










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.112 n.3-4 Pretoria Mar./Apr. 2016
http://dx.doi.org/10.17159/sajs.2016/20150086
REVIEW ARTICLE
Tuberculous meningitis in infants and children: Insights from nuclear magnetic resonance metabolomics
Shayne MasonI; Carolus J. ReineckeI; Regan SolomonsII; A. Marceline van FurthIII
ICentre for Human Metabonomics, Faculty of Natural Sciences, North-West University, Potchefstroom Campus, Potchefstroom, South Africa
IIDepartment of Paediatrics and Child Health, Faculty of Medicine and Health Sciences, Stellenbosch University, Stellenbosch, South Africa
IIIDepartment of Paediatric Infectious Diseases and Immunology, VU University Medical Center, Amsterdam, the Netherlands
ABSTRACT
Tuberculous meningitis (TBM) is a prevalent form of central nervous system tuberculosis (CNS-TB) and the most severe common form of bacterial meningitis in infants and children below the age of 13 years, especially in the Western Cape Province of South Africa. Research to identify markers for timely and accurate diagnosis and treatment outcomes remains high on the agenda for TBM, in respect of which the field of metabolomics is as yet largely unexploited. However, the national Department of Science and Technology (DST) recently established several biotechnology platforms at South African institutions, including one for metabolomics hosted at North-West University. We introduce this national platform for nuclear magnetic resonance (NMR) metabolomics by providing an overview of work on TBM. We focus on selected collaborative multidisciplinary approaches to this disease and conclude with the outcomes of an untargeted NMR metabolomics study of cerebrospinal fluid from TBM patients. This study enabled the formulation of a conceptual shuttle representing the unique metabolic plasticity of CNS metabolism towards the energy requirements for the microglia-driven neuroinflammatory responses, of which TBM is one example. From insights generated by this explorative NMR metabolomics investigation, we propose directions for future in-depth research strategies to address this devastating disease. In our view, the timely initiative of the DST, the operational expertise in metabolomics now available and the potential for involving national and international networks in this field of research offers remarkable opportunities for the future of metabolomics in South Africa and for an ever greater understanding of disease mechanisms.
Keywords: TBM; NMR spectroscopy; cerebrospinal fluid; CSF; hypothesis
Introduction
Metabolomics has become an established scientific field. It provides analytical, chemical and physiological insights into naturally occurring, low-molecular-weight organic metabolites within organisms, cells, tissues and biofluids, and helps to promote understanding of metabolite interactions.1 Explorative metabolomics investigations produce metabolite profiles and rely on the scientific method of induction to generate hypotheses in order to identify metabolites that may be indicators or biomarkers of disease or used as monitors of therapeutic responses.2 Tuberculosis (TB), in its variety of manifestations - such as pulmonary and pleural TB, as well as TB within the central nervous system (CNS-TB), with tuberculous meningitis (TBM) as one of its most dangerous forms - is high on the list for such applications. Although still limited in number, metabolomic technologies have been applied successfully to diseases of the CNS. The composition of cerebrospinal fluid (CSF), which is partially derived from interstitial fluid in the CNS, is anticipated to reflect the normal and pathological biochemical processes of the brain. In the present review, we start by addressing the pathophysiology and severity of TBM and follow with a discussion on the application of metabolomics in enriching our understanding of this infectious disease. Research on TB, in its broadest sense, has a long-standing history in South Africa, emanating from several leading research units and centres, and directed and staffed by world-renowned scholars. Their contributions warrant a review alone and are not covered here; our main focus is on TBM, the latest pioneering investigations using nuclear magnetic resonance (NMR) metabolomics, and perspectives on methodological approaches that further exploratory insights and hypotheses formulated through metabolomics studies.
Tuberculosis is caused by the bacterium Mycobacterium tuberculosis. Although TB commonly presents as a pulmonary disease, systemic spread of the tubercule bacilli can lead to extrapulmonary forms of TB (EPTB), present in up to 20% of reported TB cases,3 and particularly common in young children and immunocompromised individuals4. CNS-TB represents up to an estimated 10% of all EPTB (and 1% of total TB) cases.3,5 According to the World Health Organization Global TB Report for 2014, there were 312 380 clinically defined new cases of TB in South Africa alone; of these, 37 709 (12%) were EPTB, which included an estimated 3771 cases of CNS-TB. Among the various manifestations of TB, TBM is not only the most prevalent form of CNS-TB but is also the most severe complication of TB and the most common form of bacterial meningitis in children below the age of 13 years6, especially in the Western Cape Province of South Africa7,8.
Recent reviews have covered various aspects of childhood TBM.9-11 The general consensus emerging from these reports is that: (1) progress in understanding, prevention and treatment of TBM remains inadequate; (2) childhood deaths and disability as a result of TBM constitute a major societal burden; (3) although preliminary results obtained with the Bacillus Calmette-Guérin (BCG) vaccine seem to be promising for protection against TB, future trials of candidate vaccines are still needed; and (4) novel research to improve timely and accurate diagnosis and treatment outcomes remains high on the agenda. TBM is a disease that is largely unexploited by metabolomics technology, which predominantly makes use of hyphenated mass spectroscopy (MS) and NMR spectroscopy - the latter being the focus of this overview.
Pathogenesis and severity of TBM
Tuberculosis is spread via an aerosol route from persons who have active disease. Most people infected with M. tuberculosis have latent disease and are asymptomatic. However, a small proportion of individuals go on to develop active TB - with some developing systemic TB, such as TBM - and have significant morbidity and mortality, even though effective treatment is available.
In the late 19th century, it was thought that TBM resulted from haema-togenous spread to the meninges, as a result of the frequent finding that TBM and miliary TB were occurring in the same patients.12 In 1933 Rich and McCordock13 published their report that, in the majority of postmortems, a single caseous focus (Rich's focus) could be found, from which, when ruptured, bacilli could spread to the subarachnoid space. It was thought that this single lesion was almost always older than all the lesions found in concurrently occurring miliary TB, which set the paradigm for understanding the pathogenesis of TBM. However, this model did not fully explain the frequency of miliary TB and TBM occurring simultaneously14, nor the mechanism whereby M. tuberculosis spreads from the lungs to the meninges and crosses the blood-brain barrier15. Animal models have been developed in order to understand better the pathogenesis of CNS-TB but findings are frustrated by the poor human clinical-pathological correlation.15 Magnetic resonance imaging has detected numerous concurrent leptomeningeal granulomas in children with miliary TB, further challenging Rich's pathogenic model.15,16 It is likely that early haematogenous spread to the brain occurs before a T-cell mediated immune response is activated. This mechanism could explain the vulnerability to TBM when T-cell mediated immunity is sub-optimal in persons infected with the human immunodeficiency virus (HIV)9 and in persons on long-term glucocorticoid therapy17. The protective role of lymphocytes is essential, with the contribution of CD4+ and CD8+ T-cells, along with macrophages in isolating and engulfing M. tuberculosis; together, these eventually lead to granuloma formation. We, and others, have shown that many of the signs, symptoms and sequelae of TBM result from an immunologically directed inflammatory response to the infection.18,19
Greater understanding of the entry of M. tuberculosis into the CNS and the immunological mechanisms allowing survival of the bacilli is crucial for improving prevention and treatment. Fluctuation in cytokine levels affecting immunological function in patients with TBM can directly influence the duration of the disease and its severity.18 In particular, the balance between pro- and anti-inflammatory cytokines may be crucial to TBM disease progression19; this relationship is reflected by the upregulation in CSF of pro- and anti-inflammatory cytokines in patients with TBM compared to patients with other forms of meningitis.20,21 The early clinical presentation of TBM is often non-specific, with symptoms such as cough, loss of weight, fever, vomiting and malaise. As the disease progresses, meningism, focal neurological signs, and a depressed level of consciousness can occur. The timing of initiation of treatment is the most critical factor affecting morbidity, mortality, and healthcare costs, which emphasises the importance of early diagnosis of TBM.22 Through a large retrospective study of 554 children with TBM, we established that 14% presented with visual impairment, 16% with hearing loss, 44% with motor deficits and 77% with intellectual impairment; only a small number (16%) did not have neurological manifestations of TBM.23
Accurate prediction of outcome in childhood TBM is difficult owing to the diversity of underlying pathological mechanisms and variation in host immunological response. Multidrug-resistant (MDR) TBM in children has a poor clinical outcome and is associated with increased mortality.24 In-patient mortality rates do not differ between HIV-infected and non-HIV-infected children with TBM; however, mortality after hospital discharge is greater in HIV-infected children with TBM because of HIV-related illnesses.25 Even though outcome prediction is problematic, the clinical stage of disease at the time of starting treatment is predictive of prognosis.11 Children with Stage I TBM disease are likely to lead a normal life without neurological complications, whereas those with Stage III disease have a high risk of mortality.11 Of all the TBM staging systems, the refined Medical Research Council scale has been shown to have the highest predictive value.26,27 The criteria that define each stage are as follows: Stage I - Glasgow Coma Scale (GCS) of 15 and no focal neurology; Stage IIa - GCS of 15 plus focal neurology; Stage IIb - GCS of 11-14 with focal neurology; and Stage III - GCS <11.26,27 The importance of early diagnosis corresponding to an early TBM stage is emphasised by the significantly increased risk, as the disease progresses, of severe motor deficit and degree of intellectual disability in children classified as Stage IIb compared to those classified as Stage IIa.27 Stage III disease carries a 73% risk of the patient developing spastic quadriparesis and a mean developmental quotient of 44.27 Besides the prediction of disease outcome according to TBM stage, convulsions, headache, motor deficit, brainstem dysfunction and cerebral infarctions on neuroimaging were independently associated with poor clinical consequences of TBM in multivariate analysis.23
CSF diagnostic indicators of TBM
Cerebrospinal fluid findings are integral to the diagnosis of TBM. Typical CSF findings consist of leukocytosis with lymphocyte predominance, elevated protein and abnormally decreased CSF glucose (hypoglycorrhachia).28 Hypoglycorrhachia is indicated by either decreased CsF to plasma glucose ratio or reduced CSF glucose, and is unaffected by HIV co-infection.29 A uniform research case definition for TBM identified CSF criteria to aid in the diagnosis of TBM, including a CSF to serum glucose ratio of less than 0.5 or an absolute CSF glucose concentration of less than 2.2 mmol/L and elevated CSF protein of greater than 1 g/L.
There is potential for the use of CSF lactate in children with TBM, as CSF lactate has been shown to differentiate between bacterial and aseptic meningitis.30 CSF lactate levels, unaffected by serum lactate concentration, were significantly higher in adult TBM patients who subsequently died than in those who survived, reflecting the severity of cerebral hypoxia and therefore the overall prognosis.31 The diagnostic utility of CSF indicators considered in TBM has, however, rarely been described. Studies evaluating CSF IGRA (interferon-gamma release assay) showed good sensitivity and specificity,32 but the low CSF volumes in children are a limitation when sufficient cells are required to perform IGRA (typically 5-10 mL CSF is required).11 A study of the host immune response to M. tuberculosis showed the potential value of CSF interleukin-13, vascular endothelial growth factor and cathelicidin LL-37 as indicators when differentiating TBM from other forms of meningitis.18 Understanding the host immune response is key to a better understanding of the pathophysiology, the clinical presentation of TBM and the treatment of the disease. Novel indicators from CSF, including those from validated metabolomics studies, have the potential to be extremely useful as diagnostic tools in clinical practice.
NMR metabolomics methodology and applications
Metabolomics involves the simultaneous and comprehensive analysis of the measured responses of various biologically relevant small molecules, collectively referred to as the metabolome. The metabolome in turn is defined as the quantitative and qualitative collection of all low molecular weight molecules (metabolites, the end products of gene expression) of interest; they are found in concentrations varying in magnitude from picomoles to millimoles, they originate from endogenous or exogenous sources, and they exhibit a wide range of physico-chemical properties.33 These small molecules display characteristics which allow metabolomics to avoid several challenges associated with genomics and proteomics investigations. The great chemical complexity of the metabolome, however, means that a single analytical technique is not sufficient to provide comprehensive characterisation of all metabolites.33 Among the techniques available, NMR spectroscopy provides an attractive alternative to MS. Although it is less sensitive than its MS-based counterparts, NMR (1) allows for a robust and global look at the metabolome - it detects all classes of metabolites - making it non-biased; (2) is highly specific, allowing for detection of novel compounds; (3) running costs are markedly lower than that of other platforms, although initial set-up is expensive; (4) requires minimal sample preparation, and hence (5) does not chemically alter nor destroy the sample under investigation. NMR thus provides an excellent early or explorative view of the metabolome under investigation.34
The potential of NMR metabolomics in research on infectious human diseases has generated unique and novel insight into the underlying pathogenesis of these diseases and has unveiled new metabolic markers for disease diagnosis.35 Infectious diseases relevant to Africa that have been studied using NMR metabolomics include human African trypanosomiasis36, schistosomiasis37, malaria38, Leishmaniasis39, pneumonia40, and HIV41, as well as TB and TBM (discussed below).
NMR metabolomics of TB and TBM
The potential of metabolomics has been well documented, specifically with regard to TB.42 Its value is in the ability to (1) address simultaneously a wide array of metabolites from various different biochemical pathways; (2) detect and isolate patterns of disturbance for additional, targeted investigations; and (3) generate hypotheses to be tested. Godreuil et al.43 believe that the new quantitative and bioinformatics approaches to the study of the interaction between M. tuberculosis and the infected host, and how this interaction influences the infection process, are of particular importance, as it is fully accepted in the scientific description of infectious diseases that the outcome of transmission, infection and disease is dependent on the intrinsic characteristics of both the microbes and the host.
It is expected that at different stages of the M. tuberculosis life cycle - for example, dormancy, latent infection and active disease - there will be a different array of host- and M. tuberculosis-derived metabolites.42 These metabolic profiles can best be measured globally via NMR metabolomics (Table 1), which has been used successfully to provide insight into the metabolic changes in host response. In a pioneering NMR-based metabolomics pilot study of the CSF of patients with bacterial meningitis, Coen et al.47 detected: (1) elevated lactate and severe glucose depletion; (2) impairment of the citric acid cycle caused by reduced production of acetyl CoA, resulting in accumulation of pyruvate and generation of amino acids via transamination; (3) elevated CSF concentrations of pyruvate and amino acids - particularly alanine, isoleucine and leucine; and (4) the presence of ketone bodies - 3-hydroxybutyrate, acetoacetate and acetone - indicating compensatory response to glucose depletion and reduced ATP levels. Notably, the role of amino acid perturbation in meningitis is further supported by other studies.48 The metabolite profile described by Coen et al.47 for bacterial meningitis corresponds to recent MS-based results from our research centre on CSF from a TBM patient49 and to sputum from patients with pulmonary TB50. The pilot study of Coen et al.47 thus highlights the potential of NMR metabolomics in providing a global and unbiased view of metabolic or neurological perturbations in infectious diseases, particularly meningitis.
A study by Zhou et al.45 unambiguously identified 35 metabolites in sera of TB patients, of which 17 were altered. The majority of altered endogenous metabolites in the serum of TB patients were energy related - citric acid cycle intermediates, products of glycolysis, amino acids and indicators of enhanced lipid degradation - which is consistent with other studies. The metabolic processes found to be most significantly altered in TB patients were protein biosynthesis, followed by alanine metabolism, phenylalanine and tyrosine metabolism, and ammonia recycling. Zhou et al.45 go on to state that increased glycolytic products could be an index of tissue hypoxia and extent of necrosis as the infection progresses, that enhanced lipid degradation can be correlated with caseation of host TB granulomas, and that increased formate reflects an increased requirement for nucleotide biosynthesis, all of which indicates active host inflammatory cell division. A follow-up study by Zhou et al.46 highlighted the specificity of the metabolite profile of TB patients, compared to similar diseases such as representative metabolism-related diseases (diabetes mellitus), wasting diseases (malignancy), and lung inflammatory diseases (CAP). Consistent with their previous study, they found in the plasma: increased levels of lactate, pyruvate, lipids and ketone bodies, and decreased levels of glucose, glutamate, glutamine, glycerophosphocholine, very low-density lipoproteins and branched-chain amino acids. While Zhou et al.46 could distinguish TB patients statistically, the greatest statistical overlap occurred with the plasma profiles of patients with malignancy, indicating that, to some extent, the metabolism of TB mimics that of tumour cells. NMR metabolomics has also been used to elucidate the mechanism of action of specific medication, such as the second-line drug for TB, D-cycloserine, which is primarily used to treat MDR-TB.51
We have recently used a gas chromatography-mass spectrometry (GC-MS) approach to investigate the analytical repeatability of a CSF sample collected from a TBM patient for metabolomics analysis49 in order to develop a new method for evaluating repeatability in generating metabolomics data. This study clearly indicated the severe disease state of the TBM patient from whom the sample was obtained. We subsequently embarked on a comprehensive untargeted proton magnetic resonance (1H NMR)-based metabolomics analysis on lumbar CSF samples from three experimental groups: (1) South African infants and children with confirmed TBM; (2) treated, non-meningitis South African infants and children as controls; and (3) age-matched untreated controls from the Netherlands.52 Our primary focus was on identification of metabolites that distinguish between TBM and non-TBM cases; a total of 16 NMR-derived CSF metabolites was identified. These metabolites did not include cyclopropane, a putative marker for TBM defined in 2005.53 The defining two metabolites were the combination of perturbed glucose and highly elevated lactate, common to some other neurological disorders. Glucose, the primary source for energy production, is utilised in a rapid fashion, resulting in a significant increase in CSF lactate, both of which reflect a metabolic burst. The remaining 14 metabolites of the host's response to TBM were likewise mainly energy-associated indicators. Citrate and pyruvate, along with acetate in the form of acetyl-CoA, are key metabolites involved in the citric acid cycle as part of energy production. Alanine and the branched-chain amino acids - valine, leucine and isoleucine - have vital roles as shuttling systems transporting metabolites across cell membranes; they are also anaplerotic, replenishing citric acid cycle intermediates. Myo-inositol acts as an important signalling component necessary for immune responses such as microglia activation. The remaining other metabolites also have important roles; one such is dimethyl sulfone, which is an indicator of response to oxidative stress; another is choline, an important precursor to acetylcholine, which in excess is responsible for seizures.
The most conspicuous outcome of the NMR profiling was the high concentration of lactate in the TBM patients (7.36 mmol/L) relative to the two control groups (1.39 mmol/L and 1.70 mmol/L for the Dutch and South African controls, respectively). This observation has some clinical significance. It has been established54 that CSF lactate is applicable as a marker that can differentiate bacterial meningitis (>6 mmol/L) from other forms of meningitis, such as aseptic meningitis (<2 mmol/L). Thus, CSF concentrations of lactate, as well as immune biomarkers, are typically high before treatment and then decrease rapidly with therapy. In this regard a study55 on adults suffering from meningitis indicated that death was associated with high initial CSF concentrations of lactate and low numbers of white blood cells, in particular neutrophils. This outcome underscores the authors' proposal that CSF lactate is a good single indicator of the TBM disease state, making it better than conventional markers.
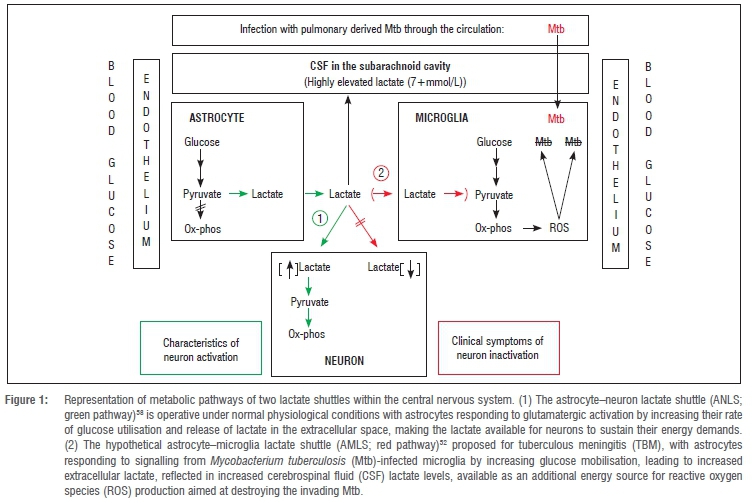
Furthermore, our NMR-profiling results52 provided a basis for hypothesis formulation of the pathogenic characteristics of TBM. We speculated that the inflammatory responses and metabolic imbalances created in the CNS following the initial phases of infection by a pathogen, such as M. tuberculosis, should advantage the microglia (the native macrophage in the brain) by fulfilling their energy-intensive immune-protective function. This speculation was made against the multi-paradigm background of the current literature and based on: (1) the shift in energy requirements as a result of the neuroinflammatory responses in TBM; (2) the notion that microglia play an important role in the neuropathogenesis of TBM; (3) recognition of the unique metabolic plasticity of cell-cell communication and regulation in CNS metabolism; and (4) the clinical profile on dysfunctional neuron activity seen in TBM. From observations and through inductive reasoning56 on the characteristics of the cell-cell interactions and factor isoforms, we advanced the following hypothesis: 'The host's response to neural infection results in an "astrocyte-microglia lactate shuttle" (AMLS) that operates in neuroinflammatory diseases, such as TBM', represented as a conceptual model detailed in the supplementary material of Mason et al.52 A simplified representation of the conceptual model is presented in Figure 1.
Briefly, it is postulated that, in TBM, lactate produced through glycolysis in astrocytes participates in the activated immune response and is directed from the neurons preferentially into microglia, where it enters the mitochondrial citric acid cycle, contributing to oxidative phosphorylation and hence producing high levels of adenosine triphosphate (ATP) and formation of reactive oxygen species (ROS) required for M. tuberculosis degradation. ROS, and a multitude of factors produced by the microglia to modulate the functions of surrounding immune cells, are toxic to neurons. Moreover, the unregulated activation of microglia in response to stimulants such as M. tuberculosis propagate neural injury57 and eventual apoptotic cell death for the over-activated microglia. By contrast, several investigations indicate that astrocytes could release large amounts of lactate under conditions of high energy demand by the neurons, leading to our hypothesis of an activity-dependent astrocyte-neuron lactate shuttle for the supply of energy substrates to neurons.58
From the preceding overview we highlight the following insights that have transpired from the NMR metabolomics of TB and TBM.
Advantages of NMR technology for clinical applications
(1) Non-invasive sample collection methods are often essential in clinical medicine, for which body fluids such as urine, cord blood and serum/plasma are particularly suited, with NMR spectroscopy providing a cornerstone for their metabolomics investigations. In addition, NMR spectroscopy can work with small sample volumes.
(2) NMR analyses require a minimum of sample pre-preparation and thereby prevent the occurrence of confounding analytical artefacts.
(3) Metabolomics produces large and complex data sets that cannot be interpreted through classic reductionist methods, but require an inductive, open-minded approach, aimed at hypothesis generation, which is philosophically a useful methodological approach in uncharted fields of research.
(4) Clinically, perturbations in metabolism are often difficult to recognise as the phenotypes may show considerable variation; however, NMR technology has provided a proven track record through the identification of inborn errors of metabolism, often already manifesting in neonates.
(5) The unique advantage of metabolomics technology is that it provides a dynamic view of host functional responses in health and disease, and offers early and rapid identification of the status and progress of a disease.
Cost-effectiveness of NMR technology
High-resolution NMR equipment, such as the instrument installed in early 2015 in a collaboration between the Technological Innovation Agency (TIA) and North-West University (NWU), is extremely expensive and requires highly skilled technical and scientific expertise for its operation. However, once an NMR facility has been established, the analytical clinical analysis is relatively inexpensive, costing only a few rands per sample to run. Moreover, the international trend in the market for miniaturisation of NMR instrumentation is well established, and is opening up new areas of study in academic and industrial settings with clear potential and appeal for future clinical practice.59
Systems insights
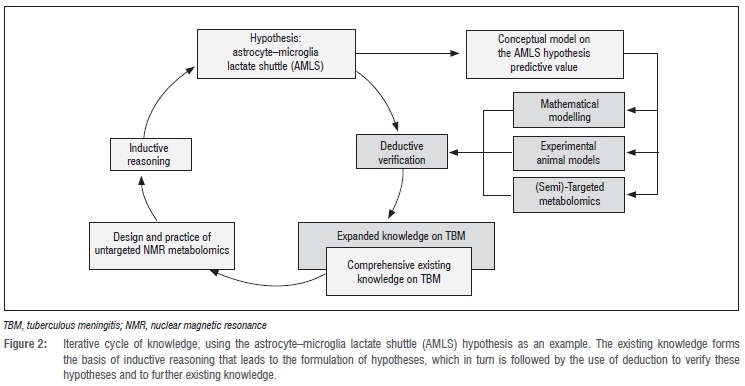
Fields that are data rich but hypothesis poor - prime examples are systems biology and many areas of medicine and clinical practice -are best approached through data-driven, essentially inductive methods. Biological systems are inherently complex; they require an inductive approach to data generation, data analysis and modelling if we are to continue to make strides in our understanding of these phenomena in health and disease. The advent of multidisciplinary interaction between biologists/clinicians, analytical chemists and informaticists/ biostatisticians in developing complex models has ushered in a new era in the mathematics of science, for the first time enabling understanding and prediction of large-scale biological systems; the 'omics' technologies are key in this process by being inductive and exploratory in their very essence. Intrinsic to this process remains the iterative relationship between the realm of ideas/hypotheses/thoughts (= induction) and that of observations/data/information (= deduction), linked in a repetitive cyclic way in which one arc is not simply the reverse of the other, but has methodological characteristics in its own right. We conclude our overview by focusing on this cyclic relationship.
Directives for hypothesis verification
Hypotheses are generalised statements derived from empirical evidence and prior knowledge of existing data, and they need to be verified or refuted by deductive methods (Figure 2). This activity can be represented as a circular process, with inductive reasoning forming one half of the iterative cycle of knowledge, and the second half (deduction) devoted to the aims of testing hypotheses and furthering existing knowledge. This representation highlights the essential nature of exploratory metabolomics investigations, and features three approaches that could be applied for hypothesis testing: (1) computational models, (2) animal models and (3) semi-targeted metabolomics.
Computational modelling
The extreme complexity of intracellular regulatory networks, involving genetic circuits, metabolic regulation and signal transduction, hampers the ability to analyse these functions quantitatively to achieve a holistic understanding of their role in health and disease. Complexity theory has thus become established over the last two decades as a novel discipline directed to such problems in the humanities and natural sciences, and is at present pursued in South Africa at the Centre for Studies in Complexity established in 2009 at Stellenbosch University. Modelling, in its various forms, demands key competencies in studying complexity. Models are used to describe complex systems through computer simulation based on quantitative information on many and key variables that characterise what is being investigated. An example is the computational model we recently developed - using in silico data that capture the unique immunological environment of the brain - which allows us to study the key mechanisms driving TB-linked granuloma formation in time.60 The model was based on a known pulmonary agent-based description of TB, representing the molecular signalling networks that affect granuloma formation during infection with M. tuberculosis. It focused on the formation of granulomas whose structure and function may reflect the success or failure of the host to contain infection. We envisage that computational model development, based on NMR data, may be a fruitful approach to use for understanding the dynamics involved in metabolic flux associated with the AMLS hypothesis, thereby permitting greater comprehension of the perturbations following M. tuberculosis infection of the microglia in the meninges.
Experimentation using animal models
Animal models have contributed significantly to knowledge of TB. However, there are certain major differences between humans and other animals in their responses to the disease. It is known that mice, for example, do not form the granulomas seen in humans and most other vertebrates with TB. Such differences caution our interpretations of data from animal models as they apply to human diseases. Nonetheless, animal models are indispensable to TB research and are well established in South African TB research centres.61,62 Until recently, for example, no murine model was available for experimental studies on TBM. We successfully developed a reproducible in vivo murine model to study the inflammatory response in TBM,63 and obtained useful insights despite limitations in translating the experimental observations to the human manifestation of the disease. Although the route of infection mimics the way TBM is acquired naturally, the cytokine profile in the mice does not show the clinical characteristics of the human disease. Nonetheless, the model has potential for further experimentation on M. tuberculosis infection because we observed that bacterial growth of TB in the CNS leads to a typical chronic inflammatory response. Such results indicate that experimental animal studies hold potential for investigating the biological changes in a M. tuberculosis-infected model at the biochemical level - an approach that could be expanded through appropriate knockout models, as has been shown for cryptococcal meningitis.64
Semi-targeted metabolomics
The metabolites present in the CSF represent the actual metabolism of the CNS and the balance between blood and CSF, rendering CSF analysis indispensable in the assessment of neurological disorders. Sampling human CSF is, however, an invasive procedure and ethical and safety constraints limit the availability of such samples for TBM, especially in work with infants and children. Thus, accessing these samples is much more difficult than collecting blood or urine. Metabolomics offers promising options. The use of MS-based metabolomics of plasma from patients suffering from propionic acidemia and methylmalonic acidemia,65 as well as our GC-MS metabolomics analysis of urine samples from patients with respiratory chain deficiencies66 and isovaleric acidemia67, have provided proof of concept that semi-targeted metabolomics can expand the range of metabolites associated with human disease. These studies also indicate the potential of metabolomics for non-invasive diagnosis and clinical evaluation of patients with TBM.
Perspective
Metabolomics studies of complex diseases are still in their infancy, although the volume of NMR- and MS-based metabolomics publications in particular is increasing exponentially.2 As it stands, NMR metabolomics is not a standalone method of diagnosis nor, by extension, of disease management of TB or of TBM, but it undeniably offers insights for potentially improving existing knowledge of infectious diseases in ways that may complement and benefit current diagnostic and treatment methods. Recent results, such as those by Zhou et al.46, strongly indicate that NMR metabolomics can contribute to improved understanding of disease mechanisms. However, a major gap in work on infectious diseases is still the paucity of subsequent validation of clinical studies, which is necessary to reap the full rewards of metabolomics technology. Ultimately, the optimal outcome, particularly in resource-limited settings, is the validation and declaration of simple and clear biomarkers for disease diagnosis - which requires appropriate and thorough follow-ups on metabolomics studies.
Fortunately, the great potential of biotechnology was recognised by the Department of Science and Technology, which enabled the creation of several technological platforms, including the Metabolomics Platform supported by the TIA and hosted by NWU on its Potchefstroom campus. This platform consists of a wide array of gas and liquid chromatography based MS instruments, as well as a 500-MHz NMR spectrometer dedicated to investigations of biofluids and other biological specimens. Furthermore, the contractual agreement between the TIA and NWU stipulates that: '...the Institution [NWU] agrees to have an open policy that encourages use of its facilities by researchers other than those based at the Institution...'. This policy of open access to the Metabolomics Platform aligns with the TIA's objective of stimulating and exploiting technological innovation in order to improve economic growth and the quality of life of all South Africans - a view that we strongly share, with the hope that this review will further promote these far-reaching ambitions.
Acknowledgements
Research funding for this project was provided by the Technological Innovation Agency of the Department of Science and Technology of South Africa. S.M. and R.S. are recipients of a Desmond Tutu-NRF-VU doctoral fellowship for a joint PhD study between the Vrije Universiteit in Amsterdam (the Netherlands), and North-West University and Stellenbosch University in South Africa.
Authors' contributions
S.M. and R.S. wrote the manuscript. C.J.R. and A.M.v.F. are the South African and Dutch project leaders, respectively, who directed the study and contributed to the writing of the manuscript.
References
1. Lindon JC, Nicholson JK, Holmes E. The handbook of metabonomics and metabolomics. London: Elsevier Science; 2011. [ Links ]
2. Goodacre R. An overflow of... what else but metabolism! Metabolomics. 2010;6:1-2. http://dx.doi.org/10.1007/s11306-010-0201-3 [ Links ]
3. Rieder HL, Kelly GD, Bloch AB, Cauthen GM, Snider DE. Tuberculosis diagnosed at death in the United States. Chest. 1991;100:678-681. http://dx.doi.org/10.1378/chest.100.3.678 [ Links ]
4. Perez-Velez CM, Marais BJ. Tuberculosis in children. New Engl J Med. 2012;367(4):348-361. http://dx.doi.org/10.1056/NEJMra1008049 [ Links ]
5. Bhigjee AI, Padayachee R, Paruk H, Hallwirth-Pillay KD, Marais S, Connoly C. Diagnosis of tuberculous meningitis: Clinical and laboratory parameters. Int J Infect Dis. 2007;11:248-254. http://dx.doi.org/10.1016/j.ijid.2006.07.007 [ Links ]
6. Wolzak NK, Cooke ML, Orth H, Van Toorn R. The changing profile of pediatric meningitis at a referral centre in Cape Town, South Africa. J Trop Pediatr. 2012;58(6):491-495. http://dx.doi.org/10.1093/tropej/fms031 [ Links ]
7. Donald PR, Cotton MF, Hendricks MK, Schaaf HS, De Villiers JN, Willemse TE. Pediatric meningitis in the Western Cape Province of South Africa. J Trop Pediatr. 1996;42:256-261. http://dx.doi.org/10.1093/tropej/42.5.256 [ Links ]
8. Van Rie A, Beyers N, Gie RP Kunneke M, Zietsman L, Donald PR. Childhood tuberculosis in an urban population in South Africa: Burden and risk factor. Arch Dis Child. 1999;80:433-437. http://dx.doi.org/10.1136/adc.80.5.433 [ Links ]
9. Thwaites GE, Van Toorn R, Schoeman J. Tuberculous meningitis: More questions, still too few answers. Lancet Neurol. 2013;12:999-1010. http://dx.doi.org/10.1016/S1474-4422(13)70168-6 [ Links ]
10. Chiang SS, Khan FA, Milstein MB, Tolman AW, Benedetti A, Starke JR, et al. Treatment outcomes of childhood tuberculous meningitis: A systematic review and meta-analysis. Lancet Infect Dis. 2014;14:947-957. http://dx.doi.org/10.1016/S1473-3099(14)70852-7 [ Links ]
11. Van Toorn R, Solomons R. Update on the diagnosis and management of tuberculous meningitis in children. Semin Pediatr Neurol. 2014;21:12-18. http://dx.doi.org/10.1016/j.spen.2014.01.006 [ Links ]
12. Hektoen L. The vascular changes of tuberculous meningitis, especially tuberculous endarteritis. J Exp Med. 1896;1:112-163. http://dx.doi.org/10.1084/jem.1.1.112 [ Links ]
13. Rich AR, McCordock HA. The pathogenesis of tuberculous meningitis. Bull Johns Hopkins Hosp. 1933;52:2-37. [ Links ]
14. Donald PR, Schaaf HS, Schoeman JF. Tuberculous meningitis and miliary tuberculosis: The Rich focus revisited. J Infect. 2005;50(3):193-195. http://dx.doi.org/10.1016/j.jinf.2004.02.010 [ Links ]
15. Thwaites GE, Schoeman JF. Update on tuberculosis of the central nervous system: Pathogenesis, diagnosis, and treatment. Clin Chest Med. 2009;30(4):745-754. http://dx.doi.org/10.1016/j.ccm.2009.08.018 [ Links ]
16. Janse van Rensburg P, Andronikou S, Van Toorn R, Pienaar M. Magnetic resonance imaging of miliary tuberculosis of the central nervous system in children with tuberculous meningitis. Pediatr Radiol. 2008;38(12):1306-1313. http://dx.doi.org/10.1007/s00247-008-1028-1 [ Links ]
17. Horsburgh CR Jr, Rubin EJ. Clinical practice. Latent tuberculosis infection in the United States. New Engl J Med. 2011;364(15):1441-1448. http://dx.doi.org/10.1056/NEJMcp1005750 [ Links ]
18. Visser DH, Solomons RS, Ronacher K, Van Well GT, Heymans MW, Walzl G, et al. Host immune response to tuberculous meningitis. Clin Infect Dis. 2014;60(2):177-187. http://dx.doi.org/10.1093/cid/ciu781 [ Links ]
19. Kashyap RS, Deshpande PS, Ramteke SR, Panchbhai MS, Purohit HJ, Taori GM, et al. Changes in cerebrospinal fluid cytokine expression in tuberculous meningitis patients with treatment. Neuroimmunomodulation. 2010;17:333-339. http://dx.doi.org/10.1159/000292023 [ Links ]
20. Patel VB, Singh R, Connolly C, Kasprowicz V Ndung'u T, Dheda K. Comparative utility of cytokine levels and quantitative RD-1-specific T cell responses for rapid immunodiagnosis of tuberculous meningitis. J Clin Microbiol. 2011;49(11):3971-3976. http://dx.doi.org/10.1128/JCM.01128-11 [ Links ]
21. Donald PR, Schoeman JF, Beyers N, Nel ED, Carlini SM, Olsen KD, et al. Concentrations of interferon γ, tumor necrosis factor α, and interleukin-1 /3 in the cerebrospinal fluid of children treated for tuberculous meningitis. Clin Infect Dis. 1995;21(4):924-929. http://dx.doi.org/10.1093/clinids/21.4.924 [ Links ]
22. Schoeman JF, Wait J, Burger M, Van Zyl F, Fertig G, Janse van Rensburg A, et al. Long-term follow-up of childhood tuberculous meningitis. Dev Med Child Neurol. 2002;44(8):522-526. http://dx.doi.org/10.1111/j.1469-8749.2002.tb00323.x [ Links ]
23. Van Well GT, Paes BF, Terwee CB, Springer P Roord JJ, Donald PR, et al. Twenty years of pediatric tuberculous meningitis: A retrospective cohort study in the Western Cape of South Africa. Pediatrics. 2009;123(1):e1-e8. http://dx.doi.org/10.1542/peds.2008-1353 [ Links ]
24. Seddon JA, Visser DH, Bartens M, Jordaan AM, Victor TC, Van Furth AM, et al. Impact of drug resistance on clinical outcome in children with tuberculous meningitis. Pediatr Infect Dis J. 2012;31(7):711-716. http://dx.doi.org/10.1097/INF.0b013e318253acf8 [ Links ]
25. Van Toorn R, Schaaf HS, Laubscher JA, Van Elsland SL, Donald PR, Schoeman JF. Short intensified treatment in children with drug-susceptible tuberculous meningitis. Pediatr Infect Dis J. 2014;33(3):248-252. http://dx.doi.org/10.1097/INF.0000000000000065 [ Links ]
26. Van Toorn R, Springer P Laubscher JA, Schoeman JF. Value of different staging systems for predicting neurological outcome in childhood tuberculous meningitis. Int J Tuberc Lung Dis. 2012;16(5):628-632. [ Links ]
27. British Medical Research Council. Streptomycin treatment of tuberculous meningitis. Br Med J. 1948;1(6503):582-596. [ Links ]
28. Thwaites G, Fisher M, Hemingway C, Scott G, Solomon T, Innes J. British Infection Society guidelines for the diagnosis of tuberculosis of the central nervous system in adults and children. J Infect. 2009;59:167-187. http://dx.doi.org/10.1016/j.jinf.2009.06.011 [ Links ]
29. Marais S, Thwaites G, Schoeman JF, Török ME, Misra UK, Prasad K, et al. Tuberculous meningitis: A uniform case definition for use in clinical research. Lancet Infect Dis. 2010;10(11):803-812. http://dx.doi.org/10.1016/S1473-3099(10)70138-9 [ Links ]
30. Huy NT, Thao NT, Diep DT, Kikuchi M, Zamora J, Hirayama K. Cerebrospinal fluid lactate concentration to distinguish bacterial from aseptic meningitis: A systemic review and meta-analysis. Crit Care. 2010;14(6), R240, 15 pages. http://dx.doi.org/10.1186/cc9395 [ Links ]
31. Thwaites GE, Simmons CP Than Ha Quyen N, Thi Hong Chau N, Phuong Mai P Thi Dung N, et al. Pathophysiology and prognosis in Vietnamese adults with tuberculous meningitis. J Infect Dis. 2003;188:1105-1115. http://dx.doi.org/10.1086/378642 [ Links ]
32. Thwaites GE. Advances in the diagnosis and treatment of tuberculous meningitis. Curr Opin Neurol. 2013;26(3):295-300. http://dx.doi.org/10.1097/WCO.0b013e3283602814 [ Links ]
33. Dunn WB, Ellis DI. Metabolomics: Current analytical platforms and methodologies. Trends Anal Chem. 2005;24(4):285-294. http://dx.doi.org/10.1016/j.trac.2004.11.021 [ Links ]
34. Kell DB, Oliver SG. Here is the evidence, now where is the hypothesis? The complementary roles of inductive hypothesis-driven science in the post-genomics era. BioEssays. 2003;66:99-105. [ Links ]
35. Duarte IF, Diaz SO, Gil AM. NMR metabolomics of human blood and urine in disease research. J Pharm Biomed Anal. 2014;93:17-26. http://dx.doi.org/10.1016/j.jpba.2013.09.025 [ Links ]
36. Wang Y Utzinger J, Saric J, Li JV Burckhardt J, Dirnhofer S, et al. Global metabolic responses of mice to Trypanosoma brucei brucei infection. Proc Natl Acad Sci USA. 2008;105(16):6127-6132. http://dx.doi.org/10.1073/pnas.0801777105 [ Links ]
37. Wang Y Holmes E, Nicholson JK, Cloarec O, Chollet J, Tanner M, et al. Metabonomic investigations in mice infected with Schistosoma mansoni: An approach for biomarker identification. Proc Natl Acad Sci USA. 2004;101(34):12676-12681. http://dx.doi.org/10.1073/pnas.0404878101 [ Links ]
38. Li JV Wang Y Saric J, Nicholson JK, Dirnhofer S, Singer BH, et al. Global metabolic responses of NMRI mice to an experimental Plasmodium berghei infection. J Proteome Res. 2008;7(9):3948-3956. http://dx.doi.org/10.1021/pr800209d [ Links ]
39. Gupta N, Goyal N, Singha UK, Bhakuni V Roy R, Rastogi AK. Characterization of intracellular metabolites of axenic amastigotes of Leishmania donovani by 1H NMR spectroscopy. Acta Trop. 1999;73(2):121-133. http://dx.doi.org/10.1016/S0001-706X(99)00020-0 [ Links ]
40. Slupsky CM, Rankin KN, Fu H, Chang D, Rowe BH, Charles PG, et al. Pneumococcal pneumonia: Potential for diagnosis through a urinary metabolic profile. J Proteome Res. 2009;8(12):5550-5558. http://dx.doi.org/10.1021/pr9006427 [ Links ]
41. Hewer R, Vorster J, Steffens FE, Meyer D. Applying biofluid 1H NMR-based metabonomic techniques to distinguish between HIV-1 positive/AIDS patients on antiretroviral treatment and HIV-1 negative individuals. J Pharm Biomed Anal. 2006;41(4):1442-1446. http://dx.doi.org/10.1016/j.jpba.2006.03.006 [ Links ]
42. Parida SK, Kaufmann SHE. The quest for biomarkers in tuberculosis. Drug Discov Today. 2010;15(3/4):148-157. http://dx.doi.org/10.1016/j.drudis.2009.10.005 [ Links ]
43. Godreuil S, Tazi L, Banuls AL. Pulmonary tuberculosis and mycobacterium tuberculosis: modern molecular epidemiology and perspectives. In: Tibayrenc M, editor. Encyclopedia of infectious diseases: Modern methodologies. Hoboken, NJ: John Wiley & Sons; 2007. http://dx.doi.org/10.1002/9780470114209.ch1 [ Links ]
44. Shin JH, Yang JY Jeon BY Yoon YJ, Cho SN, Kang YH, et al. 1H NMR-based metabolomic profiling in mice infected with Mycobacterium tibercilosis. J Proteome Res. 2011;10(5):2238-2247. http://dx.doi.org/10.1021/pr101054m [ Links ]
45. Zhou A, Ni J, Xu Z, Wang Y Lu S, Sha W, et al. Application of 1H NMR spectroscopy-based metabolomics to sera of tuberculosis patients. J Proteome Res. 2013;12(10):4642-l649. http://dx.doi.org/10.1021/pr4007359 [ Links ]
46. Zhou A, Ni J, Xu Z, Wang Y Zhang W, Wu W, et al. Metabolomics specificity of tuberculosis plasma revealed by 1H NMR spectroscopy. Tuberculosis. 2015;95:294-302. http://dx.doi.org/10.1016/j.tube.2015.02.038 [ Links ]
47. Coen M, O'Sullivan M, Bubb WA, Kuchel PW, Sorrell T. Proton nuclear magnetic resonance-based metabonomics for rapid diagnosis of meningitis and ventriculitis. Clin Infect Dis. 2005;41(11):1582-1590. http://dx.doi.org/10.1086/497836 [ Links ]
48. Qureshi GA, Baig SM, Bednar I, Halawa A, Parvez SH. The neurochemical markers in cerebrospinal fluid to differentiate between aseptic and tuberculous meningitis. Neurochem Int. 1998;32:197-203. http://dx.doi.org/10.1016/S0197-0186(97)00061-2 [ Links ]
49. Mason S, Moutloatse GP, Van Furth AM, Solomons R, Van Reenen M, Reinecke CJ, et al. KEMREP: A new qualitative method for the assessment of an analyst's ability to generate a metabolomics data matrix by gas chromatography- mass spectrometry. Curr Metabolomics. 2014;2(1):15-26. http://dx.doi.org/10.2174/2213235X02666140115214427 [ Links ]
50. Du Preez I, Loots DT. New sputum metabolite markers implicating adaptations of the host to Mycobacterium tuberculosis, and vice versa. Tuberculosis. 2013;93:330-337. http://dx.doi.org/10.1016/j.tube.2013.02.008 [ Links ]
51. Halouska S, Chacon O, Fenton RJ, Zinniel DK, Barletta RG, Powers R. Use of NMR metabolomics to analyze the targets of D-cycloserine in mycobacteria: Role of D-alanine racemase. J Proteome Res. 2007;6(12):4608-4614. http://dx.doi.org/10.1021/pr0704332 [ Links ]
52. Mason S, Van Furth AM, Mienie LJ, Engelke UFH, Wevers RA, Solomons R, et al. A hypothetical astrocyte-microglia lactate shuttle derived from a 1H NMR metabolomics analysis of cerebrospinal fluid from a cohort of South African children with tuberculous meningitis. Metabolomics. 2015;11:822-837. http://dx.doi.org/10.1007/s11306-014-0741-z [ Links ]
53. Subramanian A, Gupta A, Saxena S, Gupta A, Kumar R, Nigam A, et al. Proton MR CSF analysis and a new software as predictors of the differentiation of meningitis in children. NMR Biomed. 2005;18:213-225. http://dx.doi.org/10.1002/nbm.944 [ Links ]
54. Cunha BA. Distinguishing bacterial from viral meningitis: The critical importance of the CSF lactic acid levels. Intensive Care Med. 2006;32(8):1272-1273. http://dx.doi.org/10.1007/s00134-006-0210-x [ Links ]
55. Genton B, Berger JP. Cerebrospinal fluid lactate in 78 cases of adult meningitis. Intensive Care Med. 1990;16(3):196-200. http://dx.doi.org/10.1007/BF01724802 [ Links ]
56. Goodacre R, Vaidyanathan S, Dunn WB, Harrigan GG, Kell DB. Metabolomics by numbers: Acquiring and understanding global metabolite data. Trends Biotechnol. 2004;22:245-252. http://dx.doi.org/10.1016/j.tibtech.2004.03.007 [ Links ]
57. Block ML, Hong JS. Microglia and inflammation-mediated neurodegeneration: Multiple triggers with a common mechanism. Prog Neurobiol. 2005;76:77-98. http://dx.doi.org/10.1016/j.pneurobio.2005.06.004 [ Links ]
58. Pellerin L, Pellegri G, Bittar PG, Charnay Y Bouras C, Martin JL, et al. Evidence supporting the existence of an activity-dependent astrocyte-neuron lactate shuttle. Dev Neurosci. 1998;20:291-299. http://dx.doi.org/10.1159/000017324 [ Links ]
59. Lee H, Sun E, Ham D, Weissleder R. Chip-NMR biosensor for detection and molecular analysis of cells. Nat Med. 2008;14:869-874. http://dx.doi.org/10.1038/nm.1711 [ Links ]
60. El-Kebir M, Van der Kuip M, Van Furth AM, Kirschner DE. Computational modeling of tuberculous meningitis reveals an important role for tumor necrosis factor-α. J Theor Biol. 2013;328:43-53. http://dx.doi.org/10.1016/j.jtbi.2013.03.008 [ Links ]
61. De Souza GA, Fortuin S, Aguilar D, Pando RH, McEvoy CR, Van Helden PD, et al. Using a label-free proteomics method to identify differentially abundant proteins in closely related hypo- and hypervirulent clinical Mycobacterium tuberculosis Beijing isolates. Mol Cell Proteomics. 2010;9(11):2414-2423. http://dx.doi.org/10.1074/mcp.M900422-MCP200 [ Links ]
62. Drennan MB, Nicolle D, Quesniaux VJ, Jacobs M, Allie N, Mpagi J, et al. Toll-like receptor 2-deficient mice succumb to Mycobacterium tuberculosis infection. Am J Pathol. 2004;164(1):49-57. http://dx.doi.org/10.1016/S0002-9440(10)63095-7 [ Links ]
63. Van Well GTJ, Wieland CW, Florquin S, Roord JJ, Van der Poll T, Van Furth AM. A new murine model to study the pathogenesis of tuberculous meningitis. J Infect Dis. 2007;195:694-697. http://dx.doi.org/10.1086/511273 [ Links ]
64. Lee A, Toffaletti DL, Tenor J, Soderblom EJ, Thompson JW, Moseley MA, et al. Survival defects of Cryptococcus neoformans mutants exposed to human cerebrospinal fluid result in attenuated virulence in an experimental model of meningitis. Infect Immun. 2010;78(10):4213-4225. http://dx.doi.org/10.1128/IAI.00551-10 [ Links ]
65. Wikoff WR, Gangoiti JA, Barshop BA, Siuzdak G. Metabolomics identifies perturbations in human disorders of propionate metabolism. Clin Chem. 2007;53:2169-2176. http://dx.doi.org/10.1373/clinchem.2007.089011 [ Links ]
66. Reinecke CJ, Koekemoer G, Van der Westhuizen FH, Louw R, Lindeque JZ, Mienie LJ, et al. Metabolomics of urinary organic acids in respiratory chain deficiencies in children. Metabolomics. 2012;8(2):264-283. http://dx.doi.org/10.1007/s11306-011-0309-0 [ Links ]
67. Dercksen M, Duran M, Ijlst L, Mienie LJ, Reinecke CJ, Ruiter JPN, et al. Clinical variability of isovaleric acidemia in a genetically homogeneous population. J Inherit Metab Dis. 2012;35(6):1021-1029. http://dx.doi.org/10.1007/s10545-012-9457-2 [ Links ]
 Correspondence:
Correspondence:
Carolus Reinecke
Centre for Human Metabonomics, Faculty of Natural Sciences
Private Bag X6001, Potchefstroom 2531. South Africa
carools.reinecke@nwu.ac.za
Received: 03 Mar. 2015
Revised: 03 Aug. 2015
Accepted: 31 Oct. 2015


{kind=link}
{kind=link}
{kind=link}