South Carolina, USA
IIIMedical Degree Program, University of South Carolina School of Medicine, Greenville, South Carolina, USA
IVHealth Information Technology, University of South Carolina, Columbia, South Carolina, USA
ABSTRACT
The Mio-Pliocene guild of carnivorans of Langebaanweg (LBW), South Africa, is phylogenetically and ecologically diverse. Unlike modern African fauna, this fossil sample contains a large ursid; although there are mustelids, herpestids and viverrids in Africa today, some of the LBW members of those families were much larger than their modern confamilials. There were also numerous felid species, including some that possess a more sabretoothed dental morphology, as well as several species of hyaenids that were very different from their modern confamilials. Questions remain about the dietary morphospace occupied by these fossils. Which taxa were predominately durophagous and which were the most hypercarnivorous? Did the level of durophagy and hypercarnivory in the LBW taxa reach the level of specialisation found in modern carnivores? In the current study, we evaluate the dietary specialisations of all the large terrestrial LBW carnivorans through analysis of the radii-of-curvature and intercuspid notches present in the mandibular dentition. We found that the LBW carnivorans had less sharp premolars than do their modern confamilials -an indication of greater durophagy. However, some families contain individuals with more extreme intercuspid notch patterns, indicating greater hypercarnivory. The ursid also possessed a suite of morphology unlike any modern carnivoran, exhibiting some morphology conducive to durophagy and some that places it functionally among the most hypercarnivorous of modern carnivorans. Thus it was likely capable of consuming high levels of both flesh and bone.
Keywords: Carnivora; Agriotherium; Hayeanidae; Felidae; palaeodiet reconstruction
Introduction
Members of the order Carnivora exhibit a wide range of dietary adaptation and specialisation. Some members are 'hypercarnivorous', specialising in the consumption of vertebrate flesh1-4 (e.g. felids), while others are 'durophagous', specialising in the consumption of bone or other obdurate foods1-4 (e.g. most hyaenids), insectivorous (e.g. the aardwolf, Proteles), frugivorous (e.g. the binturong and kinkajou, Arctictis and Potos), and even strictly folivorous (e.g. the bamboo-eating red and giant pandas, Ailurus and Ailuropoda). Most taxa in the order represent a mixture of several of these specialisations. As carnivorans have diversified, their craniodental morphologies have adapted to diverse dietary niches. Important dietary information can be gained from analysing carnivoran teeth, especially in relation to tearing, shearing, cracking or grinding habits during food processing. These morphological changes and their significance can be evaluated by analysing the craniodental adaptations, and correlating these adaptions with dietary categories (e.g. durophagous or hypercarnivorous).
Analyses of premolar radii-of-curvature (ROC) and intercuspid notches (ICNs) show that tooth geometries correlate with today's observed diets of modern carnivorans; more hypercarnivorous taxa have sharper (higher ROC scores) teeth that are comparatively more intricately notched (higher ICN scores) than durophagous taxa.1-3 The functional inferences are clear: sharper teeth are best for slicing ductile flesh, and yet this sharp edge is vulnerable to damage by particularly hard or tough foods. Likewise, numerous and sharp ICNs function to increase the linear sharp crests available for flesh slicing and also help focus slicing forces for ductile foods toward pinch points like the diamond-shaped slicing notches found in wire cutters. However, the well-developed, small and unsupported accessory cusps would be particularly vulnerable to breakage against hard foods like bone.
Goal and objective
The overarching goal of this study was to expand on the assessment of LBW hyaenid diets3 to compare the morphology of the members of the terrestrial LBW carnivoran guild to modern carnivorans using analyses of dental ICN2 and ROC1 with the objective of evaluating the dietary niche space occupied by the large LBW carnivorans relative to that occupied by modern carnivoran lineages. Although both the geological context and taxonomic systematics of the LBW carnivorans are discussed more thoroughly in the previous literature (see below), we briefly discuss these in order to contextualise our fundamentally morphological study with particular emphasis on visually displaying some of the impressive specimens from this important location.
Background
Langebaanweg - a brief context
Originally discovered during phosphate mining operations in the early half of the 20th century, the fossiliferous deposits at LBW are known for their diverse terrestrial and marine taxa that make LBW one of the most significant Mio-Pliocene fossil-bearing sites in Africa. The broader Langebaanweg deposits include four distinct formations in geological succession: Elandsfontyn (underlies the formation in question and rests on local bedrock); Saldanha (early- to mid-Miocene in age); Varswater (most intensively studied Tertiary sequence in southern Africa); and Bredasdorp (post-early Pliocene deposits). The Varswater formation is divided into three distinct Members that comprise the LBW 'E' Quarry - the focal sample of the present study - from where most of the highly studied and best understood Mio-Pliocene fossils in southern Africa have been unearthed.5
The Langeberg Quartzose Sand Member (LQSM) was deposited during the late Miocene and early Pliocene and is abundant in fossils from both vertebrates and invertebrates and is thought to have resulted from repeated sedimentation events and the marine transgression of the Berg River.6 Hundreds of thousands of fossils have thus far been recovered from 'E' Quarry belonging to over 200 species, more than 80 of which are mammals.5 The remains of 13 out of 14 mammalian orders (excluding Sirenia) are represented, some of which no longer have relatives living south of the Sahara.7
The LBW 'E' Quarry is known for its fossils that have been found nowhere else in southwest Africa in such unprecedented quantity and quality.5 Fossils from this deposit reflect what has been interpreted as a deteriorating environment during the early Pliocene as well as its subsequent change to fynbos and open grasslands that can currently be seen in LBW.7 Within these two deposits, the following carnivoran families have been identified: Felidae (cats), Viverridae (civets), Hyaenidae (hyenas), Herpestidae (mongooses), Canidae (dogs), Ursidae (bears), Mustelidae (weasels) and Phocidae (seals).5,7-25
Carnivorans of Langebaanweg
'E' Quarry is especially well known for its fossil carnivorans, which comprise one third of the mammals from the locality and make Carnivora the most diverse order within the assemblage.5 The fact that carnivorans are generally poorly represented at other late Miocene and early Pliocene sites in Africa makes the 'E' Quarry assemblage that much more valuable.7 Unique in such high numbers to this fossil site are the remains of at least 14 individuals of the giant extinct African bear Agriotherium africanum, which are not generally found together in such large numbers.20,26,27 This bear and many other large carnivores found at LBW (e.g. Ikelohyaena, Hyaenictitherium, Plesiogulo and Dinofelis) originally came from Eurasia when the drop in sea level at the end of the Miocene allowed passage into Africa via the Mediterranean land bridge.7The subsequent rise of global sea levels at the beginning of the Pliocene drastically changed the climate in LBW from tropical to temperate - thus, the 'E' Quarry assemblage dates from a period when fauna were either adapting to changing environmental conditions or going extinct.7
Ursidae
Agriotherium was the most geographically dispersed member of the family Ursidae, with fossils found at sites in North America, Europe, Asia and Africa.28 It was the only ursid known to have inhabited sub-Saharan Africa.28Agriotherium africanum, the species found in LBW, was very large (~540 kg29) and exhibited several relatively unique craniodental characteristics: a short-snouted, robust skull; a 'chin'-like protrusion at the symphyseal region of the mandible; well-developed sectorial carnassials; and a premasseteric fossa.5,20,27,29,30 The only extant ursid to also have premasseteric fossae is Tremarctos, which also has sectorial carnassials reminiscent of Agriotherium. This has led to the hypothesis that Agriotherium may have given rise to the Tremarctos lineage and possibly the other extant ursids.29Agriotherium may have been outcompeted and replaced by modern bears.28 Ursidae is a fairly young family, and likely evolved from caniform ancestors only 23-24 million years ago, and Agriotherium exhibited many craniodental characteristics that are reminiscent of these ancestors (such as a robust P4) which has caused controversy over whether or not Agriotherium should be considered an ursid or an amphicyonid.27,30
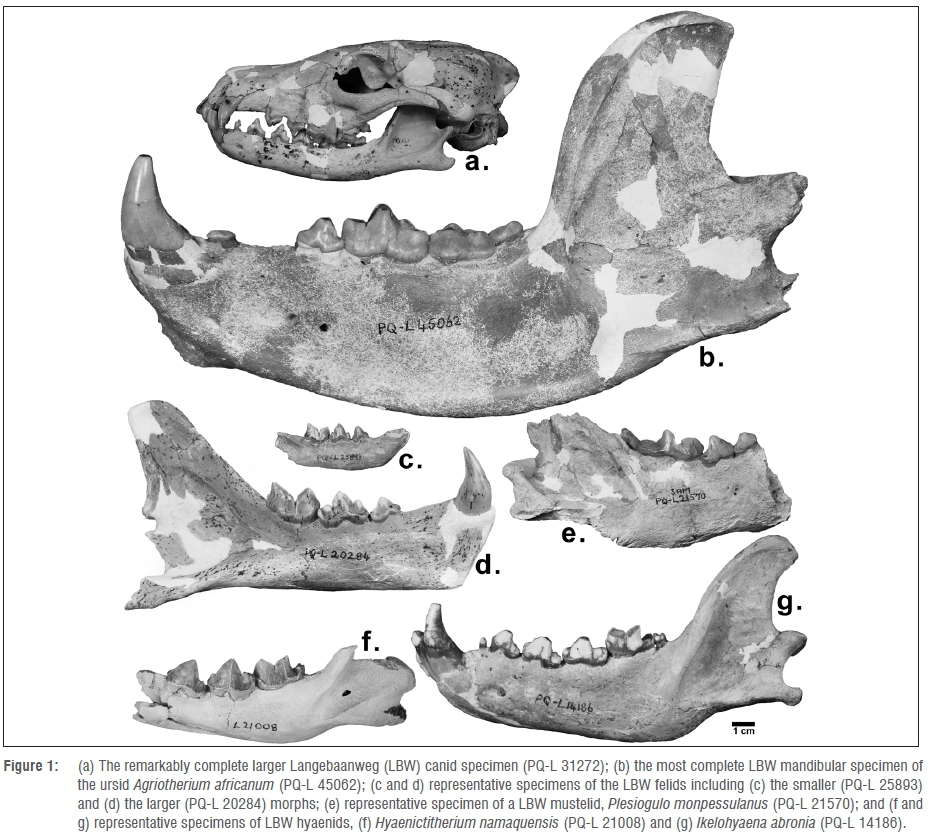
Previous research on Agriotherium has focused largely on estimating its bite force in relation to body mass for the purposes of reconstructing evolutionary anatomical adaptations and hypothetical feeding ecology. For instance, one study30 concluded that Agriotherium had an absolute bite force higher than that of any other mammalian carnivore and that, even relative to body mass, Agriotherium's bite force was still formidable. However, the wide range of feeding niches and masticatory adaptations occupied by modern ursids make it difficult to predict Agriotherium's diet. For example, the giant panda is able to generate and sustain a powerful bite force comparable to Agriotherium in order to exploit bamboo, its primary food source, although some studies have claimed that Agriotherium actively hunted and consumed large vertebrates.20,30 Other studies29have suggested that the diets of these ursids included a lot of vegetation, while still others29,31 have argued that Agriotherium is adapted largely for durophagy. Because of the presence of a true carnassial complex and robust molars fit for durophagy (Figure 1b), the growing consensus seems to be that Agriotherium was a predator-scavenger.32
Canidae
Canids are fairly poorly represented in the LBW assemblage, as they were not widespread in southern Africa during the Mio-Pliocene. There are at least two genera of canids: a larger one represented by a remarkably complete (although fragile) specimen (Figure 1a) that has been ascribed to the genus Eucyon, which is the earliest known canid from Africa; and a smaller, fox-like sample tentatively ascribed to the genus Vulpes.5,7,15,33Craniodental characteristics unique to the Vulpes specimen include short development of the P4 protocone and the presence of a second distal p4 cuspid.27,34 A third specimen may also represent an additional species of canid, but the inability to distinguish the lower dental elements from that of a viverrid (a morphologically and taxonomically diverse group in the LBW sample) have prevented a confident diagnosis7 and these specimens were not complete enough for our analyses anyway.
Felidae
The taxonomy and systematics of the LBW felids have yet to be fully resolved. The majority of the literature on the LBW felids focuses on the larger specimens ascribed to the machairodont taxon Dinofelis (Figure 1d).3,35-38 Less complete remains of a second large sabretoothed cat, ascribed to Homotherium, have also been found at LBW.7 There also are smaller felids in the sample - most of which have been questionably assigned to the modern genus Felis (Figure 1c), namely F. obscura and F. issiodorensis (both of which are lynx-like), 'Felis sp.', which resembles a wildcat, and a felid of unknown genus and species.7,16 As members of Felidae are highly specific to particular environments, a more complete systematic and functional understanding of the LBW species would help us understand the diversification and niche partitioning of this African lineage from the Quaternary.6
The wide distribution of Dinofelis and its especially broad presence (both in number of localities and specimens at those localities) in Africa suggest that it could be the most common fossil felid in the African Neogene.27,38 The specimens from LBW represent the oldest known individuals of Dinofelis15and have been ascribed to the taxon D. cf. diastemata27which differs from its Eurasian congeners by a smaller body size and the presence of craniodental characteristics similar to those of the non-sabretooth genus Panthera in that it possessed a short upper canine, a P3 less reduced than in other machairodonts, and a less elongated m1 than those found in other machairodonts.38-40 A notable evolutionary change that has been observed in LBW Dinofelis is a lengthening of the carnassials in relation to the rest of the tooth row - indicative of evolution from the more primitive state toward the more machairodont traits that are generally observed in other sabretoothed cats.38 Like modern felids, Dinofelis was almost certainly hypercarnivorous, although it possessed a reduced post-canine dentition in comparison to other members of Felidae to accommodate the maxillary canines.2 The rise of felids belonging to the genus Panthera likely led to the decline of Dinofelis and its eventual extinction.7
Hyaenidae
Members of Hyaenidae are the most commonly found terrestrial carnivores in the LBW assemblage. At least five species of hyena (Chasmaporthetes australis, Hyaenictitherium namaquensis, Hyaenictis hendeyi, Ikelo-hyaena abronia, 'Hyaenidae sp. E'41; Figure 1f-g) are represented in these assemblages that coexisted during the late Miocene and early Pliocene in southern Africa.3,7,14,18,27 These species are believed to have evolved from civet-like ancestors on more than one occasion during the early Miocene.3,18,41 Our recent study3, which utilised the same methods as our current study, found that the LBW hyaenids were poorly adapted for bone cracking and were better suited for a more generalised carnivoran diet than the highly derived extant durophagous hyaenids. This finding corresponds with the conclusion that modern levels of durophagy are known to have appeared only recently in the fossil record.42
The ability of durophagous hyaenids to efficiently process and consume obdurate foods like bone is enabled by the presence of broad, stout premolars that can transmit large forces without breaking.1-3 However, gnaw marks and acid etching on the fossilised bones of prey species (consistent with being partially digested by hyenas) have been found at LBW, indicating that at least some of these extinct species engaged in the same behaviours and occupied part of the same scavenging niche as most of their extant confamilials.7,43
Informally known as the 'hunting hyena', Chasmaporthetes australis was the largest, yet most gracile of the LBW hyaenids and was likely adapted to a more cursorial lifestyle.15 Craniodental characteristics of this species include: smaller cheek teeth and a markedly weaker masticatory apparatus when compared to the extant durophagous hyaenids (Crocuta, Hyaena and Parahyaena); strong posterior accessory cusps on p2 and p3; a large P4 protocone; a very large anterior p4 accessory cusp; and an ml with a single, blade-like talonid cusp.15,27,44 Analysis of the premolar ICN and occlusal ROC scores of this species suggests that Chasmaporthetes australis was more hypercarnivorous than modern hyenas and may have had a diet similar to those of modern felids.1-3
The holotype of Ikelohyaena abronia represents the most complete skeleton found to date within the LBW 'E' Quarry and is comprised of the skull and more than 80 postcranial bones from the same individual.7 In a study by Werdelin and Solounias41, Ictitherium preforfex was synonymised with I. abronia - these were originally thought to be two separate species because of their large variation in size.15,18 Craniodental characteristics unique to I. abronia include an enlarged p3 and P3, a lack of anterior accessory cusps on p2 and P2, a large and roughly triangular-shaped m1, and smaller, lower crowned anterior premolars than H. hyaena.15,27 Ikelohyaena abronia was the most durophagous of all LBW hyaenids3 although it lacked the strong bite force of the modern durophagous hyenas.3,45 Along with the not yet formally named Hyaenid sp. E from LBW, I. abronia belonged to a clade of early or transitional bone-cracking hyaenids.46
Hyaenictitherium namaquensis was the most widespread African member of the ictitherines, a clade of hyenas with canid characteristics that was most prominent in Eurasia from the middle of the Miocene until its end.27 Like other ictitherines, the genus Hyaenictitherium possessed a large m1 talonid as well as both an m2 and M2.27,41,47Unlike its more doglike Eurasian counterparts, however, African species within Hyaenictitherium possessed crushing dentition and shorter, more massive canines along with a stronger mandibular ramus - all adaptations conducive to consuming obdurate foods.27 Excluding many dental characteristics, the skull of H. namaquensis shares many similarities with the sympatric I. abronia and the extant Hyaena hyaena.15Craniodental characteristics specific to this species include: a marked shortening of the snout; a significantly larger M1 than that of both I. abronia and H. hyaena; and higher crowned premolars and molars than I. abronia.15
Hyaenictis hendeyi, formerly described as Euryboas sp. by Hendey15, was a gracile, long-limbed LBW hyaenid that was believed to be both cursorial and actively predacious15,44, much like C. australis. This species retained an m2 (a primitive character in hyenas), small or non-existent anterior accessory cuspids on p2 and p3, and a distinct anterior accessory cuspid on p4 that is notably smaller than its posterior accessory cusp.44 Because of the absence of extant lineages of big cats at the time when H. hendeyi lived, it has been proposed that this species could have occupied the ecological niche that is now occupied by the extant cheetah Acinonyx.7
Only a single mandibular fragment of Hyaenid sp. E with an intact p2 and p3 from an immature individual has been discovered and described.15,18Although characteristics of the dental elements of this specimen indicate that it is distinct from all known hyaenids, there is not enough material present to completely name and describe it as a species.41 Hyaenid sp. E displays the following dental characteristics: an absent p1; no noticeable anterior accessory cusp on p2; and a p3 with no anterior accessory cusp and a greatly reduced posterior accessory cusp.15 These characteristics, at the least, suggest that Hyaenid sp. E was highly durophagous and belonged to the same clade of early/transitional bone-cracking hyaenids as I. abronia.3,46 Because this specimen is rather fragile and subadult, it was excluded from our sample.
Although we have previously published these analyses for the LBW hyaenid sample3, we are including them in the present investigation to be able to more thoroughly discuss the morphospace occupied by all of the LBW large carnivore guild. As described in our previous study, the only LBW hyaenid specimens complete enough for our analyses were from Chasmaporthetes and Ikelohyaena.
Mustelidae
Although mustelids were widespread throughout Africa and the northern continents during the late Tertiary, they are relatively poorly represented at 'E' Quarry (in terms of numbers of species and overall specimens) in comparison to the species diversity and abundance of other families.19Three species of large mustelid have thus far been identified from LBW 'E' Quarry: Plesiogulo monpessulanus, Mellivora benfieldi and Sivaonyx hendeyi.15,19,48The discovery of the remains of at least three individual P. monpessulanus (Figure 1e) at LBW marks the southernmost known record of a wolverine.15,19 As members of its genus were widely spread throughout Eurasia and North America during the late Miocene and Pliocene, it has been suggested that Plesiogulo could also be found throughout the African continent.19,23 It is generally accepted that P monpessulanus is ancestral to the extant Gulo gulo, as a result of both morphological similarities and assumed behavioural similarities.19,23What so obviously sets this species apart from its modern descendant is its large size, apparently being only slightly smaller than the largest recorded fossil mustelid, the North American Megalictis ferox.18Dental characteristics of P. monpessulanus include a relatively long m1 talonid, shorter carnassials relative to those of G. gulo, and larger maxillary and mandibular cheek teeth relative to those of the extant G. gulo.27While modern Gulo is extremely powerful - capable of cracking bones49 - the dental characteristics of P. monpessulanus likely indicate even greater durophagy abilities.
Mellivora benfieldi, likely ancestral to the extant Mellivora capensis, was somewhat smaller in size than its descendant and is tentatively represented by as many as six individuals from the LBW 'E' Quarry. Dentition from these fossil specimens shows a loss of p1 (an advanced character in mellivorines), narrow premolars, and sharp-pointed cusps and sharp-edged keels on the cheek teeth compared to those of M. capensis. Mellivora benfieldi is the less specialised of the two species in terms of the lack of defined p4 accessory cusps and the presence of a comparatively unspecialised m1 talonid.19
The Mio-Pliocene otter Sivaonyx hendeyi of 'E' Quarry was removed from its original classification because its unique m1 presents characteristics too different to warrant inclusion within the pre-existing species Enhydriodon africanus.48It is believed to be an intermediate between E. iluecai (late Miocene) and E. sivalensis (late Pliocene or early Pleistocene) in terms of its craniodental characteristics as well as its temporal occupation.19Sivaonyx hendeyi was slightly larger than the extant Aonyx cinerea in terms of body size, and its cheek teeth exhibit a primitive inflation of cusps and lowering of crown height that is much more developed in extant members of Enhydra.19
Herpestidae
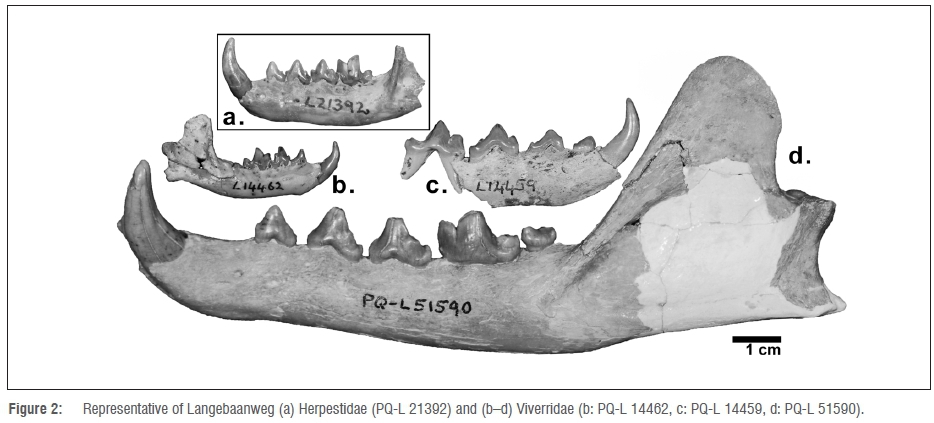
Herpestids are relatively common in LBW 'E' Quarry (Figure 2a); however, the fragmentary nature and similarity among specimens makes classifying them to a species level difficult.15 Two unidentified or undescribed species of Pliocene Herpestes (H. sp. A and H. sp. B), have been described from LBW. Herpestes Species A lacks an anterior accessory cusp on P3 and displays a P1 that is triangular in outline. Herpestes Species B was smaller in size than Species A, and lacks the first mandibular premolar. Herpestes ichneumon, H. pulverulentus, and Suricata major have been identified from 'E' Quarry. Herpestes ichneumon has been hypothesised to be ancestral to the modern Egyptian mongoose and displays many similar characteristics in its mandibular dentition, but also has a larger m2 than its modern conspecifics. Herpestes pulverulentus has been hypothesised to be ancestral to the extant Cape grey mongoose (Galerella pulverulenta) and is represented by a skull with incomplete maxillary dentition and a mandibular fragment. As in H. Species B and the modern G. pulverulus, H. pulverulentus lacks p1; it was also likely somewhat larger than its modern conspecific. Suricata major was of large size and had a p4 without an anterior accessory cusp and also displayed a p1.15
Viverridae
One of the more diverse families of carnivorans represented in the LBW sample is the Viverridae15 (Figure 2b-d). However, most of the smaller specimens have not been classified beyond the level of family.27 Of the identified LBW specimens, one was the giant viverrid Viverra leakeyi50(Figure 2d). Judging from one nearly complete LBW cranium, V. leakeyi had both a high and narrow skull as well as a relatively long mandible.15The postcranial remains of this species suggest that it was nearly the same size as the extant Civettictis civetta, while its craniodental characteristics share a strong similarity with those of the modern Canis mesomelas.15
Genetta sp. is represented by a single mandibular fragment from 'E' Quarry and appears to have been about one third of the size of extant genets. Despite the disparity in size, the dentition of this specimen is very similar to that of Genetta tigrina in that its p2 has one posterior accessory cusp, while both the p3 and p4 have two. The mandibular fragment also displays a convex curve along the entire length of its inferior margin. It is the only known fossil genet to have been described in South Africa, but the lack of additional material has prevented it from being named.15
Materials and methods
Methods
Dietary niche is assessed using two previously developed methods: premolar ROC and ICN score analyses.1-3 Both methods examine dietary correlates of mandibular dentition. Although many studies of dietary correlates have focused on lower dentition (most notably the primate work of Kay and followers thereof51-53), this focus on only maxillary dentition limits dietary reconstruction of fossils - a constraint to be remedied in further studies. Furthermore, we studied only adult specimens with relatively unworn and taphonomically unmodified crowns that are strong enough for physical moulding. (See the Discussion for notes about the inclusion of maxillary dentition and fragile specimens.)
As in our previous studies1-3, modern carnivorans are ascribed to general dietary categories based on the primary literature and previous categorisation schemes54. The first category consists of 'hypercarnivores', or species whose diet consists of more than 70% vertebrate flesh. The second category consists of 'durophages', or species that consume bone or other hard dietary products. The last two categories are 'meat/ non-vertebrate' and 'non-vertebrate/meat', which contain species that consume fruit and/or insects as either secondary or predominant components of their diets, respectively.54
Premolar radii-of-curvature
Premolar ROC is a dietary evaluation method used to predict diet and methods of consumption of flesh or bone. Post-canine specimens containing p3, p4 and the carnassial (m1) cusps are moulded using regular-body President Jet (Coltene Whaledent, Cuyahoga Falls, OH, USA), a quick-setting product used primarily in the dental industry, and then cast using Smooth-Cast (Smooth-On, Easton, PA, USA), a fast setting, low-viscosity, two-part resin. The casts are sectioned at the central cusp of each premolar and the two mesial cusps of the carnassial using a thin kerf saw in the coronal plane according to their own orientation.
These sections are then scanned onto a flatbed scanner and measured in ImageJ (NIH, Bethesda, MD, USA) by fitting the cross section of the cusp with the smallest circle that fills the tip and then taking the inverse of the radius of the fitted circle (1/radius) as the ROC. Unfortunately, teeth are not simple geometric shapes and fitting a circle to a cusp point is clearly an oversimplification. Other techniques have been employed to fit more complex shapes (e.g. a parabola) to tooth cross sections, but these too are only slightly more accurate in describing the shape. Indeed, each additional term that could be added to the equation would make the line fit the curve of the tooth more accurately. However, the fitting of circles to premolar cross sections has been shown to be reproducible and indicative of diet, and, because it is easier and faster than digitising the whole outline of the tooth and fitting lines with algorithms, more data can be incorporated into the sample more efficiently.1,3
Intercuspid notch
Intercuspid notch scoring is a dietary evaluation method in which the notches between cusps of premolars are scored on a scale from 1 to 5 with sharpness of crests leading into the notch being considered as perpendicular to the slope of the crest itself.2,3 A score of 1 represents a barely visible notch; 2 represents a notch between two rounded, dull crests; 3 represents a notch between one dull crest and one sharp crest at <45°; 4 represents a notch between two sharp crests; and 5 represents a 'carnassiform notch' 2,3 or a notch with a keyhole pattern formed at the bottom of a sharp vertical notch at the convergence of crests.
Interpretation of metrics
For both ROC and ICN, modern species that have higher values tend toward hypercarnivory and modern species with lower values tend toward durophagy.1-3 The p3 is most indicative of diet for ROC analysis1 because the carnassial is generally utilised for meat slicing in large carnivores and therefore the greatest level of durophagy is restricted away from this fragile tooth - especially in feloids. In some canoids, durophagy is done in the posterior-most portions of the tooth row - the carnassial talonid and posterior molars. However, because these regions are not universal in carnivores, dietary correlation has been restricted to premolars and carnassials. The p4 correlates most closely with diet along the hypercarnivory-durophagy scale for ICN analysis2 because this tooth contains the most variability in ICN and therefore is most discriminant along the continuum.
All statistical significance was evaluated using a non-parametric Wilcoxon test. When specific p-values are not given, p<0.05 is taken to be significant and p<0.01 is taken to be highly significant.
Sample
A total of 264 specimens of 35 modern taxa were included from the collections of the American Museum of Natural History (New York) and the National Museum of Natural History (Washington, DC) - a sample that we fully described in our previous publications.1,2 These were compared to a total of 65 LBW specimens that were complete enough to evaluate our variables of interest at least in part (Table 1).
Unfortunately, our methods were developed to evaluate dietary specialisation in relatively large carnivorans the size of black-backed jackals, Lupullela mesomelas (~8kg55) or larger - essentially focusing on carnivoran specialisations in hypercarnivory, durophagy and moderate to extreme herbivory - to the exclusion of the dependence on insectivory found extensively in small members of the order. Thus, the smallest LBW viverrids and mustelids, the smaller of the two canids, and all of the herpestids are essentially outside of the body size range of applicability of these methods. However, in an effort to address the guild as completely as possible, some individuals below this size range (one LBW herpestid and one small LBW viverrid) were included. Because of this scaling issue, their dietary assessments, as discussed at the end of this paper, should be taken as tentative. Much more work needs to be done on the small LBW carnivorans including extending methods to account for the effects of insectivory on dental sharpness and cusp notch morphology (see future directions).
Results
Family-level radii-of-curvature results
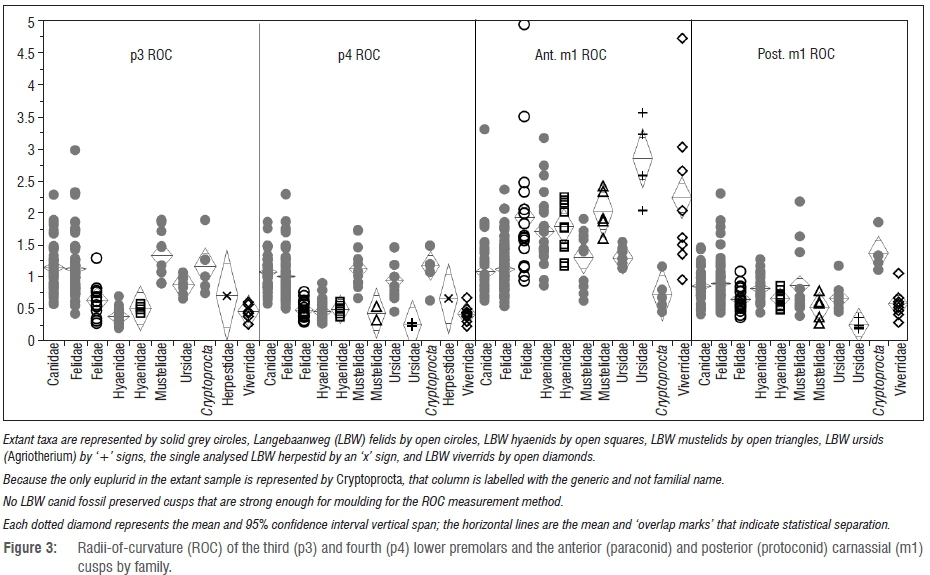
The ROC results for p3, p4 and the anterior and posterior cusps of the carnassial (m1) occupy similar morphospace as those of modern carnivorans (Figure 3). As we found previously3, the modern and fossil hyenas overlap in the sharpness of their teeth. However, comparisons of specific ROC data of the other families reveal significant differences between fossil and modern confamilials. Namely, at the family-level, the LBW felids have lower p3 and p4 ROC (i.e. are duller) than the modern felid sample (p=0.0002 and p<0.0001, respectively). Likewise, the LBW viverrids also have significantly (p=0.0131 for p3 ROC and p<0.0001 for p4 ROC) less-sharp premolars than those of Cryptoprocta, the most comparable extant viverroid in our comparative sample.
Although the LBW ursids (Agriotherium) and mustelids do not preserve third premolars that are complete enough for ROC analysis, the p4 ROC results are interesting; in the ursid comparison, the Agriotherium specimens have significantly less-sharp premolars than their modern confamilials (p=0.0048), and while there are only two LBW mustelids with well-preserved fourth premolars, these two teeth are less-sharp than those of their modern confamilials, and despite the small sample, this difference does approach significance (p=0.0567). Similar patterns exist when considering the sharpness of the posterior cusp (protoconid) of the carnassial; the LBW felids and viverrids have significantly less sharp posterior carnassial cusps than do modern felids and Cryptoprocta (p=0.0239 and p<0.0001, respectively). The fossil mustelids and ursids have duller posterior carnassial cusps than do their modern confamilials, although these differences are not significant at these small sample sizes.
The patterns notably shift when considering the anterior carnassial (paraconid) cusps (Figure 3); while the LBW taxa tend to have duller premolars than their modern confamilials, their posterior carnassial cusps are sharper for all families other than the hyaenids, significantly so for the felids, ursids and non-hyaenid viverroids (p<0.0001 for all three comparisons).
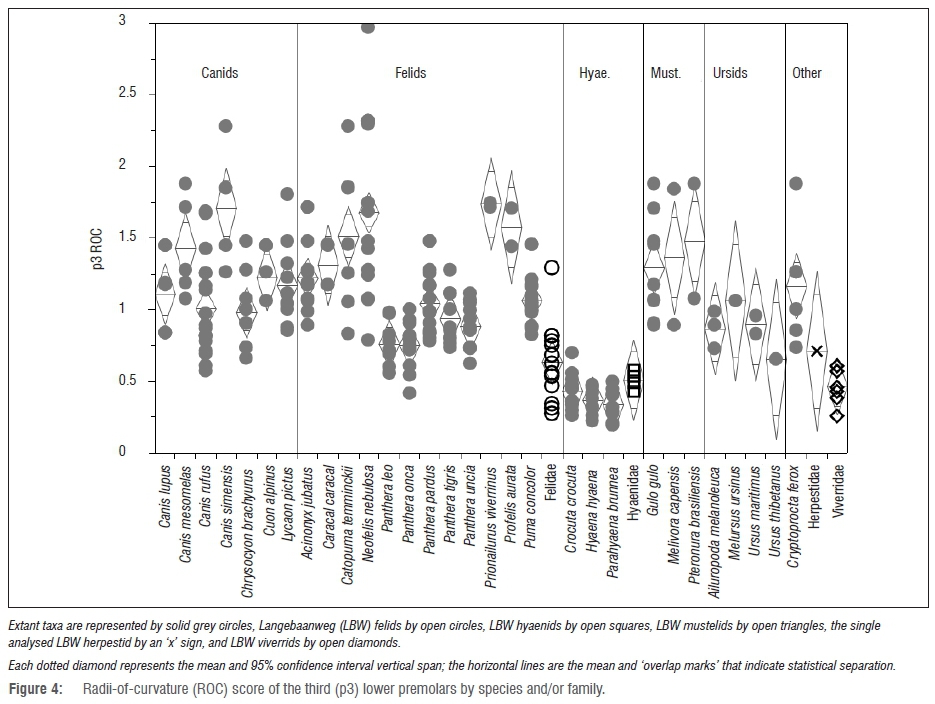
Species-level radii-of-curvature results
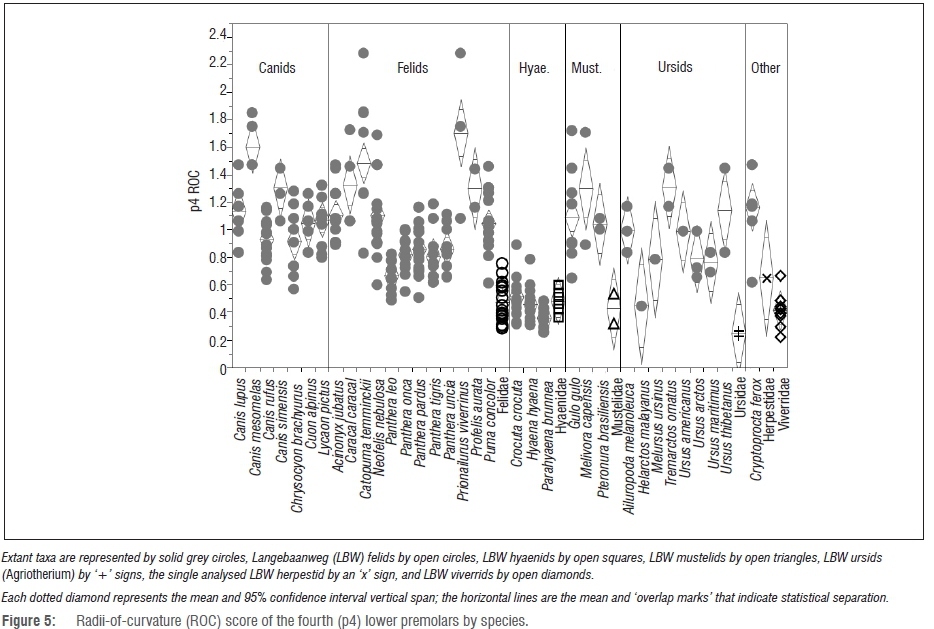
With some ambiguity about the taxonomic diversity within the LBW families, comparing the fossil families to individual modern species yields interesting results. For instance, the LBW felids have significantly less-sharp premolars (Figures 4 and 5) than those of the sharpest felids (e.g. Acinonyx, Caracal, Neofelis, Prionailurus and Profelis) and indeed overlap with all of the modern and fossil hyaenids - a group that overlaps in premolar ROC only with the most robust of the felids (e.g. Panthera leo, P. onca and P. tigris). Likewise, the LBW viverrids have significantly less sharp premolars than those of the modern Cryptoprocta (p=0.0035 and p<0.0001 for p3 and p4 ROC, respectively). When examining the ROC of mustelid and ursid fourth premolars (again, the only tooth well enough preserved in the LBW sample for these families), they are indeed statistically distinct from some of their confamilials despite the small comparative sample size.
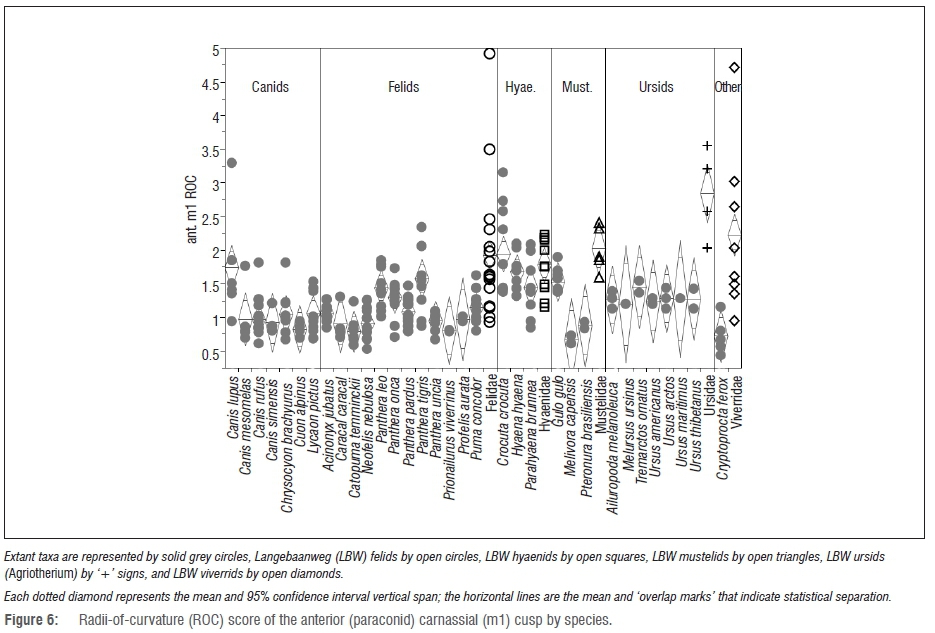
When examining the ROC of the carnassial cusps by species (Figures 6 and 7), as was evident in the family level analysis, the LBW felids have significantly sharper anterior carnassial cusps than all but a few felid species (namely Panthera leo, P. tigris and Profelis aurata). The LBW mustelids have relatively sharper anterior carnassial cusps than their modern congeners, but this find is not statistically significant (probably as a consequence of the small sample sizes). However, despite the extremely small sample sizes of modern ursids, the Agriotherium specimens are significantly sharper in this cusp than those of most of the ursids.
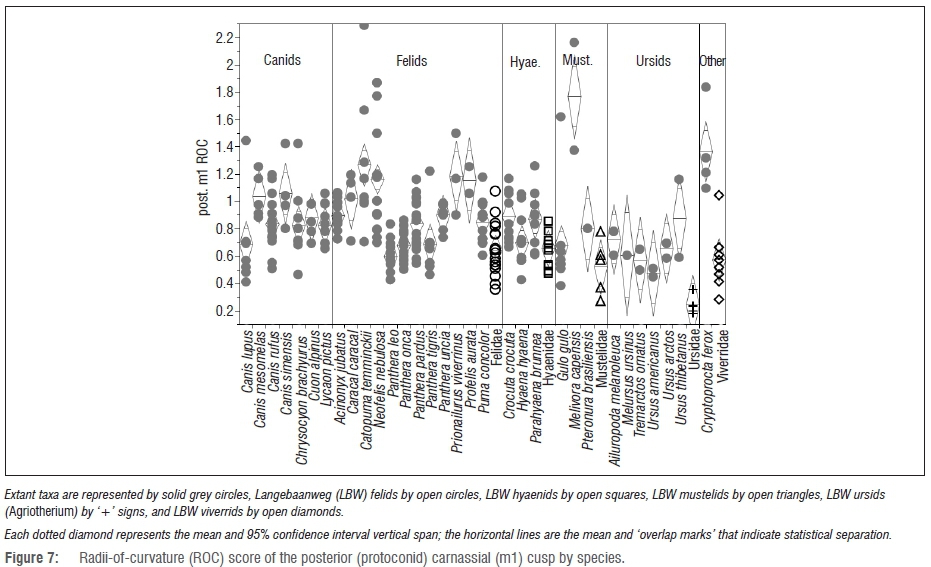
In terms of the posterior carnassial cusp ROC scores (Figure 7), the LBW specimens are statistically indistinguishable from almost all of their modern analogues. There are two notable exceptions: the LBW mustelids have significantly sharper posterior carnassial cusps than do the modern Mellivora, as do the LBW viverrids relative to Cryptoprocta (p>0.0001 for both comparisons).
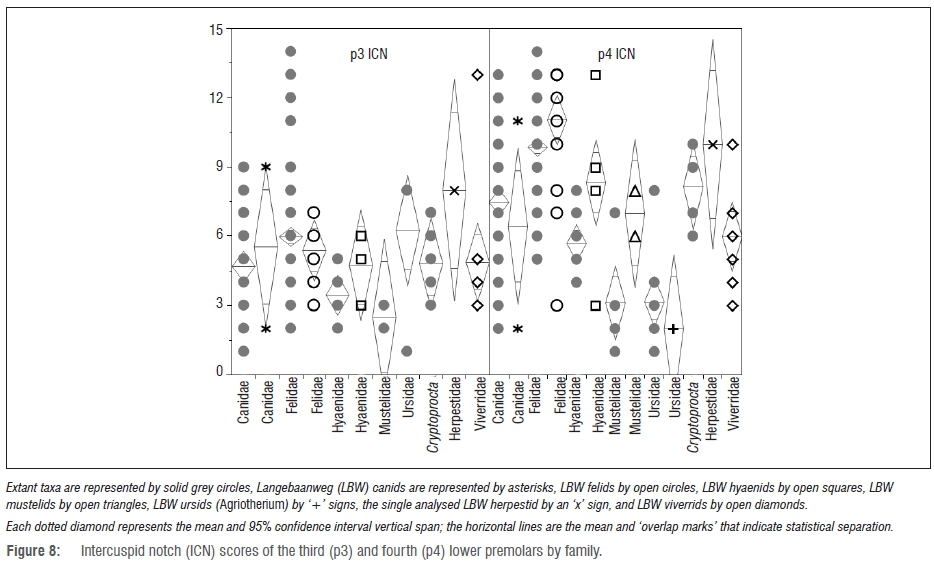
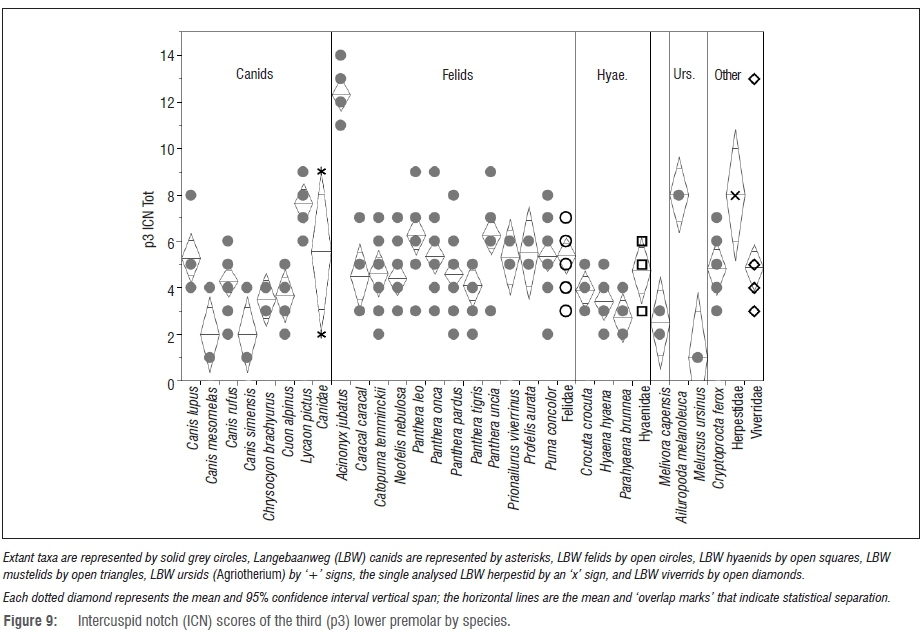
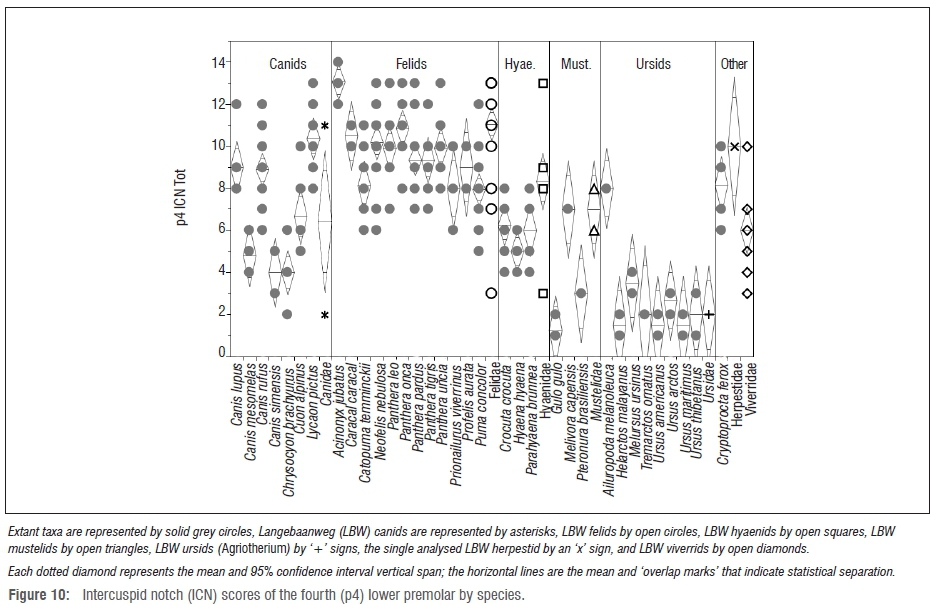
Premolar intercuspid notch results
At the family level, none of the LBW samples can be statistically distinguished from their confamilials (Figure 8). However, comparing the fossil samples to individual modern species does yield significant findings (Figures 9 and 10). Namely, the LBW felids have significantly less notched third premolars than those of Acinonyx (p<0.0001) - an unsurprising finding given that Acinonyx has a significantly higher p3 ICN score than every other carnivoran represented by an adequate sample size.
However, the LBW felids also have significantly lower p3 ICN scores than those of Lycaon (p=0.0434) - the most hypercarnivorous of the canids and a taxon with a p3 ICN score significantly higher than that of about half of the modern felids.
The only two well-represented LBW canids show vastly different mor-phospace with the small specimen (previously ascribed to Vulpes5,7) with premolar notch morphology similar to the most durophagous modern carnivorans, and the larger specimen (previously ascribed to Eucyon5,7) exhibiting notch morphology that would place it among the most hypercarnivorous modern carnivorans. Although the LBW hyaenids have slightly more notched third premolars than their confamilials, they are not significantly so.
As has been found previously1-3 (and discussed above), the p4 ICN scores are actually more telling (Figure 10). By this metric, the LBW felids have significantly higher scores than all but the most hypercarnivorous carnivorans (most of the modern felids, Canis lupus and C. rufus, Lycaon and Cryptoprocta). However, surprisingly (although as discussed previously3), the LBW hyaenids also have significantly higher p4 ICN scores than many modern carnivorans including their confamilial Hyaena, placing them statistically within the range of the more hypercarnivorous lineages (e.g. most of the felids) - and significantly more hypercarnivorous even than some of the canids, e.g. Canis simensis (p=0.0378) and Chrysocyon (p=0.0007) and all of the ursids other than the unique dentition of Ailuropoda. Among the mustelids, the LBW sample has statistically higher p4 ICN scores than those of Gulo (p=0.0439). As seen with the family-level analysis, the two LBW canids have very different p4 notch scores with the larger specimen plotting among the modern Lycaon and most hypercarnivorous felids, and the smaller specimen plotting below even the most durophagous hyaenids and sorting among Gulo and most of the bears (Figure 10). According to its p4 ICN score, Agriotherium falls well within the modern ursid with the exception of the highly derived Ailuropoda - a taxon with highly notched premolars perhaps as an adaptation for severing tough bamboo fibres.56 Despite the small ursid sample size, the difference between the Agriotherium and Ailuropoda p4 ICN scores is nearly significant (p=0.0502) as it is between Ailuropoda and all of the other modern ursids.
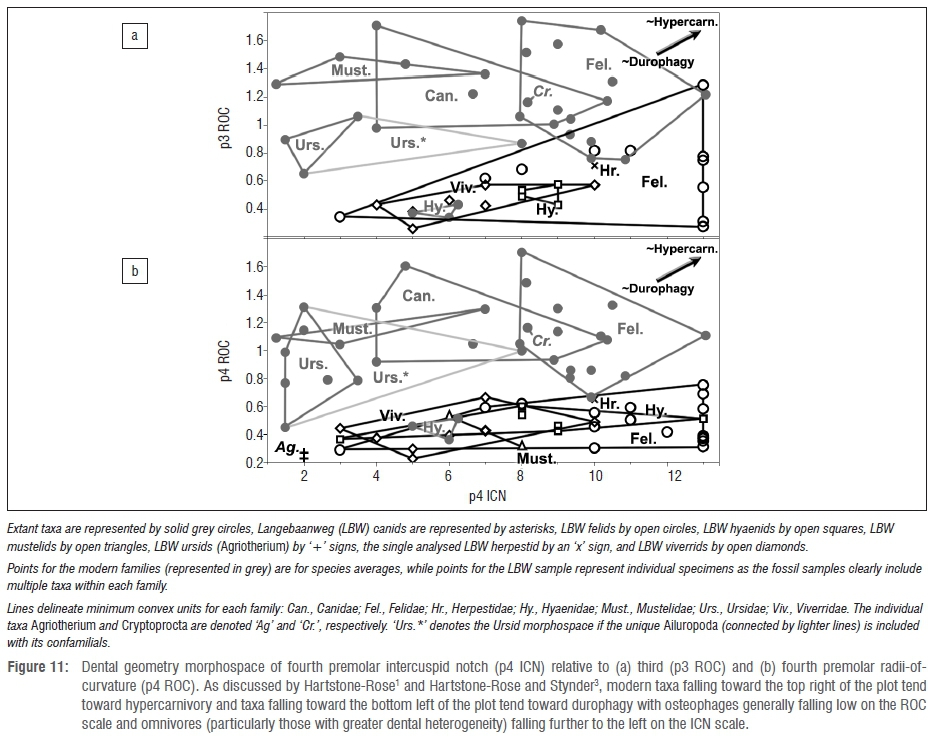
Overall dental geometry morphospace
Comparing the LBW carnivorans using individual measures of ROC and ICN relative to whole samples of modern analogues yields mixed statistical results: by some measures some of the LBW lineages are statistically distinct from their modern analogues, and by some measures they are not. However, comparing the LBW specimens to the morphospace represented by the mean of individual modern species within the families, it is readily apparent that the Mio-Pliocene carnivorans from Langebaanweg had drastically different dental morphology (Figure 11).
Although Figure 11 compares the morphospace of species means of modern carnivorans to all analysable individuals of the fossil sample (i.e. an admittedly unequal comparison), for graphical purposes, it is informative. In this respect, with the notable exceptions of the hyaenids and felids, the LBW carnivores do not overlap with the mean clouds of the modern carnivorans. That is, the Agriotherium specimens, to the extent that this small sample could be analysed, fall outside of the morphospace of the averages of the modern ursid taxa. Likewise, the modern and fossil mustelids are also separate. In both cases, the separation is predominantly in terms of their premolar ROC
- namely the fossils have less sharp fourth premolars than their modern confamilials. The same is true of the LBW herpestids and viverrids. Because the ROC scores of the LBW canids could not be evaluated, they are not on these plots. However, their respective notch scores of 2 and 9 for their third premolars and 2 and 11 for their fourth premolars are more divergent than the species averages within any of the modern families. In other words, had their premolars been preserved enough for evaluation, these two specimens represent an amount of variation more extreme than that found between average individuals of any two modern confamilials in our sample.
The notable difference between the LBW and modern families, the hyaenids, falls along the other scale: the fossil specimens have similar premolar sharpness (ROC) as their modern relatives, but their teeth generally have higher notch scores (ICN; Figure 11a). Although there is overlap in some measures of morphospace (as exemplified by Figure 11b), the ICN difference is notable. Namely, this measure documents the shift that occurred in the lineage: a transformation from the highly cusped feliform ancestral hyaenids to the most durophagous modern carnivorans - the latter representative of an ecological niche greatly facilitated by the notable reduction in premolar accessory cusps. The many caniforms that also have low ICN scores achieve that status
- not by reducing the accessory cusps independently, but rather by having overall reduced premolar morphology. This reflects their greater reliance on post-carnassial dental elaboration - a condition not found in feliforms and essentially the opposite emphasis compared to that found in hyaenids.
Discussion
Our previous research has shown that recent carnivoran (i.e. Plio-Pleistocene and Pleisto-Holocene) fossil taxa generally sort within the morphospace of the modern congeners and confamilials.1,2,57 However, the pattern seems to be slightly different in older carnivoran clades; for instance, although in one recent analysis two large Mio-Pliocene felids from Langebaanweg fell at the most hypercarnivorous end of the modern felid range, a sample of sympatric hyaenids exhibited significantly higher ICN scores than their modern confamilials.3 As would be expected, detailed analysis of these dental geometries documented the state of the teeth of the ancestors in this lineage before the highly derived modern taxa reached their most durophagous state among modern carnivores. However, questions remained about the dietary patterns of the other enigmatic and diverse members of the LBW carnivore guild - the subject of our current study.
As seen in the previous study of the LBW hyaenids,3 the other LBW carnivorans are very different from their modern analogues. In fact, in some respects, the LBW hyaenids are more like modern hyaenids in their tooth morphology than are the other LWB carnivorans with their modern relatives; that is, in terms of ROC, the LBW hyaenids are indistinguishable from their modern confamilials, and they occupy overlapping morphospace while all of the other LBW carnivorans occupy fairly distinct morphospace (Figure 11). In this respect, even the LBW felids appear quite different from their modern confamilials and the larger Dinofelis-like LBW felids included in the previous analysis3 are more similar to the modern felids than are the other LBW felids that tend to have significantly lower premolar ROC scores and occupy a much wider range of variation in ICN scores than their modern counterparts.
The other LWB carnivorans are harder to compare to a modern sample because they are even more morphologically and phylogenetically distinct than either the LBW felids or hyaenids. Namely, the diverse sample of LBW viverrids, which includes some exceptionally large specimens, occupies morphospace (and likely ecology) that is unoccupied by any modern viverrid. Although the LBW mustelids are not as richly represented in the fossil record, their morphology is also impressive in its extreme divergence from the modern confamilials with giant forms significantly exceeding the range of modern mustelids. Likewise, the two LBW canids seem to represent extreme dietary divergence. Whether Agriotherium is an ursid or hemicyonid it is unlike any modern bear or dog or any other carnivore; like other bears, it has relatively low ICN scores but exceptionally low premolar ROC, which would seem to indicate an extreme adaptation for durophagy, even more so than the modern hyaenids with the exception of its anterior carnassial cusp (the paraconid) ROC score which makes it significantly sharper than not only the modern ursids, but all of the modern carnivoran families including the hypercarnivorous felids. Thus, in some regards its dental morphology appears optimally adapted for extreme durophagy while in other ways the taxon exhibits just the opposite. In the final consideration, this large, impressive carnivoran was likely able to dominate the entire carnivore guild, utilising the whole carcass of its prey possibly more efficiently than any modern carnivoran.
There is still much to be analysed in terms of the dietary niche partitioning of the Mio-Pliocene carnivores from Langebaanweg. For instance, the LBW carnivoran assemblage includes many small specimens, particularly those specimens ascribed to Herpestidae as well as small Viverridae, which are unfortunately beyond the comparative scope of our modern sample. However, the small carnivoran sample from LBW is apparently both taxonomically and morphologically diverse and is worthy of deeper evaluation both in terms of its systematics and ecodiversity. It is our hope that an extension of the ROC and ICN methodology, perhaps with a refinement of its approach to geometric analyses, and with an expansion of the modern comparative sample, will be conducted to evaluate the dietary specialisations of small fossil carnivorans.
Another constraint of this study is that the current analyses are limited in focus by the ICN and ROC metrics. While these have been proven to be highly informative indicators of dietary specialisation, they can only be assessed for specimens that have well-preserved lower premolar and carnassial morphology. But the LBW sample contains elements beyond these teeth that could aid in the dietary reconstruction of its carnivorans. Namely, there are numerous well-preserved maxillary teeth and remarkable crania from most of the represented families. These elements open up possibilities for analyses based on cranial biomechanics and masticatory muscle reconstruction - potentially in three dimensions.
In conclusion, the Mio-Pliocene carnivoran guild from Langebaanweg, South Africa, is clearly impressive both in its systematic and ecological diversity. Although it consists of only a few hundred, mostly fragmentary and isolated specimens, some of the fossils are truly astonishing in terms of their preserved completeness, including postcranial elements beyond the scope of this paper, as well as magnificent whole crania preserved or fully reconstructed for several of the enigmatic species. This unique sample, for its place and time, gives us an unparalleled window into a remarkably complete carnivore guild allowing us to examine subtle niche partitioning between sympatric taxa at a level that exists for few other older palaeontological locations in the world.
Acknowledgements
We thank the staff of Iziko Museums of South Africa and the West Coast Fossil Park, particularly Romala Govender, Graham Avery and Pippa Haarhoff for granting us access to the collections, providing us with valuable feedback on some of our ideas and being wonderful and welcoming hosts. We also thank Deano Stynder for providing valuable discussion and conversation about this research, Francis Thackeray for providing editorial advice, an anonymous reviewer for providing valuable recommendations, and the University of South Carolina for funding this research, particularly the students involved.
Authors' contributions
A.H-R. is the principal investigator and mentor to the other authors (who are his students); he conceptualised the project, taught the students the methods and oversaw all aspects of the data collection; he conducted the analyses and wrote the results and discussion sections and rewrote all other parts of the paper; and he created all of the figures. K.N.B. and K.D.D. performed the majority of data collection. K.N.B. wrote most of the first draft of the introduction and methods. C.L.L. wrote the majority of the introduction and methods as they stand in the current version and undertook the most substantial revision to the manuscript other than the contribution of A.H-R. K.D.D. also contributed to the background as well as editing of the manuscript.
References
1. Hartstone-Rose A, Wahl S. Using radii-of-curvature for the reconstruction of extinct South African carnivoran masticatory behavior. C R Palevol. 2008;7(8):629-643. http://dx.doi.org/10.10167j.crpv.2008.09.015 [ Links ]
2. Hartstone-Rose A. Reconstructing the diets of extinct South African carnivorans from premolar 'intercuspid notch' morphology. J Zool (Lond). 2011;285:119-27. http://dx.doi.org/10.1111/j.1469-7998.2011.00821.x [ Links ]
3. Hartstone-Rose A, Stynder D. Hypercarnivory, durophagy or generalised carnivory in the Mio-Pliocene hyaenids of South Africa? S Afr J Sci. 2013;109(5/6):77-87. http://dx.doi.org/10.1590/sajs.2013/20120040 [ Links ]
4. Van Valkenburgh B. Iterative evolution of hypercarnivory in canids (Mammalia: Carnivora): Evolutionary interactions among sympatric predators. Paleobiology. 1991;17:340-362. [ Links ]
5. Hendey QB. Langebaanweg: A record of past life. Cape Town: South African Museum; 1982. [ Links ]
6. Roberts DL, Matthews T, Herries AIR, Boulter C, Scott L, Dondo C, et al. Regional and Global Palaeoenvironmental and Sea Level Context of the Late Cenozoic Langebaanweg (LBW) Palaeontological Site: West Coast of South Africa. Earth-Sci Rev. 2011;106:191-214. http://dx.doi.org/10.1016/j.earscirev.2011.02.002 [ Links ]
7. Hendey QB. Palaeoecology of the late tertiary fossil occurrences in Quarry, Langebaanweg, South Africa and a reinterpretation of their geological context. Ann S Afr Mus. 1981;84:1-104. [ Links ]
8. Hendey QB. Quaternary vertebrate fossil sites in the south-western Cape Province. S Afr Archaeol Bull. 1969:96-105. http://dx.doi.org/10.2307/3888285 [ Links ]
9. Hendey QB. A review of the geology and palaeontology of the Plio/Pleistocene deposits at Langebaanweg, Cape Province. Ann S Afr Mus. 1970;56:75-117. [ Links ]
10. Hendey QB. Further observations on the age of the mammalian fauna from Langebaanweg, Cape Province. Paleoecology. 1972;6:172-175. [ Links ]
11. Hendey QB. A Pliocene ursid from South Africa. Ann S Afr Mus. 1972;59(6):115-132. [ Links ]
12. Hendey QB, Repenning CA. A Pliocene phocid from South Africa. Ann S Afr Mus. 1972;59:71-98. [ Links ]
13. Hendey QB. Fossil occurrences at Langebaanweg, Cape Province. Nature. 1973;244:13-14. http://dx.doi.org/10.1038/244013a0 [ Links ]
14. Hendey QB. New fossil carnivores from the Swartkrans australopithecine site (Mammalia, Carnivora). Ann Transv Mus. 1974;29(3):27-48. [ Links ]
15. Hendey QB. The late Cenozoic Carnivora of the south-western Cape Province. Ann S Afr Mus. 1974;63:1-369. [ Links ]
16. Hendey QB. Faunal dating of the late Cenozoic of southern Africa, with special reference to the Carnivora. Quaternary Res. 1974;4(2):149-161. http://dx.doi.org/10.1016/0033-5894(74)90004-0 [ Links ]
17. Hendey QB. The Pliocene fossil occurrences in' E' Quarry, Langebaanweg, South Africa. Cape Town: South African Museum; 1976. [ Links ]
18. Hendey QB. Late Tertiary Hyaenidae from Langebaanweg, South Africa, and their relevance to the phylogeny of the family. Ann S Afr Mus. 1978;76:265-297. [ Links ]
19. Hendey QB. Late Tertiary Mustelidae (Mammalia, Carnivora) from Langebaanweg, South Africa. Ann S Afr Mus. 1978;76:329-357. [ Links ]
20. Hendey QB. Agriotherium (Mammalia, Ursidae) from Langebaanweg, South Africa, and relationships of the genus. Ann S Afr Mus. 1980;81:1-109. [ Links ]
21. Hendey QB. Southern African Late Tertiary vertebrates. In: Klein RG, editor. Southern African prehistory and palaeoenvironments. Rotterdam: Balkema; 1984. p. 81-106. [ Links ]
22. Hunt R Jr. Basicranial anatomy of the giant viverrid from Quarry, Langebaanweg, South Africa. In: Stewart KM, Seymour KL, Churcher CS, editors. Palaeoecology and palaeoenvironments of Late Cenozoic mammals: Tributes to the career of CS Churcher. Toronto: University of Toronto Press; 1996. p. 588-597. [ Links ]
23. Kurtén B. The Neogene wolverine Plesiogulo and the origin of Gulo (Carnivora, Mammalia). Helsinki: Societas pro Fauna et Flora Fennica; 1970. [ Links ]
24. Peigné S, De Bonis L, Likius A, Mackaye HT, Vignaud P Brunet M. The earliest modern mongoose (Carnivora, Herpestidae) from Africa (late Miocene of Chad). Naturwissenschaften. 2005;92(6):287-292. http://dx.doi.org/10.1007/s00114-005-0626-0 [ Links ]
25. Peigné S, De Bonis L, Mackaye HT, Likius A, Vignaud P Brunet M. Late Miocene Carnivora from Chad: Herpestidae, Viverridae and small-sized Felidae. C R Palevol. 2008;7(8):499-527. http://dx.doi.org/10.1016/j.crpv.2008.10.002 [ Links ]
26. Howell FC, Garcia N. Carnivora (Mammalia) from Lemudong'o (Late Miocene: Narok District, Kenya). Kirtlandia. 2007;56:121-139. [ Links ]
27. Werdelin L, Sanders WJ. Carnivora. In: Werdelin L, editor. Cenozoic mammals of Africa. Berkeley, CA: University of California Press; 2010. p. 609-663. http://dx.doi.org/10.1525/california/9780520257214.003.0032 [ Links ]
28. Samuels JX, Meachen-Samuels JA, Gensler PA. The first mid-Blancan occurrence of Agriotherium (Ursidae) in North America: A record from Hagerman fossil beds National Monument, Idaho. J Paleontol. 2009;83(4):597-603. http://dx.doi.org/10.1666/08-112R.1 [ Links ]
29. Sorkin B. Ecomorphology of the giant short-faced bears Agriotherium and Arctodus. Hist Biol. 2006;18(1):1-20. http://dx.doi.org/10.1080/08912960500476366 [ Links ]
30. Oldfield C, McHenry C, Clausen P Chamoli U, Parr W, Stynder D, et al. Finite element analysis of ursid cranial mechanics and the prediction of feeding behaviour in the extinct giant Agriotherium africanum. J Zool (Lond). 2012;286:93-171. http://dx.doi.org/10.1111/j.1469-7998.2011.00862.x [ Links ]
31. Stynder DD, Kupczik K. Tooth root morphology in the early Pliocene African bear Agriotherium africanum (Mammalia, Carnivora, Ursidae) and its implications for feeding ecology. J Mammal Evol. 2013;20(3):227-237. http://dx.doi.org/10.1007/s10914-012-9218-x [ Links ]
32. Miller WE, Carranza-Castaóeda O. Agriotherium schneideri from the Hemphillian of Central Mexico. J Mammal. 1996;77(2):568-577. http://dx.doi.org/10.2307/1382830 [ Links ]
33. Pickford M, Soria D. Carnivores from the late Miocene and basal Pliocene of the Tugen Hills, Kenya. Revista de la Sociedad Geológica de Espana. 2005;18(1):39-61. [ Links ]
34. Montoya P, Morales J, Abella J. Eucyon debonisi n. sp., a new Canidae (Mammalia, Carnivora) from the latest Miocene of Venta del Moro (Valencia, Spain). Geodiversitas. 2009;31(4):709-722. http://dx.doi.org/10.5252/g2009n4a709 [ Links ]
35. Lacruz R, Turner A, Berger LR. New Dinofelis (Carnivora: Machairodontinae) remains from Sterkfontein Valley sites and a taxonomic revision of the genus in southern Africa. Ann Transv Mus. 2006;43:89-106. [ Links ]
36. Turner A. Megantereon cultridens (Cuvier) (Mammalia, Felidae, Machairodontinae) from Plio-Pleistocene deposits in Africa and Eurasia, with comments on dispersal and the possibility of a New World origin. J Paleon. 1987;61:1256-1268. [ Links ]
37. Turner A, Antón M. The big cats and their fossil relatives. New York: Columbia University Press; 1997. [ Links ]
38. Werdelin L, Lewis ME. A revision of the genus Dinofelis (Mammalia, Felidae). Zool J Linn Soc. 2001;132(2):147-258. http://dx.doi.org/10.1111/j.1096-3642.2001.tb02465.x [ Links ]
39. Hemmer H. Zur nomenklatur und verbreitung des genus Dinofelis Zdansky, 1924 (Therailurus Priviteau, 1948) [For nomenclature and spread of the genus Dinofelis Zdansky, 1924 (Therailurus Priviteau, 1948)]. Palaeontol Afr. 1965;9:75-89. German. [ Links ]
40. Hemmer H. Neue Befunde zur Verbreitung und Evolution der pliozan-pleistozanen Gattung Dinofelis (Mammalia, Carnivora, Felidae) [New findings on the distribution and evolution of the Pliocene-Pleistocene genus Dinofelis (Mammalia, Carnivora, Felidae)]. Neues Jb Geol Palaeont Mh. 1973;1973(3):157-169. German. [ Links ]
41. Werdelin L, Solounias N. The Hyaenidae: Taxonomy, systematics, and evolution. Fossils Strata. 1991;30:1-104. [ Links ]
42. Stynder DD, Ungar PS, Scott JR, Schubert BW. A dental microwear texture analysis of the Langebaanweg (South African Mio-Pliocene) fossil Hyaenidae. Acta Palaeontol Pol. 2012;57(3):485-496. http://dx.doi.org/10.4202/app.2011.0053 [ Links ]
43. Cruz-Uribe K. Distinguishing hyena from hominid bone accumulations. J Field Archaeol. 1991;18(4):467-486. http://dx.doi.org/10.2307/530408 [ Links ]
44. Werdelin L, Turner A, Solounias N. Studies of fossil hyaenids: The genera Hyaenictis Gaudry and Chasmaporthetes Hay, with a reconsideration of the Hyaenidae of Langebaanweg, South Africa. Zool J Linn Soc. 1994;111(3):197-217. http://dx.doi.org/10.1006/zjls.1994.1022 [ Links ]
45. Tseng ZJ, Stynder D. Mosaic functionality in a transitional ecomorphology: Skull biomechanics in stem Hyaeninae compared to modern South African carnivorans. Biol J Linn Soc. 2011;102:540-559. http://dx.doi.org/10.1111/ j.1095-8312.2010.01602.x [ Links ]
46. Stynder DD. Tooth crown form as an indicator of niche partitioning among Late Miocene/Early Pliocene hyenas from 'E' Quarry, Langebaanweg, South Africa. Paleogeogr Paleoclimatol Paleoecol. 2009;283(3-l):148-159. http://dx.doi.org/10.1016/j.palaeo.2009.09.017 [ Links ]
47. Kretzoi M. Die Raubtiere von Gombaszög nebst einer Übersicht der Gesamtfauna [The predators of Gombaszög along with an overview of the total fauna]. Hist Nat Mus NationHungarici, Pars Miner Geol Paleont. 1938;31:88-157. German. [ Links ]
48. Morales J, Pickford M. Giant bunodont Lutrinae from the Mio-Pliocene of Kenya and Uganda. Estudios Geologicos (Madrid). 2005;61(3-6):233-246. [ Links ]
49. Pasitschniak-Arts M, Larivière S. Gulo gulo. Mamm Species. 1995;499:1-10. [ Links ]
50. Werdelin L. The position of Langebaanweg in the evolution of Carnivora in Africa. Afr Nat History. 2006;2:201-202. [ Links ]
51. Kay RF. Molar structure and diet in extant Cercopithecidae. In: Butler PM, Joysey KA, editors. Development, function, and evolution of teeth. New York: Academic Press; 1978. p. 309-340. [ Links ]
52. Kay RF, Simons EL. The ecology of Oligocene African Anthropoidea. Int J Primatol. 1980;1:21-37. http://dx.doi.org/10.1007/BF02692256 [ Links ]
53. Kirk EC, Simons EL. Diets of fossil primates from the Fayum Depression of Egypt: A quantitative analysis of molar shearing. J Hum Evol. 2001;40(3):203-229. http://dx.doi.org/10.1006/jhev.2000.0450 [ Links ]
54. Van Valkenburgh B. Trophic diversity in past and present guilds of large predatory mammals. Paleobiology. 1988;14:155-173. [ Links ]
55. Walton LR, Joly DO. Canis mesomelas. Mamm Species. 2003;715:1-9. http://dx.doi.org/10.1644/715 [ Links ]
56. Hartstone-Rose A. Evaluating the hominin scavenging niche through analysis of the carcass-processing abilities of the carnivore guild [MSc dissertation]. Durham, NC: Duke University; 2008. [ Links ]
57. Simpler EM, Perry JMG, Hartstone-Rose A. Determining diet from the crossectional shape and intercuspid notches of the teeth of the carnivores of Rancho La Brea. J Vertebr Paleontol. 2011;31(Suppl. 3):195A. [ Links ]
 Correspondence:
Correspondence:
Adam Hartstone-Rose
Department of Cell Biology & Anatomy, University of South Carolina School of Medicine
University of South Carolina
6439 Garners Ferry Road Building 1, CBA rm C-36
Columbia, South Carolina 29209-1445, USA
AdamHR@sc.edu
Received: 15 Dec. 2015
Revised:01 Mar. 2016
Accepted: 05 Mar. 2016


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}