










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.112 n.5-6 Pretoria May./Jun. 2016
http://dx.doi.org/10.17159/sajs.2016/20150278
RESEARCH ARTICLE
A hominin first rib discovered at the Sterkfontein Caves, South Africa
Gaokgatlhe TawaneI; Daniel García-MartínezI, II, III; Jennifer EyreIV, V; Markus BastirI, II; Lee BergerI; Peter SchmidI, VII; Shahed NallaI, VI; Scott A. WilliamsI, IV, V
IEvolutionary Studies Institute and Centre for Excellence in Palaeosciences, University of the Witwatersrand, Johannesburg, South Africa
IIPalaeoanthropology Group, National Museum of Natural Sciences (MNCN-CSIC), Madrid, Spain
IIIBiology Department, Faculty of Sciences, Autonomous University of Madrid, Madrid, Spain
IVCenter for the Study of Human Origins, Department of Anthropology, New York University, New York, New York, USA
VNew York Consortium in Evolutionary Primatology, New York, New York, USA
VIDepartment of Human Anatomy and Physiology, Faculty of Health Sciences, University of Johannesburg, Johannesburg, South Africa
VIIAnthropological Institute and Museum, University of Zurich, Zurich, Switzerland
ABSTRACT
First ribs - the first or most superior ribs in the thorax - are rare in the hominin fossil record, and when found, have the potential to provide information regarding the upper thorax shape of extinct hominins. Here, we describe a partial first rib from Member 4 of the Sterkfontein Caves, South Africa. The rib shaft is broken away, so only the head and neck are preserved. The rib is small, falling closest to small-bodied Australopithecus first ribs (AL 288-1 and MH1). Given that it was recovered near the StW 318 femur excavation, which also represents a small individual, we suggest that the two may be associated. Three-dimensional geometric morphometric analyses were used to quantify the rib fragment morphology and compare it to extant hominoid and other fossil hominin ribs. While only the proximal end is preserved, our analyses show that South African Australopithecus share derived features of the proximal first rib more closely resembling A. afarensis and later hominins than great apes.
Keywords: upper thorax; rib cage; Australopithecus africanus; geometric morphometrics; Gauteng Province
Introduction
Ribs are rare in the human fossil record because of their delicate structure; this scarcity is particularly pronounced in the early hominin fossil record. Moreover, those that have been discovered are usually fragmentary and distorted, and their interpretation is difficult because of the complicated morphological quantification of their 3D curvature.1-5 In spite of the scarcity of these remains, it is important to note that, among ribs, the first rib is generally the best preserved in the fossil record as a result of its unique morphology relative to the rest of the ribs in the thorax.6,7 The first rib is also important because it bears diagnostic features informative of upper thorax morphology.7-9 For example, the modern human first rib usually has a univertebral articulation (with the T1 vertebra), as do those of early hominins, whereas the other great apes have a bivertebral articulation (with the C7 and T1 vertebrae).10
In early hominins, first ribs are known from Australopithecus afarensis (AL 288-1ax)11, Homo erectus (KNM-WT 15000 AG and AY&AZ)12, A. sediba (MH1, UW88-148; MH2, UW88-198 and UW88-187)8 and H. naledi (UW101-83)13. A. afarensis AL 288-1 and A. sediba (MH1 and MH2) were hypothesised as presenting a narrow upper thorax, as extant apes do, and this thoracic shape is probably related to suspensory locomotor behaviour.8,11,14,15 This narrow upper thorax has also been observed in the recently published small-bodied species H. naledi.13 In contrast, other early hominin specimens, such as H. erectus (KNM-WT 15000) and the large-bodied A. afarensis (KSD-VP-1/1), have been proposed to have modern human-like, expanded upper thoraces.12,16
Sterkfontein Caves (located 40 km from Johannesburg) - one of the most important South African sites regarding the quantity and quality of the fossils discovered17-20 - has yielded several costal remains, but a first rib has not been reported from this site to date. The A. africanus partial skeleton Sts 14, discovered at Sterkfontein in 1947, is associated with a number of ribs.21 Robinson's analysis positioned the preserved ribs toward the lower thorax. According to Robinson21, these ribs are smaller than those of modern H. sapiens but are characterised by a similar degree of curvature. The features observed on the ribs suggested a modern human-like lower thorax shape, but there is still some degree of uncertainty about the upper thorax shape of A. africanus.
In 1987, another partial skeleton, StW 431, this time of a larger-bodied, presumed male A. africanus was discovered at Sterkfontein in Bed B of Member 4.22 StW 431 preserves a right rib with the head, neck and tubercle, which was assessed as a probable third rib.22 However, the preservation of this fragment was not sufficient to discuss upper thorax morphology in A. africanus from an evolutionary point of view.
Here we report, present and discuss, in a comparative anatomical context, a well-preserved proximal part of a first rib previously recovered at Sterkfontein but neither identified nor published, as well as its provenance and its relation to the previously discovered remains from the Sterkfontein Caves.
Materials and methods
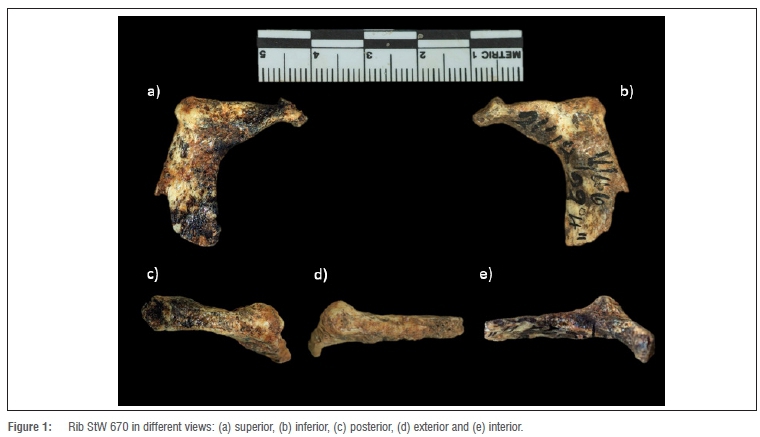
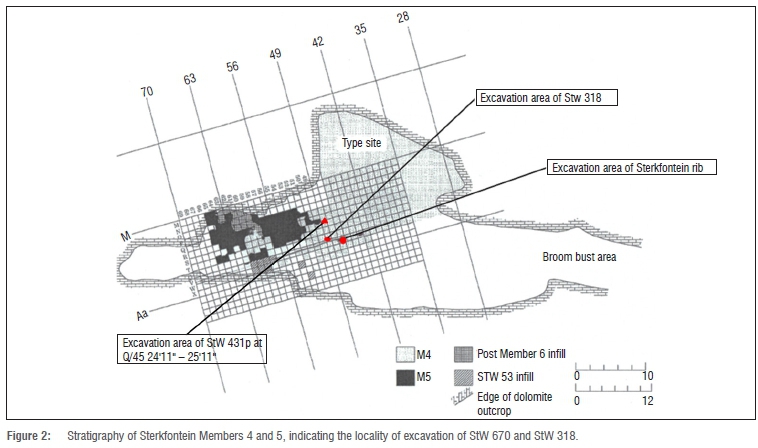
The new fossil, a proximal fragment of a first rib from the right side (StW 670; Figure 1) was discovered at Sterkfontein site (Cradle of Humankind, Gauteng, South Africa) in grid square V 46, at 21 feet 1 inches to 22 feet 4 inches (27'2-28'2), Member 423 (Figure 2). The date of the discovery of the rib is unknown. The collection from Sterkfontein Caves housed at the University of the Witwatersrand dates from 1966 to present excavations. Material collected from Sterkfontein prior to 1966 is housed at Ditsong Museum (formerly known as the Transvaal Museum).
Preservation, morphology and ontogenetic assessment of StW 670
A detailed description of the preservation status as well as the morphology was carried out on StW 670 based on the principal anatomical features of the first rib. Frequencies of single/double articular facet/s of rib head in the comparative sample were also studied (Supplementary table 1 of the supplementary material).
The maturation state of the first rib was evaluated based on the epiphyseal fusion of the articular tubercle of the rib and the articular facet of the rib head.24 However, it should be noted that the maturation rate of Australopithecus epiphyseal fusion could differ from that of H. sapiens.
Linear measurements
Linear measurements taken on StW 670 were carried out on the original remains. The linear measurements taken were: head height (superoinferior diameter of the rib head), head width (anteroposterior diameter of the rib head), neck length (distance from the medial-most point on the rib head to the medial-most point on the articular facet of the tubercle) and neck width (taken midway between the rib head and articular tubercle in anteroposterior dimension).2,9,25 Each measurement was calculated from the average of three measurements in order to reduce intra-observer error.1,2,25 The measurements are given in Table 1.
Measurements on StW 670 were made with standard anthropometric instruments following the definitions used in previous studies.2,9,25
Geometric morphometric analyses
For 3D geometric morphometrics, 3D high-resolution laser scans of original fossils - StW 670, A. sediba MH1 (UW88-148) and MH2 (UW88-198) - and of a high-quality cast of AL 288-1ax (A. afarensis) were used. These scans were obtained using a NextEngine 3D laser scanner in 'wide mode' (with a resolution of 0.38 mm and an accuracy of 6 points per mm). Additionally, computerised tomography (CT) scans of the original remains of H. erectus KNM-WT 15000 (AG and AY&AZ) were employed for the analyses (Supplementary table 1).
For the 3D geometric morphometrics analyses, remains of 33 H. sapiens (20 Europeans, 10 sub-Saharan Africans, 1 small-bodied member of the San population and 2 small-bodied individuals from the Andaman Islands) were analysed (Supplementary table 1). These scans were also obtained through a NextEngine 3D laser scanner in 'wide mode' (with a resolution of 0.38 mm and an accuracy of 6 points per mm). Additionally, 8 ribs of Pan troglodytes (chimpanzee), 3 ribs of Gorilla gorilla (gorilla), 3 ribs of Pongo sp. (orangutan) and 3 ribs of Hylobates sp. (gibbon) from Kyoto University Primate Research Institute (www.pri.kyoto-u.ac.jp) were used to represent non-hominin morphological pattern (Supplementary table 1). Technical specifications of CT scanning can be obtained from www.pri.kyoto-u.ac.jp.
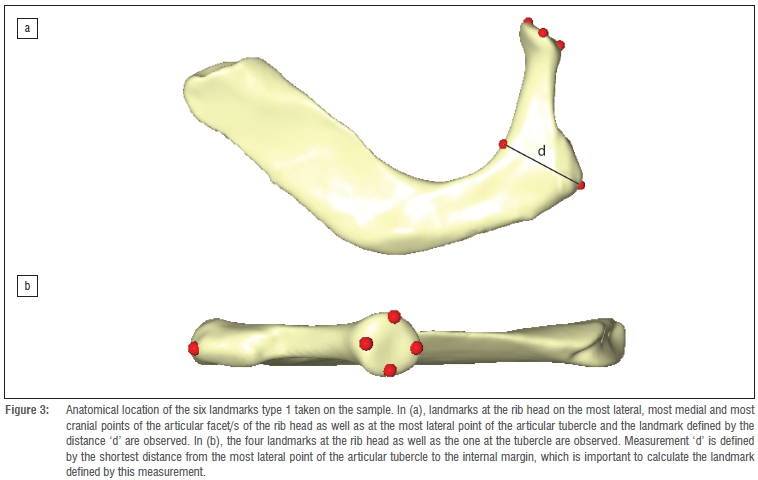
Six 3D type 1 landmarks were collected on each rib, quantifying morphological information of the rib head and the articular tubercle as well as the thickness of the rib neck (Figure 3). Landmarks were collected at the rib head on the most lateral, most medial, most caudal and most cranial points of the articular facet/s of the rib head and at the most lateral point of the articular tubercle. Additionally, one landmark was located at the point defined by the shortest distance from the most lateral point of the articular tubercle to the internal margin of the rib curvature ('d' distance in Figure 3). Because StW 670 is missing a large part of the body of the rib and the sternal end, no landmarks were taken on this area of the comparative sample.
Size was measured as centroid size.26,27 The size relations between the Sterkfontein first rib and the rest of the individuals in our sample were explored using a box plot. These analyses were carried out in PAST software version 3.28 In order to study the shape relations along the variability of the sample, we superimposed the landmark configurations using generalised Procrustes analysis.26,27 The superimposed coordinates were then submitted to principal component analysis (PCA) in shape space in order to reduce the dimensionality of data and to visualise the main axes of variation.
Ordinations were computed into MorphoJ 1.05f software29 and the shape differences of the surface associated with PC1 and PC2 axes were warped and visualised using the EVAN Toolkit software version 1.62 (www.evan-society.org/). A box plot of Procrustes distances was used to compare and explore shape relations from the individual under study (StW 670) to the different groups of the sample. These analyses were carried out in the Virtual Morphology Lab of the National Museum of Natural Sciences (Madrid, Spain).
Results
Preservation, morphology and ontogenetic assessment of StW 670
There is sufficient preservation of features in the first rib fragment (StW 670) to make description, comparison and analysis possible, even though the fossil is incomplete. The fragment has a well-preserved head, neck, tubercle and most of the proximal part of the body, although the rest of the body is lacking distally.
The rib fragment StW 670 was determined as a first rib from the right side. Siding was determined because the head is positioned at an angle caudal to the tubercle and shaft which produces a mediolateral slope of the neck. Moreover, as the caudal surface of the shaft is smoother than the cranial surface, this feature also confirms the side of the rib as a first rib from the right side.
The head has a rim that is thickened for the attachment of the costovertebral ligament. The angle and the tubercle coincide - a feature that is presented in extant and fossilised primate first ribs. The neck is narrow proximally and widens distally as the tubercle is approached. The neck is also rounded proximally and flattened distally. The tubercle is well developed and has a well-defined, smooth and rounded articular facet. The non-articular part of the tubercle for the attachment of the lateral costotransverse ligament is present.
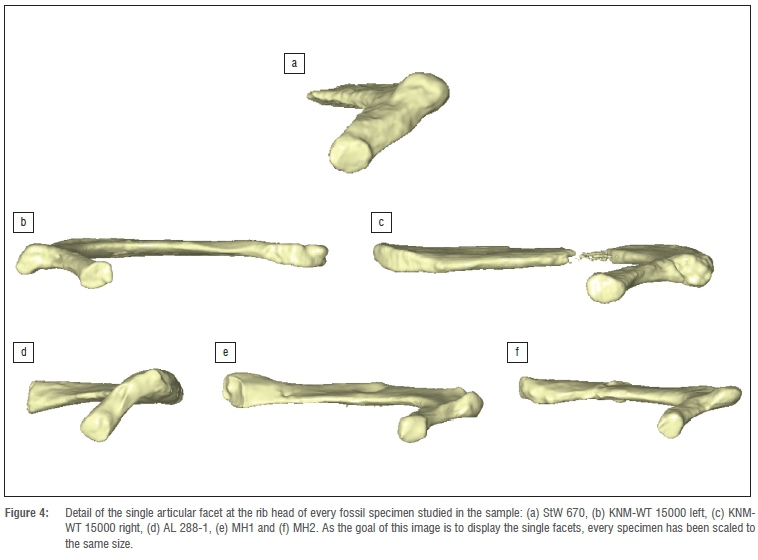
The head has a single articular facet, which is rounded - a morphology that most closely resembles modern humans. The rest of the fossil specimens studied also presented a single articular facet (Figure 4). Regarding the comparative sample, 95% of the European modern humans presented a single articular facet at the rib head for articulation with T1, while 100% of the African modern humans presented this feature. In contrast, all of the non-hominin hominoids (Pan, Gorilla, Pongo and Hylobates) presented a double facet for articulation with C7 and T1.
The preservation of the fragment does not allow for the finer detail of anterior scalene and anterior serrate muscle insertions elevations, but it does allow us to study the scalene medium muscle insertion, which is only slightly marked in StW 670. The epiphysis at the articular tubercle and the rib head are well preserved and totally fused with the metaphysis, suggesting that the individual was adult at the time of death.24
Geometric morphometric analyses
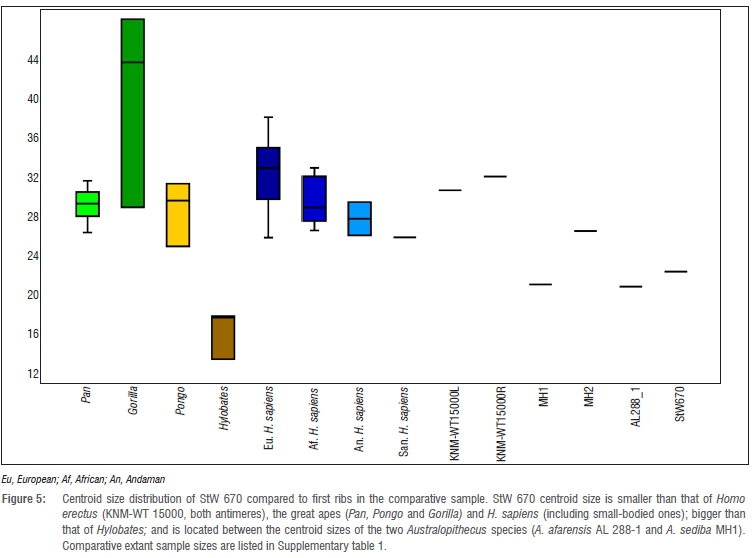
Figure 5 shows a box plot of centroid size distribution of the different groups included in the sample. The centroid size of the StW 670 fossil is smaller than that of H. erectus (KNM-WT 15000, both antimeres), A. sediba (MH2), the great apes (Pan, Gorilla and Pongo) and H. sapiens (including the small-bodied individuals), and larger than that of Hylobates, and is located closest to the centroid size of A. afarensis AL 288-1 and A. sediba MH1.
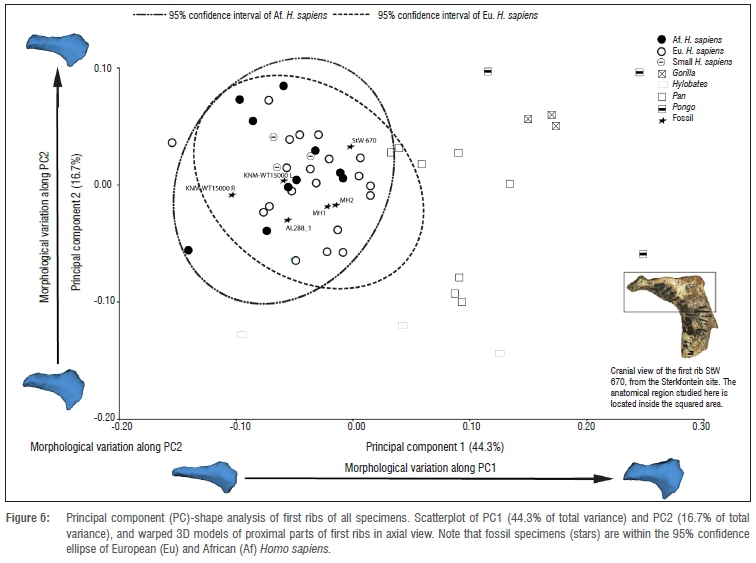
Regarding shape, in Figure 6, we can observe that PC1 (44.3% of the total variation) polarises most of the non-hominin hominoids on the positive side of the axis, whereas the scatter of H. sapiens of different populations is located mostly towards the negative values of the PC1 axis. As it can be observed on the associated warps (Figure 6), the positive values of PC1 (associated with great apes) correspond to first ribs with a shorter relative distance between the head and the articular tubercle and a larger neck width at the articular tubercle. The negative values of PC1 (mainly associated with hominins) correspond to first ribs with a larger relative distance between the head and the articular tubercle and a shorter neck width at the articular tubercle. Moreover, PC1 polarisation differentiates between different rib head orientations. That is, in the positive values of PC1 (associated with non-hominin hominoids), the plane of the rib head is more oblique to the plane of the neck, while in the negative values of PC1 (mainly associated with modern humans and fossil specimens - so hominins), the plane of the rib head is more parallel to the plane of the neck.
As PC2 (16.7% of the total variation), which explains intraspecific variation, does not polarise between groups observed in PC1, as is observed in Figure 6, we will not discuss PC2. The rest of the principal components explain little of the variation so they also will not be discussed here.
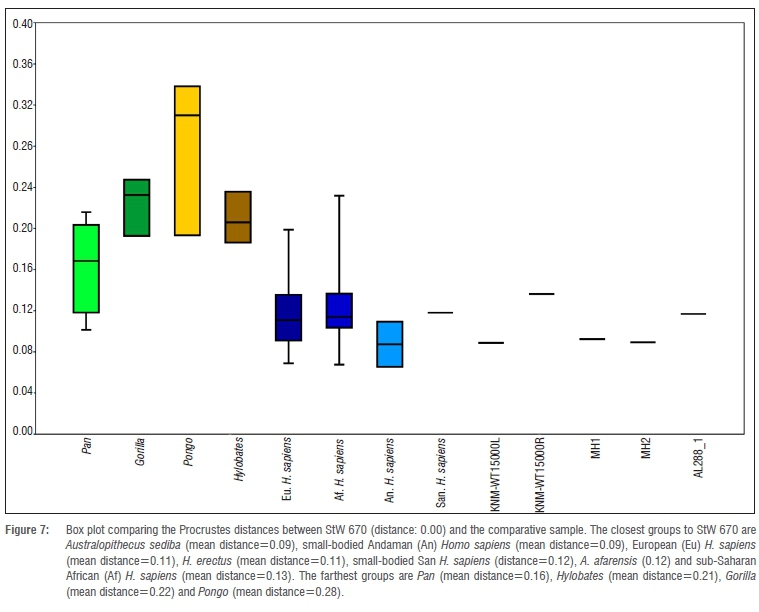
Finally, looking at shape similarities according to the Procrustes distances, it is observed in the box plot (Figure 7) that the closest groups to StW 670 are A. sediba (mean distance=0.09), small-bodied Andaman H. sapiens (mean distance=0.09), European H. sapiens (mean distance=0.11), H. erectus (mean distance=0.11), small-bodied San H. sapiens (distance=0.12), A. afarensis (distance=0.12) and the sub-Saharan African H. sapiens (mean distance=0.13). The farthest groups are P. troglodytes (mean distance=0.16), Hylobates sp. (mean distance=0.21), G. gorilla (mean distance=0.22) and Pongo sp. (mean distance=0.28).
Discussion and conclusion
Sterkfontein Member 4 has yielded quite a number of hominin remains, with Sts 14 and StW 431 (both A. africanus) among the best-known and well-studied hominins from this locality.21,22 Four beds are recognised within Member 4 - labelled A, B, C and D.23 StW 431 was excavated in Member 4 Bed B. The excavation beds of Sts 14 and StW 670 were apparently never recorded, so we do not know for certain from which bed StW 670 was excavated. However, StW 670 was excavated close to the excavation area of StW 318, a femur found at U/49 24'0"-25'0". Berger and McHenry30 surmised the femur to be from an individual of small stature, weighing about 45-50 kg. The small size of the StW 670 rib may mean it is associated with this femur.
Three-dimensional geometrics morphometric analyses, which take into account the 3D spatial relationship between rib head and tubercle, show that the morphology of the preserved portion of StW 670 first rib is quite similar to A. sediba as it is close to this taxon in the box plot of the Procrustes distances. Regarding the morphological similarities with the rest of the groups, we can see that, besides A. sediba, the morphology of StW 670 is more similar to the H. sapiens groups, H. erectus and A. afarensis than it is to non-hominin hominoids (Pan, Pongo, Gorilla and Hylobates). In fact, in the PCA analyses, every fossil specimen (StW 670, MH1, MH2, AL 288-1 and KNM-WT 15000) falls inside the 95% confidence interval ellipse of H. sapiens (Figure 6).
Morphologically, the similarity of the fossils to modern humans observed in this PCA is because these groups are characterised by a larger relative distance between the head and the articular tubercle, and a shorter neck width at the articular tubercle, and because the plane of the rib head is more parallel to the plane of the neck in modern humans and the fossils than in the non-human hominoids. This latter feature observed at the rib head should probably be reflected in the orientation of the transverse processes of the T1 vertebrae, thus future studies should investigate this possibility.
Additionally, the presence of a single facet observed in every early hominin studied (Australopithecus and H. erectus) also supports the modern-like pattern of this fragment. This observation contrasts strongly with the pattern observed in the great apes (Pan, Pongo, Gorilla and Hylobates) as all of them present two articular facets at the rib head. Although we are conscious of the limitations of our sample size, we think that the observations made here are relevant to discuss the modernity of this character in StW 670. So, in the light of these observations, we conclude that the single facet of the rib head of the first rib is a feature unique to the hominin lineage, which emerged, at least, with the genus Australopithecus.10 Moreover, aspects observed in the rib head should be reflected in the adjacent vertebrae as well, but because an associated vertebra does not exist and an assessment of vertebral morphology is not the goal of this work, it should be addressed in future studies.
The similarity of the proximal rib morphology observed in Australopithecus and H. sapiens first ribs is interesting because it has traditionally been thought that the upper thorax morphology of Australopithecus is characterised by a great ape-like form.6,7 So, another hypothesis to be tested in future studies is that if we accept the derived morphology of the proximal part of the first rib in Australopithecus (A. afarensis, A. sediba and A. africanus), we can infer that the archaic pattern of the upper Australopithecus thorax should be found at the distal part of the ribs. This would suggest that the proximal part of the rib became derived in the direction of the H. sapiens morphology earlier than did the distal part of the first rib. This suggestion is consistent with the fact that the proximal and distal parts of the first rib belong to different morphogenetic modules in ontogeny,31 which allows both units to evolve at different rates according to different requirements.
Great apes are often described as having a narrow upper thorax, while humans have a more expanded one.32 It would be useful to be able to use fragmentary ribs in the hominin fossil record to determine when the transition between the two morphologies took place. The 2D analyses performed in previous studies do not make this goal look promising because of the complex morphology of ribs and the ribcage more broadly; however, the 3D analyses performed here show that the spatial configuration of the rib head and articular tubercle could be informative about the archaic or derived morphology, at least of the proximal part of the first rib. This fact, together with features such as the single facet of the rib head10, could be informative in finding evolutionary traits in the upper thorax of early hominins. More research employing 3D techniques is needed on complete first ribs as well as the rest of the ribs in the costal sequence (from first to twelfth) in order to better quantify and understand Australopithecus rib morphology and the role of the thorax in hominin evolution more broadly.
Acknowledgements
We thank Bernhard Zipfel and the Access Committee of the Evolutionary Studies Institute, University of the Witwatersrand, as well as the South African National Centre of Excellence in Palaeosciences for hosting us whilst the materials were studied. D.G-M. and M.B. were funded through CGL2012-37279-MINECO (Spain) and a Leakey Research Grant to D.G-M. S.A.W. and J.E. were funded through the New York University Research Challenge Fund. We thank Emma Mbua, Fred Spoor and the National Museums of Kenya Earth Science Department for providing CT scans of the KNM WT-15000 axial skeleton. We also thank the reviewers for their comments which improved the previous version of the manuscript.
Authors' contributions
G.T. identified the rib fragment as hominin and described it. D.G-M. and J.E. analysed the 3D and linear data, respectively. G.T., D.G-M., J.E., S.N. and S.A.W. wrote the paper. M.B., P.S. and L.B. provided data and supervisory support, respectively.
References
1. Franciscus RG, Churchill SE. The costal skeleton of Shanidar 3 and a reappraisal of Neandertal thoracic morphology. J Hum Evol. 2002;42:303-356. http://dx.doi.org/10.1006/jhev.2001.0528 [ Links ]
2. Gómez-Olivencia A, Eaves-Johnson KL, Franciscus RG, Carretero JM, Arsuaga JL. Kebara 2: New insights regarding the most complete Neandertal thorax. J Hum Evol. 2009;57:75-90. http://dx.doi.org/10.1016/j.jhevol.2009.02.009 [ Links ]
3. Bastir M, García-Martínez D, Recheis W, Barash A, Coquerelle M, Rios L, et al. 3D analysis of human ribcage ontogeny. Am J Phys Anthropol. 2013;S56:75. [ Links ]
4. Garcia-Martinez D, Barash A, Recheis W, Utrilla C, Sanches T, Garcia Rio F, et al. On the chest size of Kebara 2. J Hum Evol. 2014;70:69-72. http://dx.doi.org/10.1016/j.jhevol.2014.02.003 [ Links ]
5. García-Martínez D, Recheis W, Bastir M. Ontogeny of 3D rib curvature and its importance for the understanding of human thorax development. Am J Phys Anthropol. 2016;159(3):423-431. http://dx.doi.org/10.1002/ajpa.22893 [ Links ]
6. Aiello L, Dean C. An Introduction to human evolutionary anatomy. London: Academic Press; 1990. [ Links ]
7. Nalla S. The morphology of the upper thorax of Australopithecus sediba within the context of the selected hominoids [PhD thesis]. Johannesburg: University of the Witwatersrand; 2013. [ Links ]
8. Schmid P Churchill SE, Nalla S, Weissen E, Carlson KJ, De Ruiter DJ, et al. Mosaic morphology in the thorax of Australopithecus sediba. Science. 2013;340:163-165. http://dx.doi.org/10.1126/science.1234598 [ Links ]
9. Bastir M, Garcia-Martinez D, Estalrrich A, García-Tabernero A, Huguet R, Ríos L, et al. The relevance of the first ribs of the El Sidron site (Asturias, Spain) for the understanding of the Neandertal thorax. J Hum Evol. 2015;80:64-73. http://dx.doi.org/10.1016/j.jhevol.2014.10.008 [ Links ]
10. Ohman JC. The first rib of hominoids. Am J Phys Anthropol. 1986;70:209-229. http://dx.doi.org/10.1002/ajpa.1330700208 [ Links ]
11. Johanson DC, Lovejoy CO, Kimbel WH, White TM, Ward SC, Bush ME, et al. Morphology of the Pliocene partial hominid skeleton (A.L. 288-1) from the Hadar Formation, Ethiopia. Am J Phys Anthropol. 1982;57:403-451. http://dx.doi.org/10.1002/ajpa.1330570403 [ Links ]
12. Jellema L, Latimer B, Walker A. The rib cage. In: Leakey RE, Walker A, editors. The Nariokotome Homo erectus skeleton. Cambridge, MA: Harvard University Press; 1993. p. 294-325. http://dx.doi.org/10.1007/978-3-662-10382-1_13 [ Links ]
13. Berger LR, Hawks J, De Ruiter DJ, Churchill SE, Schmid P Delezene LK, et al. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife. 2015;4, e09560, 35 pages. http://dx.doi.org/10.7554/eLife.09560 [ Links ]
14. Schmid P. The trunk of the australopithecines. Paris: Origine(s) de la Bipédie chez les Hominidés Presses du CNRS; 1991. p. 225-234. [ Links ]
15. Williams SA. Modern or distinct axial bauplan in early hominins? Comments on Haeusler et al. (2011). J Hum Evol. 2012;63:552-556. http://dx.doi.org/10.1016/j.jhevol.2012.01.007 [ Links ]
16. Haile-Selassie Y Latimer BM, Alene M, Deino AL, Gibert L, Melillo SM, et al. An early Australopithecus afarensis postcranium from Woranso-Mille, Ethiopia. Proc Natl Acad Sci USA. 2010;107(27):12121-12126. http://dx.doi.org/10.1073/pnas.1004527107 [ Links ]
17. Broom RR, Robinson JT. Man contemporaneous with the Swartkrans apeman. Am J Phys Anthropol. 1950;8:151-156. http://dx.doi.org/10.1002/ajpa.1330080211 [ Links ]
18. Hughes AR, Tobias PV. A fossil skull probably of the genus Homo from Sterkfontein, Transvaal. Nature. 1977;265:310-312. http://dx.doi.org/10.1038/265310a0 [ Links ]
19. Kuman K, Clarke RJ. Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5. J Hum Evol. 2000;38:827-847. http://dx.doi.org/10.1006/jhev.1999.0392 [ Links ]
20. Partridge TC, Granger DE, Caffee MW, Clarke RJ. Lower Pliocene hominid remains from Sterkfontein. Science. 2003;300:607-612. http://dx.doi.org/10.1126/science.1081651 [ Links ]
21. Robinson JT. Early hominid posture and locomotion. Chicago: University of Chicago Press; 1972. [ Links ]
22. Toussaint M, Macho GA, Tobias PV, Partridge TC, Hughes AR. The third partial skeleton of a late Pliocene hominin (StW 431) from Sterkfontein, South Africa. S Afr J Sci. 2003;99:215-223. [ Links ]
23. Partridge TC, Watt IB. The stratigraphy of the Sterkfontein hominid deposit and its relationship to the underground cave system. Palaeontol Afr. 1991;28:35-40. [ Links ]
24. Rios L, Cardoso HFV. Age estimation from stages of union of the vertebral epiphyses of the ribs. Am J Phys Anthropol. 2009;140:265-274. http://dx.doi.org/10.1002/ajpa.21065 [ Links ]
25. Gómez-Olivencia A, Carretero JM, Lorenzo C, Arsuaga JL, Bermúdez de Castro J-M, Carbonell E. The costal skeleton of Homo antecessor: preliminary results. J Hum Evol. 2010;59:620-640. http://dx.doi.org/10.1016/j.jhevol.2010.07.023 [ Links ]
26. Zelditch ML, Swiderski DL, Sheets HD, Fink WL. Geometric morphometrics for biologists: A primer. London: Academic Press; 2004. [ Links ]
27. O'Higgins P. The study of morphological variation in the hominid fossil record: Biology, landmarks and geometry. J Anat. 2000;197:103-120. http://dx.doi.org/10.1046/j.1469-7580.2000.19710103.x [ Links ]
28. Hammer 0, Harper D, Ryan P PAST: Paleontological statistics software package for education and data analysis. Palaeontol Electron. 2001;4(1), Art. #4, 9 pages. http://palaeo-electronica.org/2001_1/past/issue1_01.htm [ Links ]
29. Klingenberg CP. MorphoJ: An integrated software package for geometric morphometrics. Mol Ecol Resour. 2011;11:353-357. http://dx.doi.org/10.1111/j.1755-0998.2010.02924.x [ Links ]
30. Berger LR, McHenry HM. Body proportions in Australopithecus afarensis and A. africanus and the origin of the genus Homo. J Hum Evol. 1998;35:1-22. http://dx.doi.org/10.1006/jhev.1997.0197 [ Links ]
31. Aoyama H, Mizutani-Koseki S, Koseki H. Three developmental compartments involved in rib formation. Int J Dev Biol. 2005;49:325-333. http://dx.doi.org/10.1387/ijdb.041932ha [ Links ]
32. Schultz AH. Vertebral column and thorax. Primatologia. 1961;4:1-66. [ Links ]
 Correspondence:
Correspondence:
Daniel García-Martínez
Palaeoanthropology Group, Museo Nacional de Ciencias Naturales (MNCN-CSIC) - Paleobiology
José Gutierrez Abascal 6
28006 Madrid, Spain
Email: dan.garcia@mncn.csic.es
Received: 29 July 2015
Revised: 11 Dec. 2015
Accepted: 08 Jan. 2016
Note: This article includes supplementary material


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}