










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.112 no.1-2 Pretoria Jan./Fev. 2016
http://dx.doi.org/10.17159/sajs.2016/20150227
RESEARCH ARTICLE
Spoilage potential of a novel group of bacteria isolated from dairy products
L. Ingrid TsôeuI; Piet J. JoosteII; George CharimbaI; Celia J. HugoI
IDepartment of Microbial, Biochemical and Food Biotechnology, University of the Free State, Bloemfontein, South Africa
IIDepartment of Biotechnology and Food Technology, Tshwane University of Technology, Pretoria, South Africa
ABSTRACT
Cold-tolerant bacteria, also known as psychrotrophic bacteria, are notorious contaminants of milk in the refrigerated dairy food chain. These organisms, especially the pseudomonads, may produce heat-resistant enzymes that are responsible for the breakdown of proteins and lipids in milk and dairy products. Such reactions result in a variety of defects in the raw or unprocessed milk that may affect the suitability of such milk for further processing. The enzymes produced may cause defects in long-life dairy products such as cheese, butter and long-life milk. In the present study, a range of 18 yellow pigmented psychrotrophic bacteria, collectively known as flavobacteria, were isolated from local dairy products. One aim of this study was to identify these bacteria to species level using molecular techniques. A second aim was to determine the spoilage potential of these organisms based on profiles generated by the BIOLOG system (that may relate to hydrolytic enzymes produced). Of the 18 isolates, 14 belonged to the genus Chryseobacterium while 4 were identified as Empedobacter isolates. The most active spoilage organisms in this group were shown to be C. bovis, C. shigense and E. brevis. These findings illustrate that enzymatically catalysed defects in dairy products should not be attributed solely to acknowledged psychrotrophic bacteria such as the pseudomonads, but that flavobacterial species may also be actively involved.
Keywords: identification; spoilage; Chryseobacterium, Empedobacter; dairy
Introduction
Extended cold storage times of raw milk at 2-6 °C have a significant influence on the natural microbial population. At these temperatures, the predominantly Gram-positive mesophilic aerobic bacteria are gradually replaced by Gram-negative and Gram-positive psychrotrophic bacteria.1,2 Psychotrophic bacteria are able to grow at 7 °C or less regardless of their optimal growth temperature.3 In addition to this ability, psychrotrophic bacteria have the ability to produce heat stable extracellular and/or intracellular hydrolytic enzymes which may retain their activity even after the conventional heat treatment of milk.4
The genera Pseudomonas, Aeromonas, Serratia, Acinetobacter, Alcaligenes, Achromobacter, Enterobacter and Flavobacterium, with the predominance of Pseudomonas as the most frequent representative of Gram-negative psychrotrophic bacteria, have been isolated from raw milk.5 Yellow pigmented psychrotrophs included in the bacterial family Flavobacteriaceae have the potential to decompose milk and dairy products.6 These flavobacteria produce proteolytic enzymes that may survive pasteurisation7-9 and cause bitter flavours, gelation of long-life heat treated milk10 and the production of off-odours.11-13 Jooste and Britz8 found that the practical importance of dairy flavobacteria lies as much in their psychrotrophic growth and consequent proteinase production in refrigerated milk as in their contamination of milk via poorly sanitised pipelines and equipment.
While the taxonomy of the so-called flavobacterial group was in disarray for many years, this situation has stabilised14,15 and more recently 114 genera have been included in the Flavobacteriaceae family.16 Of the 114 genera in this family, 10 are associated with food, namely Bergeyella, Chryseobacterium, Empedobacter, Flagellimonas, Flavobacterium, Myroides, Salegentibacter, Tenacibaculum, Vitellibacter and Weeksella.17 The latter reclassification and, in some cases erroneous classification of the flavobacteria in the past, makes interpretation of information about the incidence and role of flavobacteria in food deterioration a challenging task.18,19
Because of the latter changes in the flavobacterial taxonomy, it was decided to re-evaluate flavobacterial isolates from a previous study.14 The aims of the study were therefore to more adequately identify flavobacterial strains isolated from dairy sources by 16S rRNA sequencing and phenotypic characterisation and then to estimate or predict the potential spoilage characteristics of the isolates by utilisation of the considerable range of substrates in the BIOLOG system.
Materials and methods
Revival of freeze-dried dairy isolates
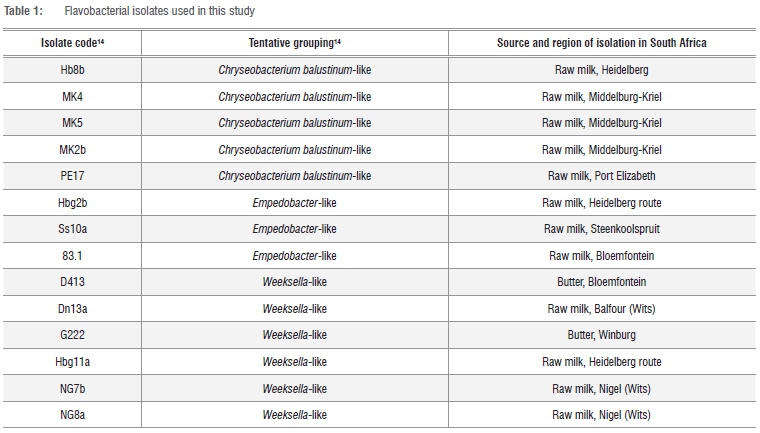
The 18 isolates used in this study were isolated in a previous study, from raw cow's milk and butter samples from diverse regions of South Africa.14 (Table 1). The isolates were preserved in the freeze-dried state and maintained at -20 °C. These freeze-dried cultures were revived by inoculation into nutrient broth (NB; Oxoid CM67, Thermoscientific, Basingstoke, United Kingdom) and incubated for 48 h at 25 °C. This was followed by streaking onto nutrient agar (NA; Oxoid CM003; Thermoscientific) and incubation for 48 h at 25 °C. Gram staining, oxidase and catalase tests20 were performed on pure cultures to verify that the isolates were Gram-negative and oxidase and catalase positive, before further analysis.
16S rRNA sequencing
The DNA of the 18 isolates was extracted as previously described.21 Polymerase chain reaction (PCR) amplification of the 16S rRNA gene was performed using an Eppendorf Mastercycler Temperature Gradient Personal Thermal cycler (Applied Biosystems, Johannesburg, South Africa) and the forward, 27F (5'-GAGTTTGATCCTGGCTCAG-3') and reverse, 1492R (5'-GGTTACCTTGTTACGACTT-3') primers.22 The PCR product was visualised on a 1% (w/v) agarose gel.
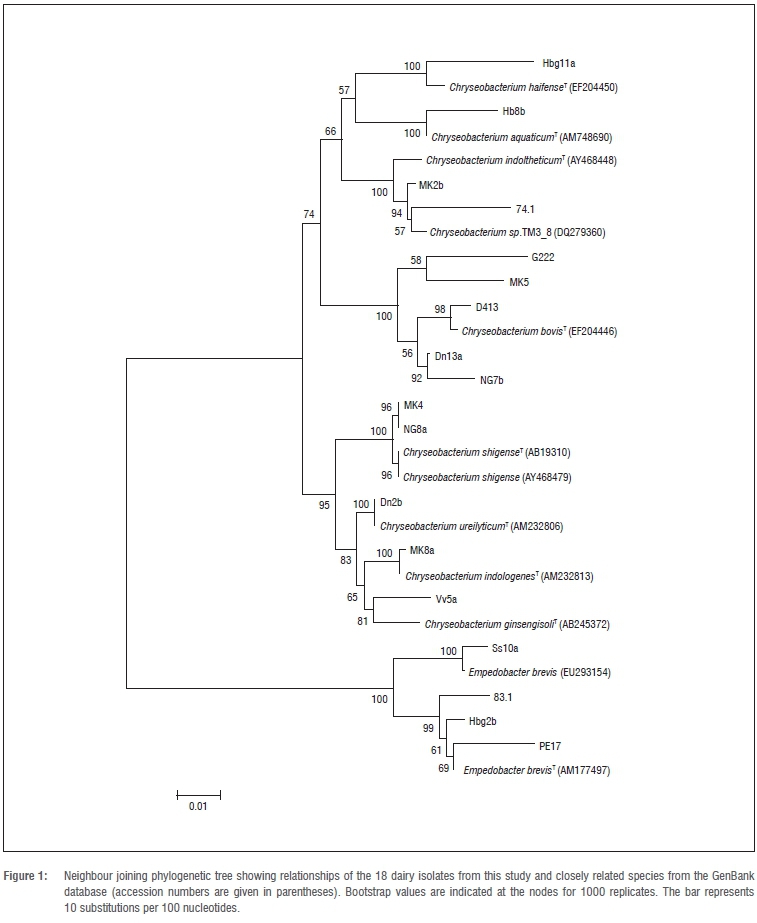
The 1.5 kb bands obtained were excised from the agarose gel and purified using the DNA clean and concentrator kit according to the manufacturer's protocol (Zymo Research, Irvine, CA, USA). Sequencing was performed using both the forward and reverse primers with an ABI BigDye® Terminator v1.1 sequence cycler (Applied Biosystems, Johannesburg, South Africa) according to the manufacturer's instructions. The newly determined 16S rRNA sequences (ca. 1500 bp) were compared to those available on the NCBI GenBank database23 using the Basic Local Alignment Search Tool (BLAST). A phylogenetic tree was constructed using the neighbour-joining method in the MEGA 4 software24 to determine the relationship of isolates in this study with those on the NCBI GenBank database.
Phenotypic characterisation
In order to support the data obtained from the 16S rRNA sequencing, a range of phenotypic tests19,20,25 were conducted using 24 h-old cultures on Na at 25 °C. The tests included pigment production, motility (flagellar type), growth in 0-5% (w/v) sodium chloride, growth at 5 °C, 25 °C, 37 °C and 42 °C; growth on β-hydroxybutyrate agar and MacConkey agar (Oxoid CM115, Thermoscientific); production of catalase, oxidase, phosphatase, DNase, urease, indole (Kovacs reagent, Merck 1.09293), β-galactosidase (ONPG) and reduction of nitrate; hydrolysis of esculin, gelatine (tube method) and starch; and acid production from carbohydrates in ammonium salts medium containing the following sugars: arabinose, cellobiose, D-fructose, D-glucose, lactose, maltose, mannitol, rhamnose, sorbitol, sucrose, trehalose, and D-xylose.
Estimation of potential spoilage
For the estimation and prediction of potential spoilage characteristics, the isolates were streaked out and incubated at 25 °C for 24 h. The isolates were inoculated onto BIOLOG GN2 microplates (BIOLOG Inc., Hayward, California) according to the manufacturer's protocol. The 96-welled microplates contain 96 different oxidisable carbon sources which could be grouped into carbohydrates, amino acids, carboxylic acids and polymers and which were used for the estimation of the potential spoilage characteristics.
Results and discussion
Identification of isolates
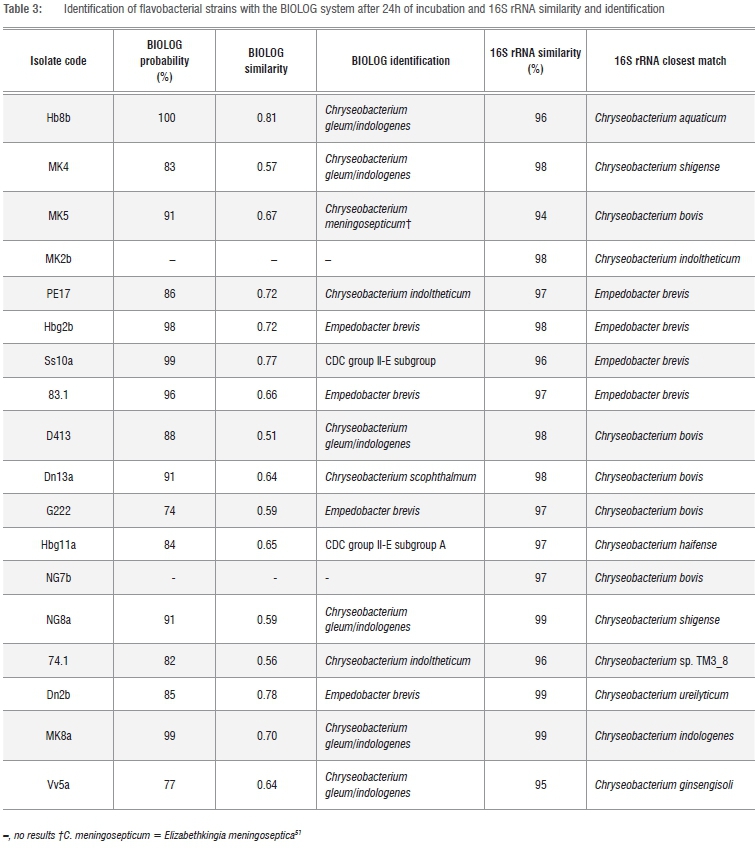
The results of the 16S rRNA sequencing indicated that 14 of the 18 dairy isolates belonged to the Chryseobacterium genus with sequence similarities between 94% and 99%, while four isolates belonged to the Empedobacter genus with sequence similarities between 96% and 98%. The phylogenetic relationships of the isolates to the type strains of Chryseobacterium species and Empedobacter brevis are shown in Figure 1.
Isolate Hb8b was closely associated with the type strain of C. aquaticum which was originally isolated from a water reservoir.26 Isolate Hbg11a was closely associated with the type strain of C. haifense while five isolates (G222, MK5, D413, Dn13a and NG7b) were identified as C. bovis. The type strains of both C. haifense and C. bovis were isolated from raw milk in Israel.27,28
Two isolates (MK2b and 74.1) were identified as C. indoltheticum. The type strain of C. indoltheticum was originally isolated from marine mud.29 Isolates MK4 and NG8a were identified as C. shigense. The type strain of C. shigense was originally isolated from a lactic acid beverage in Japan.30 Isolate Dn2b was identified as C. ureilyticum. The type strain of this species was originally isolated from a beer bottling plant.31 One isolate (MK8a) was identified as C. indologenes. Although C. indologenes is also known as a human pathogen, strains of this species have been isolated from food and the environment in previous studies.11,18,20
Although isolate Vv5a grouped closely with C. ginsengisoli, it could not be assigned to this species because of the low bootstrap value of 81 (Figure 1) when compared to the type strain. These data may suggest that isolate Vv5a could represent a new species. Isolates Ss10a, 83.1, Hbg2b and PE17 were identified as Empedobacter brevis.
Phenotypic analyses supported the 16S rRNA sequencing results by demonstrating typical phenotypic characteristics of the genera Chryseobacterium and Empedobacter.21 All isolates were strictly aerobic, Gram-negative rods with yellow flexirubin or carotenoid pigments. All isolates were positive for the production of catalase, oxidase, phosphatase, DNase; growth at 25 °C and growth in 0 and 1% NaCl. All the isolates were negative for motility; growth in 5% NaCl and acid production from arabinose, cellobiose, rhamnose, sorbitol and xylose. The differential characteristics of the genera and species identified in this study are shown in Table 2.
Although the BIOLOG method can also be used for identification purposes, the only flavobacteria that formed part of the BIOLOG database were C. gleum/indologenes, C. meningosepticum, C. indoltheticum, C. scophthalmum, E. brevis and CDC group II-E. The BIOLOG method could therefore only confirm that the isolates belonged to the Flavo-bacteriaceae family with grouping in either the Chryseobacterium or the Empedobacter genera (Table 3).
Potential spoilage characteristics
Utilisation of carbohydrates
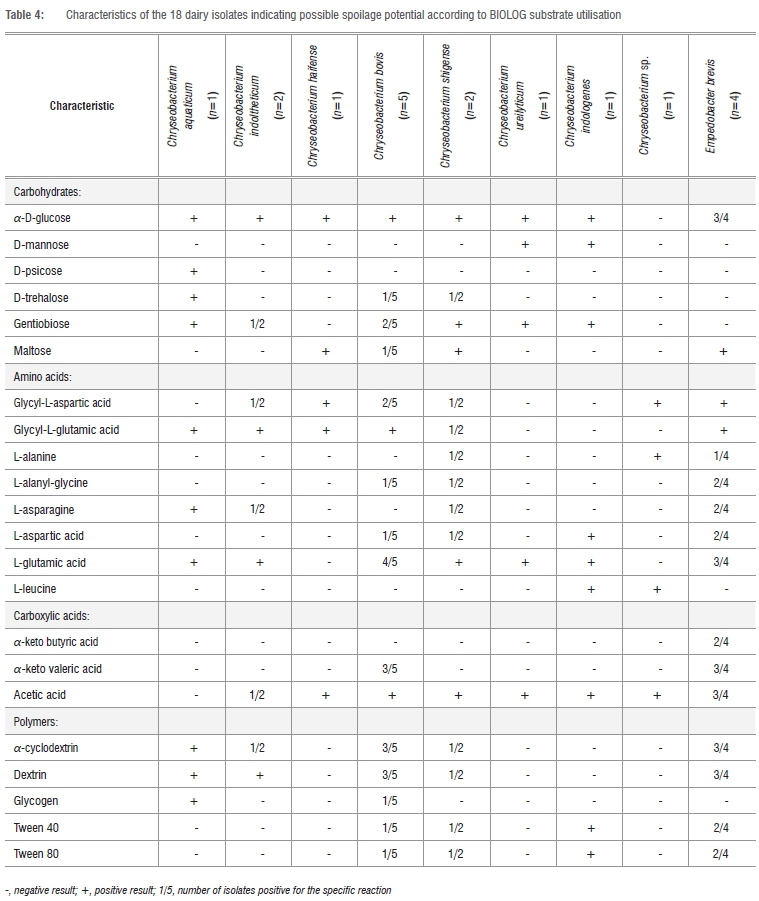
In this study, all the Chryseobacterium isolates (except Chryseobacterium strain Vv5a) and most of the Empedobacter isolates utilised glucose (Table 4). The other carbohydrates were used in varying degrees by specific isolates. Bacterial types that are capable of oxidising a specific carbohydrate will use it as a carbon source to support growth. In the process, the carbohydrate is oxidised to CO2 and water and, therefore, should not contribute to food spoilage. Only when the supply of simple, utilisable carbohydrates has been exhausted, will recognisable off-odours develop. These off-odours arise from the formation, and increase in concentration, of a variety of malodorous substances that include ammonia, amines, dimethylsulphide and hydrogen sulphide from amino acids.32,33
Interestingly, Chryseobacterium strain Vv5a did not utilise any of the carbohydrates tested in this study. This was in accordance with the 16S rRNA sequencing results, which showed that isolate Vv5a did not cluster with any of the type strains tested. These results lend impetus to placing the strain in a new species.
Utilisation of amino acids
All the Chryseobacterium species (except strain Vv5a) and E. brevis isolates tested in this study were able to utilise glycyl-L-glutamic acid and/or L-glutamic acid (Table 4). The second most utilised amino acid was glycyl-L-aspartic acid. The oxidation/reduction of glutamic acid may yield acetic acid, butyric acid, carbon dioxide, ammonia and hydrogen.
Butyric acid and ammonia impart obnoxious (rancid) odours to food.34 The incomplete metabolism of amino acids may result in putrescence, while the liberation of hydrogen sulphide from amino acids may result in a sulphide type of spoilage.35
On the more beneficial side, amino acids are the key precursors for ripened cheese flavour. They are metabolised by the action of amino acid-converting enzymes to aldehydes, alcohols, ketones, amines, acids, esters and sulfur-containing compounds, which all contribute to cheese flavour. While this action is desirable in some cheeses, it may be regarded as food spoilage in other products.36-41 The off-flavours resulting from the above-mentioned compounds may range from fruity, malty, ripened-cheese to rancid, sweaty and putrid.42
While the catabolism of amino acids is not a direct indication of hydrolytic breakdown of peptides or proteins, it is known that the flavobacteria are often psychrotrophic and proteolytic.7-9 The majority of psychrotrophic bacteria have the ability to form hydrolytic, thermostable enzymes that break down the major constituents of milk, such as the protein.2 The action of such enzymes destabilises the casein in milk through hydrolysis, and the result is the formation of a gel structure or coagulation of sterilised milk during storage.2 The problem of the destabilisation of milk proteins leading to the precipitation or 'flocculation' of milk proteins during heat processing of milk has been identified as an important problem by the National Milkproducer's Research Workgroup of MilkSA.43 Proteolytic enzymes of psychrotrophic bacteria have been cited as a possible contributing factor to this problem in certain milk producing regions of South Africa.
Utilisation of carboxylic acids
While acetic acid was metabolised by all the Chryseobacterium species (except C. aquaticum) and E. brevis in this study (Table 4), C. bovis and E. brevis were the only isolates capable of utilising o-keto valeric acid. The metabolism of carboxylic acids by micro-organisms results in the coagulation of milk accompanied by sour and bitter tastes in milk.44
Formation of flavour compounds from branched chain o-keto acids in fermented foods such as cheese is believed to be mainly an enzymatic process, because fermented product flavours are derived largely from amino acids.45 Amino acids are mostly enzymatically transaminated in the bacterial cell to the corresponding o-keto acid. These o-keto acids are converted to various metabolites, such as the aldehydes and sulfur compounds which may result in off-flavours.46
Utilisation of polymers
α-Cyclodextrin and dextrin were the polymers most widely utilised by C. aquaticum, C. indoltheticum, C. bovis and E. brevis (Table 4). The polymers include polysaccharides. During food spoilage, complex di-, tri- or polysaccharides are usually hydrolysed to simple sugars before utilisation by microorganisms. Some of the metabolic products resulting from the oxidation of these polysaccharides include organic acids, alcohols and CO2, which may produce off-odours, sourness and bitter defects.34,47
Some strains of C. bovis, C. shigense, C. indologenes and E. brevis were able to utilise Tween 40 and Tween 80 (Table 4). Tween 40 and 80 hydrolysis serves as an indicator of lipase and other esterase activity48 that can lead to spoilage of long-life dairy products during storage. Lipolysis by flavobacteria/chryseobacteria is known to break down fatty acids, resulting in rancid, butyric, bitter, soapy and astringent off-flavours and off-odours in some dairy products.49,50
When comparing the utilisation of the 22 substrates by the isolates tested in this study, C. bovis, C. shigense and E. brevis had the ability to utilise 16 of the 22 substrates, which could indicate that these species have the potential to cause more pronounced types of spoilage than the other species tested. The frequency of utilisation of the 22 carbon sources by the remaining isolates were as follows: C. aquaticum (10/22), C. indologenes (9/22), C. indoltheticum (9/22), C. haifense (5/22), C. ureilyticum (5/22), Chryseobacterium sp. Vv5a (4/22).
Conclusions
Flavobacteria previously isolated from dairy sources were shown in this study to belong to a range of Chryseobacterium species and to Empedobacter brevis by employing 16S rRNA sequencing and a variety of phenotypic tests. Although the BIOLOG system could not be used as an identification tool because of the limited database, it proved to be an excellent differentiation tool on the basis of the substrates utilised by the isolates. The BIOLOG system was also valuable in determining potential food spoilage characteristics of the isolates. The carbon sources most frequently utilised in the BIOLOG microplate system by the isolates were glucose, acetic acid, dextrin as well as amino acids such as glycyl-L-aspartic acid, glycyl-glutamic acid and L-glutamic acid. These may lead to spoilage characteristics such as off-odour production (e.g. fruity, malty, ripened-cheese, rancid, sweaty, putrid, sulphide-type), coagulation of milk, sourness and bitterness. Many of these spoilage characteristics result from the action of hydrolytic enzymes produced by the organisms. Proteolytic enzymes of psychrotrophic bacteria have been cited as a possible contributing factor to the important problem of milk 'flocculation' recently being encountered in certain milk producing regions of South Africa.
The flavobacterial species are believed to have the potential to cause spoilage defects in dairy products because they were able to utilise a wide range of compounds in the BIOLOG system. In this regard C. bovis, C. shigense and E. brevis were the most active and could utilise 16 of the 22 carbon sources even though varied reactions were shown where multiple strains of the organisms were tested. This was followed by C. aquaticum (10/22), C. indoltheticum and C. indologenes (9/22). Species that were least active included C. haifense and Chryseobacterium sp. Vv5a (4/22). These findings illustrate that enzymatically catalysed defects in dairy products should not be attributed solely to acknowledged psychrotrophic bacteria such as the pseudomonads, but that flavobacterial species may also be actively involved.
Authors' contributions
C.H. was the project leader and responsible for experimental and project design; L.I.T. performed the experiments; P.J.J. made conceptual contributions; G.C. contributed to the interpretation and discussion of the biological results; L.I.T., P.J., G.C. and C.J.H. wrote the manuscript.
Acknowledgements
This work was supported by funding from the South African National Research Foundation.
References
1. Lafarge V, Ogier J-C, Girard V, Maladen V, Leveau J-Y, Gruss A, Delacroix-Buchet A. Raw cow milk bacterial population shifts attributable to refrigeration. Appl Environ Microbiol. 2004;70:5644-5650. http://dx.doi.org/10.1128/AEM.70.9.5644-5650.2004 [ Links ]
2. Samaržija D, Zamberlin Š, Pogačič T. Psychrotrophic bacteria and milk and dairy products quality. Mljekarstvo. 2012;62:77-95. [ Links ]
3. IDF Bulletin. Psychrotrophs in milk and milk products: E-doc 68. Brussels: International Dairy Federation; 1976. [ Links ]
4. Chen L, Daniel RM, Coolbear T. Detection and impact of protease and lipase activities in milk and milk powder (review). Int Dairy J. 2003; 13:255-275. http://dx.doi.org/10.1016/S0958-6946(02)00171-1 [ Links ]
5. Stepaniak, L. Psychrotrophic bacteria, bacteria other than Pseudomonas spp. In: Roginski H, Fuguay J, Fox PF, editors. Encyclopedia of dairy sciences, Vol. 4. London: Academic Press; 2002. p. 2345-2351. http://dx.doi.org/10.1016/B0-12-227235-8/00407-7 [ Links ]
6. Cousin MA. Presence and activity of psychrotrophic microorganisms in milk and dairy products: A review. J Food Protect. 1982;45:172-207. [ Links ]
7. Gilmour A, Rowe MT. Microorganisms associated with milk. In: Robinson RK, editor. Dairy microbiology. London: Applied Science; 1981. p. 35-75. [ Links ]
8. Jooste PJ, Britz TJ. The significance of flavobacteria as proteolytic psychrotrophs in milk. Milchwissenschaft. 1986;41:618-621. [ Links ]
9. Venter H, Osthoff G, Litthauer D. Purification and characterization of a metalloprotease from Chryseobacterium indologenesIx9a and determination of the amino acid specificity with electrospray mass spectrometry. Prot Expr Purif. 1999;15:282-295. http://dx.doi.org/10.1006/prep.1998.1020 [ Links ]
10. Meer RR, Baker J, Bodyfelt FW, Griffiths MW. Psychrotrophic Bacillus spp. in fluid milk products: A review. J Food Protect. 1991;54:969-979. [ Links ]
11. Jooste PJ, Hugo CJ. The taxonomy, ecology and cultivation of bacterial genera belonging to the family Flavobacteriaceae (a review). Int J Food Microbiol. 1999;53:81-94. http://dx.doi.org/10.1016/S0168-1605(99)00162-2 [ Links ]
12. Mielmann A, Hugo CJ, Jooste PJ. Food spoilage characteristics of Chryseobacterium species. Role and significance in food. Saarbrücken: VDM Verlag Dr. Müller; 2009. [ Links ]
13. Bekker A. Growth and spoilage characteristics of Chryseobacterium species in milk [MSc dissertation]. [ Links ] Bloemfontein: University of the Free State; 2011.
14. Jooste PJ. The taxonomy and significance of Flavobacterium-Cytophaga strains from dairy sources [PhD thesis]. [ Links ] Bloemfontein: University of the Orange Free State; 1985.
15. Reichenbach H. Flavobacteriaceae fam. nov.: Validation of the publication of new names and new combinations previously effectively published outside the IJSB, List no. 41. Int J Syst Evol Microbiol. 1992;42:327-329. http://dx.doi.org/10.1099/00207713-42-2-327 [ Links ]
16. Parte AC. List of prokaryotic names with standing in nomenclature [document on the Internet]. [ Links ] c2015 [cited 2015 Aug 07]. Available from http://www.bacterio.net
17. Hugo CJ, Jooste PJ. Culture media for food associated genera in the family Flavobacteriaceae. In: Corry JEL, Curtis GDW, Baird RM, editors. Handbook of culture media for food and water microbiology. Cambridge: RSC Publishing; 2012. p. 508-556. [ Links ]
18. Vandamme P Bernardet J-F, Segers P Kersters K, Holmes B. New perspectives in the classification of the flavobacteria: description of Chryseobacterium gen. nov., Bergeyella gen. nov., and Empedobacter nom. rev. Int J Syst Bacteriol. 1994;44:827-831. http://dx.doi.org/10.1099/00207713-44-4-827 [ Links ]
19. Bernardet J-F, Hugo CJ, Bruun B. Genus X. Chryseobacterium Vandamme, Bernardet, Segers, Kersters and Holmes 1994, 829VP In: Krieg NR, Staley JT, Brown DR, Hedlund BP, Paster BJ, Ward NL, et al., editors. Bergey's manual of systematic bacteriology. 2nd ed. New York: Springer; 2011. p. 180-196. [ Links ]
20. Bernardet J-F, Nakagawa Y, Holmes B. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family. Int J Syst Evol Microbiol. 2002;52:1049-1070. http://dx.doi.org/10.1099/00207713-52-3-1049 [ Links ]
21. Labuschagne M, Albertyn J. Cloning of an epoxide hydrolase-encoding gene from Rhodotorula mucilaginosa and functional expression in Yarrowia lipolytica. Yeast. 2007;24:69-78. http://dx.doi.org/10.1002/yea.1437 [ Links ]
22. Lane DJ. 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M, editors. Nucleic acid techniques in bacterial systematics. Chichester: Wiley; 1991. p. 115-175. [ Links ]
23. National Center for Biotechnology Information (NCBI). GenBank Overview. Rockville, MD: NCBI; 2011. Available from: http://www.ncbi.nlm.nih.gov/genbank/. [ Links ]
24. Kumar S, Nei M, Dudley J, Tamura K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief Bioinform. 2008;9:299-306. http://dx.doi.org/10.1093/bib/bbn017 [ Links ]
25. Hugo CJ, Segers P Hoste B, Vancanneyt M, Kersters K. Chryseobacterium joostei sp. nov., isolated from the dairy environment. Int J Syst Evol Microbiol. 2003;53:771-777. http://dx.doi.org/10.1099/ijs.0.02232-0 [ Links ]
26. Kim KK, Lee KC, Oh H-M, Lee JS. Chryseobacterium aquaticum sp. nov., isolated from a water reservoir. Int J Syst Evol Microbiol. 2008;58:533-537. http://dx.doi.org/10.1099/ijs.0.65491-0 [ Links ]
27. Hantsis-Zacharov E, Halpern M. Chryseobacterium haifense sp. nov., a psychrotolerant bacterium isolated from raw milk. Int J Syst Evol Microbiol. 2007;57:2344-2348. http://dx.doi.org/10.1099/ijs.0.65115-0 [ Links ]
28. Hantsis-Zacharov E, Senderovich Y Halpern M. Chryseobacterium bovis sp. nov., isolated from raw cow's milk. Int J Syst Evol Microbiol. 2008;58:1024-1028. http://dx.doi.org/10.1099/ijs.0.65500-0 [ Links ]
29. Campbell LL, Williams OB. A study of chitin-decomposing micro-organisms of marine origin. J Gen Microbiol. 1951;5:894-905. http://dx.doi.org/10.1099/00221287-5-5-894 [ Links ]
30. Shimomura K, Kaji S, Hiraishi A. Chryseobacterium shigense sp. nov., a yellow-pigmented, aerobic bacterium isolated from a lactic acid beverage. Int J Syst Evol Microbiol. 2005;55:1903-1906. http://dx.doi.org/10.1099/ijs.0.63690-0 [ Links ]
31. Herzog P Winkler I, Wolking D, Kämpfer P Lipski A. Chryseobacterium ureilyticum sp. nov., Chryseobacterium gambrini sp. nov., Chryseobacterium pallidum sp. nov. and Chryseobacterium molle sp. nov., isolated from beer-bottling plants. Int J Syst Evol Microbiol. 2008;58:26-33. http://dx.doi.org/10.1099/ijs.0.65362-0 [ Links ]
32. Dainty RH. Chemical/biochemical detection of spoilage. Int J Food Microbiol. 1996;33:19-33. http://dx.doi.org/10.1016/0168-1605(96)01137-3 [ Links ]
33. Ellis DI, Goodacre R. Quantitative detection and identification methods for microbial spoilage. In: De Blackburn C, editor. Food spoilage microorganisms. Cambridge: Woodhead Publishing; 2006. p. 3-27. http://dx.doi.org/10.1533/9781845691417.1.3 [ Links ]
34. Banwart GJ. Basic food microbiology. 2nd ed. New York: Van Nostrand Reinhold; 1989. http://dx.doi.org/10.1007/978-1-4684-6453-5_3 [ Links ]
35. Ayres JC, Mundt JO, Sandine WE. Microbiology of foods. San Francisco: WH Freeman and Company; 1980. [ Links ]
36. Aston JW, Creamer LK. Contribution of the components of the water-soluble fraction to the flavour of cheddar cheese. N Z J Dairy Sci Technol. 1986;21:229-248. [ Links ]
37. Crow VL. Metabolism of aspartate by Propionibacterium freudenreichii subsp. shermanii: Effect on lactate fermentation. Appl Environ Microbiol. 1986;52:359-365. [ Links ]
38. Crow VL. Properties of alanine dehydrogenase and aspartase from Propionibacterium freudenreichii subsp. shermanii. Appl Environ Microbiol. 1987;53:1885-1892. [ Links ]
39. Harper WJ, Wang JY. Amino acid catabolism in cheddar cheese slurries. I. Formation of selected products from alanine. Milchwissenschaft. 1980a;35:531-535. [ Links ]
40. Harper WJ, Wang JY. Amino acid catabolism in cheddar cheese slurries. II. Evaluation of transamination. Milchwissenschaft. 1980b;35:598-599. [ Links ]
41. Harper WJ, Wang JY. Amino acid catabolism in cheddar cheese slurries. III. Selected products from glutamic acid. Milchwissenschaft. 1981;36:70-72. [ Links ]
42. Engels WJM, Dekker R, De Jong C, Neeter R, Visser S. A comparative study of volatile compounds in the water-soluble fraction of various types of ripened cheese. Int Dairy J. 1997;7:255-263. http://dx.doi.org/10.1016/S0958-6946(97)00003-4 [ Links ]
43. Milk South Africa. Heat stability of reconstituted, protein-standardized skim milk powders [column on the Internet]. [ Links ] c2011 [cited 2015 Aug 07]. Available from: http://www.milksa.co.za/research/research-column/heat-stability-reconstituted-protein-standardized-skim-milk-powders
44. Urbach G. The flavour of milk and dairy products: II. Cheese: Contribution of volatile compounds. Int J Dairy Technol. 1997;50:79-89. http://dx.doi.org/10.1111/j.1471-0307.1997.tb01743.x [ Links ]
45. Smit BA, Engels WJM, Alewijn M, Lommerse GTCA, Kippersluisjs EAH, Wouters JTM, et al. Chemical conversion of a-keto acids in relation to flavor formation in fermented foods. J Agric Food Chem. 2004;52:1263-1268. http://dx.doi.org/10.1021/jf035147z [ Links ]
46. Gao S, Oh DH, Broadbent JR, Johnson ME, Weimer BC, Steele JL. Aromatic amino acid catabolism by lactococci. Le Lait. 1997;77:371-381. http://dx.doi.org/10.1051/lait:1997325 [ Links ]
47. Coultate TP. Food: The Chemistry of its components. London: Royal Society of Chemistry; 1984. [ Links ]
48. Harrigan WF, McCance ME. Laboratory methods in food and dairy microbiology. London: Academic Press; 1976. [ Links ]
49. Deeth HC, Fitz-Gerald CH. Lipolytic enzymes and hydrolytic rancidity in milk and milk products. In: Fox PF, editor. Advanced dairy chemistry, Vol. 2: Lipids. London: Chapman & Hall; 1995. p. 247-308. [ Links ]
50. Liao CH. Pseudomonas and related bacteria. In: De Blackburn C, editor. Food spoilage microorganisms. Cambridge: Woodhead Publishing; 2006. p. 507-540. http://dx.doi.org/10.1533/9781845691417.5.507 [ Links ]
51. Kim KK, Kim MK, Lim JH, Park HY, Lee ST. Transfer of Chryseobacterium meningosepticum and Chryseobacterium miricola to Elizabethkingia gen. nov. as Elizabethkingia meningoseptica comb. nov. and Elizabethkingia miricola comb. nov. Int J Syst Evol Microbiol. 2005;55:1287-1293. http://dx.doi.org/10.1099/ijs.0.63541-0 [ Links ]
 Correspondence:
Correspondence:
Celia Hugo
Department of Microbial, Biochemical and Food Biotechnology, University of the Free State, PO Box 339, Bloemfontein, 9301, South Africa
hugocj@ufs.ac.za
Received: 18 June 2015
Revised: 07 Aug. 2015
Accepted: 07 Aug. 2015


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}