










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.112 no.1-2 Pretoria Jan./Fev. 2016
http://dx.doi.org/10.17159/sajs.2016/20150072
REVIEW ARTICLE
Neutrophil extracellular traps and their role in health and disease
Jan G. NelI; Annette J. TheronII; Roger PoolI; Chrisna DurandtIII; Gregory R. TintingerIV; Ronald AndersonIII
IDepartment of Haematology, University of Pretoria, Pretoria, South Africa
IIDepartment of Immunology, University of Pretoria, Pretoria, South Africa
IIIInstitute for Cellular and Molecular Medicine, SAMRC Extramural Unit for Stem Cell Research and Therapy, Department of Immunology, University of Pretoria, Pretoria, South Africa
IVDepartment of Internal Medicine, University of Pretoria and Steve Biko Academic Hospital, Pretoria, South Africa
ABSTRACT
The human innate immune system is indispensable for protection against potentially invasive microbial and viral pathogens, either neutralising them or containing their spread until effective mobilisation of the slower, adaptive (specific), immune response. Until fairly recently, it was believed that the human innate immune system possessed minimal discriminatory activity in the setting of a rather limited range of microbicidal or virucidal mechanisms. However, recent discoveries have revealed that the innate immune system possesses an array of novel pathogen recognition mechanisms, as well as a resourceful and effective alternative mechanism of phagocyte (predominantly neutrophil)-mediated, anti-infective activity known as NETosis. The process of NETosis involves an unusual type of programmed, purposeful cell death, resulting in the extracellular release of a web of chromatin heavily impregnated with antimicrobial proteins. These structures, known as neutrophil extracellular traps (NETs), immobilise and contribute to the eradication of microbial pathogens, ensuring that the anti-infective potential of neutrophils is sustained beyond the lifespan of these cells. The current review is focused on the mechanisms of NETosis and the role of this process in host defence. Other topics reviewed include the potential threats to human health posed by poorly controlled, excessive formation of NETs, specifically in relation to development of autoimmune and cardiovascular diseases, as well as exacerbation of acute and chronic inflammatory disorders of the airways.
Keywords: antimicrobial activity; autoimmune disease; innate immunity; phagocytes; reactive oxygen species
Introduction
Until fairly recently, the protective activities of the human innate immune system, which are highly conserved throughout evolution, were thought to be achieved by a limited range of mechanisms with minimal discriminatory potential. Predominant amongst these mechanisms are the physical barriers presented by the skin and mucous membranes, engulfment and destruction of microbial and viral pathogens by resident and migratory phagocytes, and the non-specific antimicrobial activity of various blood and tissue proteins. These mechanisms either prevent infection or, in the case of a breach by a pathogen, contain the infection until adaptive (specific) host defences are effectively mobilised. Over the last decade, however, a number of significant discoveries have revealed that the human innate immune system not only possesses a level of discrimination previously considered improbable, but also includes additional, resourceful mechanisms of phagocyte-mediated antimicrobial and antiviral activity.
With respect to pathogen detection, cells of the innate immune system (phagocytes, mast cells, basophils and dendritic cells), as well as epithelial cells, have been found to possess various types of pathogen recognition receptors which recognise conserved molecular structures broadly expressed on or in microbial and viral pathogens. These receptors include the Toll-like receptors, the nucleotide oligomerisation domain-like receptors, and the abundant cytosolic microbial and viral nucleic acid sensors, activation of which initiates a potentially protective inflammatory response. These receptors have been the subject of several recent reviews.1,2
In 2004, Brinkmann et al.3 described an unusual mechanism by which human blood neutrophils immobilise pathogens extracellularly, exposing them to a highly concentrated array of anti-infective proteins. Neutrophils (also known as granulocytes or polymorphonuclear leucocytes) are the predominant small circulating phagocytes. These cells have an estimated lifespan of 5.4 days in the circulation,4 which is longer when they are exposed to anti-apoptotic cytokines. These cells exit the circulation via transendothelial migration and chemotaxis to sites of microbial and viral infection where they phagocytose and destroy pathogens via intracellular exposure to microbicidal and virucidal reactive oxygen species (ROS), proteases and proteins.
The studies of Brinkmann et al.3 enhanced the body of knowledge on neutrophil function through the discovery that these cells also respond to infectious challenges via the formation of neutrophil extracellular traps (NETs). NETs are web-like structures composed of decondensed chromatin heavily impregnated with different antimicrobial granular proteins which capture, neutralise and kill a variety of pathogens. NETs are produced predominantly by neutrophils, but also by other cell types of the innate immune system such as monocytes and macrophages, eosinophils, basophils and mast cells, in which the process is termed ETosis. Phylogenetic studies have revealed that ETosis is a highly conserved 'ancient defence weapon, predating the evolution of the coelom' operative in haemocytes, the phagocytic cells of invertebrates.5
NETs form large extracellular barriers to bacterial dissemination, and provide a mechanism for localised concentration of effector molecules. Importantly, NET formation has been demonstrated in both the clinical and experimental infection settings using immunohistochemistry and spinning disc vital microscopy.6 Several strategies to measure NET formation in vitro have also been described, including immunofluorescence and electron microscopic procedures, as well as spectrofluorimetric and other methods which detect extracellular DNA and associated granule proteins.3 Human neutrophils undergoing NETosis in vitro are shown in Figure 1.
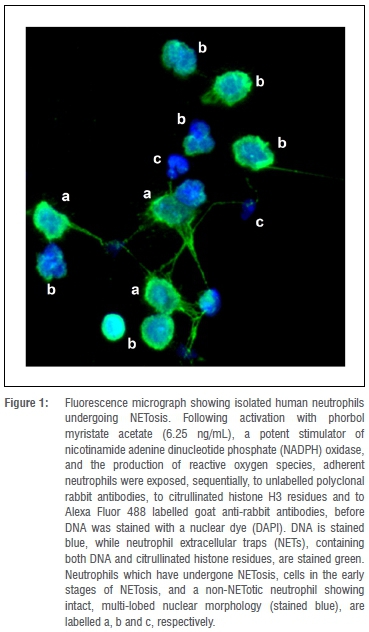
NETs are the topic of this review, which is focused on mechanisms of NETosis and the role of this process in host defence, as well as on the potentially harmful consequences of excessive NETosis for the host and possible pharmacological control strategies.
Mechanisms of NET formation
Although the exact molecular and biochemical mechanisms involved in the formation and release of NETs are incompletely understood, three different types of NETotic pathway have been described. The best characterised of these pathways is slow and leads to lytic cell death over the course of 2-3 h. The second pathway has been described as a rapid mechanism (vital), independent of cell lysis, which requires the rapid (within minutes) vesicular release of neutrophil nuclear contents.6,7 Unlike the first two mechanisms of NETosis which involve release of nuclear DNA, the third mechanism, which is also non-lytic, involves the release of mitochondrial DNA.8
Lytic NETosis, also known as suicidal NETosis, is an active cell death related process distinct from either necrosis or apoptosis.9 This process requires chromatin decondensation, nuclear envelope disintegration and a mixing of nucleic acids and granule proteins within an intracellular vacuole. Subsequent release of vacuole contents into the extracellular domain is preceded by plasma membrane perforation or lysis. Various stages in the process constituting lytic NETosis have been identified. The interaction of pathogens and their products with their counter-receptors on or in neutrophils includes, in addition to the pathogen recognition receptors mentioned above, the opsonin receptors FcR (receptor for pathogen-bound immunoglobulin G) and CR3 (receptors for pathogen-bound complement components C3b and C3bi), which promote adherence of pathogens to neutrophils. This interaction, in turn, leads to a series of pathogen-activated intracellular signalling events, most importantly: (1) activation of the receptor-linked signalling complex protein kinase C(PKC)-raf/-MEK-ERK which mediates activation of the neutrophil membrane-associated, electron-transporting, ROS-generating
system, NADPH oxidase10; (2) ROS-mediated activation of intracellular signalling pathways converging on the cytosolic transcription factor, nuclear factor kappa B (NFkB), which, following nuclear translocation, has been proposed to promote NETosis by initiating transcription of the peptidylarginine deiminase 4 (PAD4) gene, as well as by blocking apoptosis, an alternative pathway of programmed cell death11; and (3) receptor-mediated increases in cytosolic Ca2+ via activation of phospholipase C, which, in turn, leads to Ca2+-dependent activation of PAD4. ROS also promote collapse of both the cytoplasmic granule and nuclear membranes, enabling access of cytoplasmic PAD4 and granule proteins to chromatin. PAD4 mediates histone hypercitrullination (conversion of protein arginine residues to citrulline), a key event in chromatin decondensation12,13, which is facilitated by the limited proteolysis of nuclear histones mediated by granule enzymes - neutrophil elastase and myeloperoxidase (MPO) - operating in unison3,14. While ROS appear to promote the rupture of both the cytosolic granule and nuclear membranes, a recent study has also implicated the involvement of the granule antimicrobial polypeptide LL-37 (which consists of 2 N-terminal leucines and a total of 37 amino acids).15 LL-37 is a cationic amphiphilic polypeptide of the cathelicidin family which binds to anionic membrane phospholipids promoting membrane disruption. The nuclear membrane appears particularly vulnerable to the disruptive actions of LL-37. In addition, the cationic properties of LL-37 also promote binding of the polypeptide to neutrophil DNA, increasing resistance to degradation of NETs by microbial nucleases.16
The proposed mechanism of lytic NET formation based on current knowledge is shown in Figure 2.
The concept that alternative NETosis pathways exist, in addition to the lytic-cell death pathway, has been advanced by several groups.6,8,17,18 One such pathway is vital NETosis. Vital NETosis is a rapid process whereby cell viability and function are retained in the context of controlled, incremental discharge of nuclear material, following exposure of neutrophils to various microorganisms and their products, and appears to be a generalised response against various classes of microbial pathogens.18,19 In this setting, neutrophils are stimulated to 'release NETs via nuclear envelope bleb formation and vesicular exportation, preserving the integrity of the plasma membrane'.6,20 Importantly, and also in contradistinction to lytic NETosis, the requirement for involvement of NADPH oxidase in vital NETosis is variable, with increases in cytosolic calcium seemingly adequate in the case of some stimuli such as bacterial pore-forming toxins6 and calcium ionophores7, or alternatively via ROS-independent activation of NFkB21. Unlike lytic NETosis, only 20-25% of the neutrophil population undergoes NETosis on exposure to microbial pathogens or their products in vitro.20 This observation is not only consistent with the existence of a sub-population of neutrophils highly specialised for the performance of vital NETosis, but also suggests that vital, as opposed to lytic, NETosis is the more physiologically relevant of the two processes.20
The formation of mitochondrial-derived NETs is also a type of vital NETosis with variable dependence on activation of NADPH oxidase according to the nature of the cell activator. Although the existence of mitochondrial DNA-containing NETs has been demonstrated in vitro, less is known about its role in host defence which is likely to be limited by the absence of histones.22,23 Interestingly, mitochondrial DNA has been found to induce NET formation, consistent with a role in the amplification of NETosis.22
NET constituents
The DNA scaffolding of NETs is provided by decondensed nuclear chromatin. A variety of NET-associated proteins, predominantly highly negatively charged histones, is arranged on this scaffolding. Via their strong positive charge, histones facilitate the adhesion to, as well as the sequestration of, microbial and viral pathogens24 and also confer bactericidal activity, as does DNA.25 The remaining NET-associated proteins comprise granule, cytoplasmic and cytoskeletal proteins, as well as metabolic enzymes.24 The concept of a core NET-associated proteome that remains constant regardless of the specific agent responsible for NET induction, has been proposed by Rahman and Gadjeva26. The core NET-associated proteome is made up of a combination of 19 constituent proteins, arranged in association with a variety of decorative proteins. The fact that the protein structure of NETs exhibits a constant element to its composition supports the hypothesis that NET formation is indeed an innate immune system response which is non-specific.26 The major antimicrobial components of NETs are summarised in Table 1.
Beneficial effects of NETosis
Role in host defence
NETs have been shown to degrade microbial and viral virulence factors and to restrict pathogens by forming a physical barrier that prevents dissemination. Examples of bacterial, fungal and protozoal parasitic pathogens which are ensnared in NETs are shown in Table 2. While some bacterial pathogens such as Pseudomonas aeruginosa27, Borrelia burgdorferi28 and Burkholderi pseudomallei29 are killed following entrapment in NETs, others appear less vulnerable, probably as a result of the production of anti-adhesive surface structures such as polysaccharide capsules and/or NET-degrading nucleases, as indicated in Table 2.24,27-47
Neutrophils appear to be particularly adept at sensing microbial size, with large microorganisms the most effective inducers of NETosis.31 For example, in the case of Candida albicans, the yeast responsible for most fungal infections in humans, NETs are effectively induced by this pathogen in both the yeast and hyphal forms.48 Protozoa such as Toxoplasma gondii, Plasmodium falciparum and Leishmania spp. have also been shown to possess the requisite signals to trigger NET formation, which, in some cases, may lead to the death of entrapped parasites, as well as interference with the invasion of host cells. Some microbial pathogens, such as the opportunistic Gram-negative bacterium Acinetobacter baumannii and the fungus Cryptococcus neoformans, do not appear to activate NETosis, which in the case of the latter has been attributed predominantly to the ability of the polysaccharide capsule to prevent the requisite signalling mechanisms.49,50
Based on observations of pathogens such as Staphylococcus aureus and Streptococcus pneumoniae, which withstand NETs, it has been proposed that the primary function of NETs is to immobilise, weaken and expose infective agents to other antimicrobial cellular and humoral components of the innate immune system, including tissue macrophages and the proteins of the complement system respectively.51-53 An example of this type of cooperation involves the interaction of neutrophils and macrophages in the eradication of the bacterial pathogen Mycobacterium tuberculosis. This pathogen has also been reported to induce NETs in which it is trapped extracellularly, but remains viable. Entrapped bacilli are then engulfed by alveolar macrophages, the primary cell type involved in the eradication of M. tuberculosis.30
With respect to the role of NETs in antiviral host defence, neutrophils have been shown to detect HIV-1 via interaction with pathogen recognition receptors which recognise viral RNA. This detection in turn triggers NET formation, leading to NET-mediated inactivation of HIV-1, resulting from exposure to MPO-derived oxidants and a-defensins.54 This response may, however, be attenuated via the release of the anti-inflammatory cytokine, interleukin-10, from bystander HIV-infected dendritic cells, which, in turn, inhibits NET formation.54
Although the exact role of NETs in the host response to infection remains to be convincingly elucidated, the increased susceptibility of patients with chronic granulomatous disease (CGD) to infection is noteworthy in this context. This condition is an inherited primary immunodeficiency disorder caused by a complete absence of NADPH oxidase and failure of phagocytes to generate ROS, and consequently NETs. The severe impairment of neutrophil protective activity as a result of the combined absence of production of antimicrobial ROS and NETs causes the patients to suffer from severe and often life-threatening infections.9 However, NET production by neutrophils in chronic granulomatous disease is responsive to activation by ROS-independent pro-NETotic mechanisms.21 In addition, transient and acquired abnormalities of NET formation have also been demonstrated in human neonates and the elderly, respectively - a previously unrecognised deficit in extracellular bacterial killing which may underpin age-associated vulnerability to microbial and viral infection.55,56
In addition, the increased susceptibility of humans with stable or transient severe neutropenia for development of disseminated fungal infections has been attributed to attenuation of 'trapping' by NETs.20
Evasion of NETs by pathogens
Several strategies have been described that enable pathogens to evade NET-mediated immobilisation and/or killing by NET-associated proteins, many of which are listed in Table 2. These strategies include the production of nuclease enzymes that degrade the DNA backbone of the NET structures by various types of microbial pathogens, including, but not limited to, pathogens of the Streptococcus and Staphylococcus genera. In addition, the acquisition by microorganisms and viruses of molecular patterns that interfere with pathogen recognition by pathogen recognition receptors has also been reported to attenuate NET formation.34-36 The acquisition of a cell capsule, for example, is of particular benefit in evading NET-mediated trapping of organisms, by altering the surface charge of bacteria to neutral and thereby negating the electrostatic attraction posed by positively charged NET fibres and histone residues.36
The involvement of NETs in the pathophysiology of disease
Despite beneficial effects in host defence, NETosis may occur at the expense of injury to the host.57 Inappropriate and/or excessive NET formation has been documented in the following autoimmune, cardiovascular and pulmonary diseases.
The role of NETs in autoimmune diseases
Systemic lupus erythematosus
Systemic lupus erythematosus (SLE) is a complex and heterogeneous disease, with patients displaying a variety of symptoms of which glomerulonephritis is particularly serious.57,58 This condition has a definite female preponderance and a prevalence which seemingly varies according to race. The hallmark of SLE is the overproduction of autoantibodies against a range of nuclear antigens including, not only DNA and histones, but also neutrophil granule proteins.57 These autoantibodies are believed to contribute significantly to disease pathogenesis. In this context, several studies have reported that the ability to degrade NETs was reduced in a subset of patients with a severe form of SLE which was associated with both glomerulonephritis and the presence of circulating autoantibodies reactive with various constituents of NETs.59,60 Mechanistically, disassembly of NETs in the physiological setting is mediated by the enzyme serum endonuclease DNase1, interference with which is likely to favour persistence and exaggerated immunogenicity of NETs.60 In this setting, binding of anti-NET antibodies to NETs has been reported to prevent access of DNase I to NETs. The consequence is impairment of DNase1 function, resulting in failure to dismantle NETs, correlating with renal involvement in SLE.60 As alluded to earlier, it appears that NETosis in SLE also involves a subset of neutrophils known as low density granulocyes.61
Rheumatoid arthritis
Rheumatoid arthritis (RA) is a chronic, systemic autoinflammatory disease which primarily affects the synovial joints. It occurs at high frequency (1-3%) in the general population with a female preponderance and, like SLE, is associated with high morbidity and mortality. The majority of RA patients present with high levels of circulating antibodies to citrullinated proteins known as anti-citrullinated peptide antibodies (ACPAs) which are serodiagnostic for RA. Although a clear mechanistic relationship between dysregulation of NETosis and production of ACPAs remains to be established, it is noteworthy that neutrophils from patients with RA exhibit exaggerated NETosis in the circulation, the skin and rheumatoid joint when compared with neutrophils from healthy controls and patients with osteoarthritis.62,63 Other supporting evidence includes:
(1) the finding of a significant positive correlation between NET formation and serum levels of anti-citrullinated peptide antibodies, as well as with other circulating biomarkers of inflammation and neutrophil activation; (2) a report that citrullinated histone 4, a component of NETs, is reactive with ACPAs; and (3) the finding that NETs act as strong stimulants of fibroblast-like synoviocytes (cells that invade cartilage in RA).62 Taken together, these findings appear to implicate dysregulation of NETosis in the pathogenesis of RA which, in turn, may lead to the identification of novel targets for the treatment of this and other diseases.62
Small vessel vasculitis
Small vessel vasculitis is a chronic autoinflammatory condition in which small blood vessels show necrotic inflammation. The condition is associated with the presence of anti-neutrophil cytoplasmic autoantibodies (ANCAs). The main targets for ANCAs are the granule enzymes MPO and PR3 (proteinase 3).64 Kessenbrock et al.65 observed that the binding of ANCAs to neutrophils resulted in activation of NETosis. They also demonstrated typical components of NETs present in kidney biopsies of patients with small vessel vasculitis. The NETs were decorated with the autoantigens MPO and PR3. Deposition of NETs in inflamed kidneys suggests that NET formation plays a pathogenic role in autoimmune small vessel vasculitis by presenting autoantigens to the immune system with resultant vascular damage.65,66
The role of NETs in deep vein thrombosis
Deep vein thrombosis (DVT) is the formation of a blood clot (or thrombus) in a deep vein, predominantly in the legs. DVT can be triggered by disturbances in venous blood flow, activation or dysfunction of the vascular endothelium, and hypercoagulability. NETs provide a new link between innate immunity and hypercoagulability,67 stimulating the coagulation process by activating platelets, the coagulation cascade and the vascular endothelium.68 NETs provide a scaffold for platelet and red blood cell adhesion and also concentrate effector proteins involved in thrombosis.68 It is speculated that NET-associated enzymes may enhance coagulation indirectly through proteolytic degradation of tissue factor pathway inhibitor, the major trigger protein in the onset of blood clotting.69 Histones have also been shown to increase thrombin generation, causing platelet activation and coagulation.70 In the experimental setting, administration of DNase1 to mice was found to promote disassembly of NETs with resultant suppression of DVT enlargement,71,72 underscoring the importance of NETS in the pathogenesis of DVT.67 The presence of NETs, according to the detection of citrullinated histone H3 positive cells, has also been described in human venous thrombi.73
NETs in lung disease
NETs may also contribute to the pathogenesis and severity of several inflammatory lung conditions, including cystic fibrosis74, acute lung injury and acute respiratory distress syndrome75, severe asthma76 and chronic obstructive pulmonary disease77.
Acute lung injury and acute respiratory distress syndrome
Acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) represent a spectrum of lung diseases resulting from direct and indirect insults to the lung. These insults may be a result of infectious or sterile causes.78 The disease process is characterised by a disruption of the endothelial-epithelial barriers, alveolar damage, pulmonary oedema and various degrees of respiratory failure.78 ALI/ARDS is characterised by an influx of neutrophils into the pulmonary capillaries, with retention of hyperreactive neutrophils in the damaged vasculature.79 NETs are able to contribute directly to the pathology of ALI/ARDS by inducing lung epithelial cell death.66
NETs in cystic fibrosis
Cystic fibrosis (CF) is a lifelong inherited condition primarily affecting the lungs and digestive tract, with prevalence varying according to race, and seemingly more common in those of North European descent. CF patients develop chronic lung infections associated with airway obstruction mediated by viscous and insoluble mucus secretions.80 In such patients, chronic bacterial colonisation of the airways develops, usually with the intransigent bacterial pathogens S. aureus and P. aeruginosa. Sputum viscosity is caused by extracellular DNA released from invading inflammatory cells, much of which is believed to originate from NETosis. This contention is supported by the observation that neutrophil elastase and MPO, which are found in high concentration in CF sputum, are bound to DNA, a key molecular signature of NETs.81 If detached from NETs, neutrophil-derived proteolytic enzymes may also damage components of pulmonary connective tissue, especially elastin, compromising airway elasticity and function which may underpin the correlation between the magnitude of NET formation in the airways of CF patients and the degree of impairment of lung function.74,82
Pharmacological control of NETosis
Given the apparent involvement of excessive NETosis in the development of autoimmune and cardiovascular diseases, as well as in exacerbation of CF, pharmacological regulation of aberrant NETosis has definite therapeutic potential. This potential has yet to be realised, however, due in large part to the relatively recent discovery of NETosis, as well as the current limited insights into the diversity of the molecular mechanisms underpinning this process. Possible strategies include: (1) inhibition of the generation of pro-NETotic ROS, or, alternatively, neutralisation of ROS using oxidant-scavengers such as N-acetylcysteine or ascorbic acid;11 (2) inhibitors of the activation of NFkB such as ascorbic acid and acetylsalicylic acid (aspirin), both of which have shown promise in experimental animal models of excessive NET formation11,83; and (3) inhibitors of PAD4 which are currently in pre-clinical development84.
With respect to alternative therapies, inhaled recombinant human DNase is widely used in the treatment of CF, primarily as a strategy to degrade neutrophil-derived DNA, a significant contributor to the viscosity of airway mucus.85
Conclusions
NETs appear to increase the versatility and potency of the anti-infective armamentarium of neutrophils, as well as several other cell types of the innate immune system, possibly prolonging protective activity beyond cell death, thereby ensuring maximal utilisation of antimicrobial granule proteins. However, several important questions relating to the exact role of NETs in host defence remain incompletely understood. Remaining avenues for exploration include: (1) determining the biological relevance of the various types of NETosis; (2) unravelling the precise molecular and biochemical mechanisms underpinning these processes; and (3) characterising cooperative, beneficial interactions of NETosis with other cellular and humoral components of the innate and adaptive immune systems. As with other indiscriminate phagocyte-derived antimicrobial systems, such as the generation of tissue damaging and carcinogenic ROS, the extracellular release of nuclear material and enzymes such as elastase and MPO during NETosis presents the potential threat of development of autoimmune, cardiovascular and other disorders. Balancing NETosis in favour of host defence using pharmacological and other strategies represents an ongoing challenge.
Authors' contributions
All the authors contributed equally to the compilation of the manuscript, as well as to the coordination of the final version.
References
1. Opitz B, Van Laak V,Eitel J, Suttorp N. Innate immune recognition in infectious and noninfectious diseases of the lung. Am J Respir Crit Care Med. 2010;181(12):1294-1309. http://dx.doi.org/10.1164/rccm.200909-1427SO [ Links ]
2. Barber GN. Cytoplasmic DNA innate immune pathways. Immunol Rev. 2011;243(1):99-108. http://dx.doi.org/10.1111/j.1600-065X.2011.01051.x [ Links ]
3. Brinkmann V,Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532-1535. http://dx.doi.org/10.1126/science.1092385 [ Links ]
4. Pillay J, Den Braber I, Vrisekoop N, Kwast LM, De Boer RJ, Borghans JA, et al. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood. 2010;116(4):625-627. http://dx.doi.org/10.1182/blood-2010-01-259028 [ Links ]
5. Robb CT, Dyrynda EA, Gray RD, Rossi AG, Smith VJ. Invertebrate extracellular phagocyte traps show that chromatin is an ancient defence weapon. Nat Commun. 2014;5:4627. http://dx.doi.org/10.1038/ncomms5627 [ Links ]
6. Pilsczek FH, Salina D, Poon KK, Fahey C, Yipp BG, Sibley CD, et al. A novel mechanism of rapid nuclear neutrophil extracellular trap formation in response to Staphylococcus aureus. J Immunol. 2010;185(12):7413-7425. http://dx.doi.org/10.4049/jimmunol.1000675 [ Links ]
7. Parker H, Dragunow M, Hampton MB, Kettle AJ, Winterbourn CC. Requirements for NADPH oxidase and myeloperoxidase in neutrophil extracellular trap formation differ depending on the stimulus. J Leukoc Biol. 2012;92(4):841-849. http://dx.doi.org/10.1189/jlb.1211601 [ Links ]
8. Yousefi S, Mihalache C, Kozlowski E, Schmid I, Simon HU. Viable neutrophils release mitochondrial DNA to form neutrophil extracellular traps. Cell Death Differ. 2009;16(11):1438-1444. http://dx.doi.org/10.1038/cdd.2009.96 [ Links ]
9. Fuchs TA, Abed U, Goosmann C, Hurwitz R, Schulze I, Wahn V, et al. Novel cell death program leads to neutrophil extracellular traps. J Cell Biol. 2007;176(2):231-241. http://dx.doi.org/10.1083/jcb.200606027 [ Links ]
10. Hakkim A, Fuchs TA, Martinez NE, Hess S, Prinz H, Zychlinsky A, et al. Activation of the Raf-MEK-ERK pathway is required for neutrophil extracellular trap formation. Nat Chem Biol. 2011;7(2):75-77. http://dx.doi.org/10.1038/nchembio.496 [ Links ]
11. Mohammed BM, Fisher BJ, Kraskauskas D, Farkas D, Brophy DF, Fowler AA, et al. Vitamin C: A novel regulator of neutrophil extracellular trap formation. Nutrients. 2013;5(8):3131-3151. http://dx.doi.org/10.3390/nu5083131 [ Links ]
12. Wang Y, Li M, Stadler S, Correll S, Li P Wang D, et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extracellular trap formation. J Cell Biol. 2009;184(2):205-213. http://dx.doi.org/10.1083/jcb.200806072 [ Links ]
13. Leshner M, Wang S, Lewis C, Zheng H, Chen XA, Santy L, et al. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap-like structures. Front Immunol. 2012;3, Art. #307, 11 pages. http://dx.doi.org/10.3389/fimmu.2012.00307 [ Links ]
14. Metzler KD, Goosmann C, Lubojemska A, Zychlinsky A, Papayannopoulos V. A myeloperoxidase-containing complex regulates neutrophil elastase release and actin dynamics during NETosis. Cell Rep. 2014;8(3):883-896. http://dx.doi.org/10.1016/j.celrep.2014.06.044 [ Links ]
15. Neumann A, Berends ET, Nerlich A, Molhoek EM, Gallo RL, Meerloo T, et al. The antimicrobial peptide LL-37 facilitates the formation of neutrophil extracellular traps. Biochem J. 2014;464(1):3-11. http://dx.doi.org/10.1042/BJ20140778 [ Links ]
16. Neumann A, Vollger L, Berends ET, Molhoek EM, Stapels DA, Midon M, et al. Novel role of the antimicrobial peptide LL-37 in the protection of neutrophil extracellular traps against degradation by bacterial nucleases. J Innate Immun. 2014;6(6):860-868. http://dx.doi.org/10.1159/000363699 [ Links ]
17. Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM, et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat Med. 2007;13(4):463-469. http://dx.doi.org/10.1038/nm1565 [ Links ]
18. Yipp BG, Petri B, Salina D, Jenne CN, Scott BN, Zbytnuik LD, et al. Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Nat Med. 2012;18(9):1386-1393. http://dx.doi.org/10.1038/nm.2847 [ Links ]
19. Byrd AS, O'Brien XM, Johnson CM, Lavigne LM, Reichner JS. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J Immunol. 2013;190(8):4136-4148. http://dx.doi.org/10.4049/jimmunol.1202671 [ Links ]
20. Yipp BG, Kubes P NETosis: How vital is it? Blood. 2013;122(16):2784-2794. http://dx.doi.org/10.1182/blood-2013-04-457671 [ Links ]
21. Arai Y Nishinaka Y Arai T, Morita M, Mizugishi K, Adachi S, et al. Uric acid induces NADPH oxidase-independent neutrophil extracellular trap formation. Biochem Biophys Res Commun. 2014;443(2):556-561. http://dx.doi.org/10.1016/j.bbrc.2013.12.007 [ Links ]
22. Itagaki K, Kaczmarek E, Lee YT, Tang IT, Isal B, Adibnia Y et al. Mitochondrial DNA released by trauma induces neutrophil extracellular traps. PLoS One. 2015;10(3), e0120549, 10 pages. http://dx.doi.org/10.1371/journal.pone.0120549 [ Links ]
23. Hirsch JG. Bactericidal action of histone. J Exp Med. 1958;108(6):925-944. http://dx.doi.org/10.1084/jem.108.6.925 [ Links ]
24. Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W, et al. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009;5(10), e1000639, 18 pages. http://dx.doi.org/10.1371/journal.ppat.1000639 [ Links ]
25. Halverson TW, Wilton M, Poon KK, Petri B, Lewenza S. DNA is an antimicrobial component of neutrophil extracellular traps. PLoS Pathog. 2015;11(1), e1004593, 23 pages. http://dx.doi.org/10.1371/journal.ppat.1004593 [ Links ]
26. Rahman S, Gadjeva M. Does NETosis contribute to the bacterial pathoadaptation in cystic fibrosis? Front Immunol. 2014;5, Art. #378, 6 pages. http://dx.doi.org/10.3389/fimmu.2014.00378 [ Links ]
27. Young RL, Malcolm KC, Kret JE, Caceres SM, Poch KR, Nichols DP et al. Neutrophil extracellular trap (NET)-mediated killing of Pseudomonas aeruginosa: Evidence of acquired resistance within the CF airway, independent of CFTR. PLoS One. 2011;6(9), e23637, 10 pages. http://dx.doi.org/10.1371/journal.pone.0023637 [ Links ]
28. Menten-Dedoyart C, Faccinetto C, Golovchenko M, Dupiereux I, Van Lerberghe PB, Dubois S, et al. Neutrophil extracellular traps entrap and kill Borrelia burgdorferi sensu stricto spirochetes and are not affected by Ixodes ricinus tick saliva. J Immunol. 2012;189(11):5393-5401. http://dx.doi.org/10.4049/jimmunol.1103771 [ Links ]
29. Riyapa D, Buddhisa S, Korbsrisate S, Cuccui J, Wren BW, Stevens MP et al. Neutrophil extracellular traps exhibit antibacterial activity against Burkholderia pseudomallei and are influenced by bacterial and host factors. Infect Immun. 2012;80(11):3921-3929. http://dx.doi.org/10.1128/IAI.00806-12 [ Links ]
30. Ramos-Kichik V Mondragon-Flores R, Mondragon-Castelan M, Gonzalez-Pozos S, Muniz-Hernandez S, Rojas-Espinosa O, et al. Neutrophil extracellular traps are induced by Mycobacterium tuberculosis. Tuberculosis (Edinb). 2009;89(1):29-37. http://dx.doi.org/10.1016/j.tube.2008.09.009 [ Links ]
31. Branzk N, Lubojemska A, Hardison SE, Wang Q, Gutierrez MG, Brown GD, et al. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat Immunol. 2014;15(11):1017-1025. http://dx.doi.org/10.1038/ni.2987 [ Links ]
32. Sumby P, Barbian KD, Gardner DJ, Whitney AR, Welty DM, Long RD, et al. Extracellular deoxyribonuclease made by group A Streptococcus assists pathogenesis by enhancing evasion of the innate immune response. Proc Natl Acad Sci USA. 2005;102(5):1679-1684. http://dx.doi.org/10.1073/pnas.0406641102 [ Links ]
33. Beiter K, Wartha F, Albiger B, Normark S, Zychlinsky A, Henriques-Normark B. An endonuclease allows Streptococcus pneumoniae to escape from neutrophil extracellular traps. Curr Biol. 2006;16(4):401-407. http://dx.doi.org/10.1016/j.cub.2006.01.056 [ Links ]
34. Khatua B, Bhattacharya K, Mandal C. Sialoglycoproteins adsorbed by Pseudomonas aeruginosa facilitate their survival by impeding neutrophil extracellular trap through siglec-9. J Leukoc Biol. 2012;91(4):641-655. http://dx.doi.org/10.1189/jlb.0511260 [ Links ]
35. Bruns S, Kniemeyer O, Hasenberg M, Aimanianda V Nietzsche S, Thywissen A, et al. Production of extracellular traps against Aspergillus fumigatus in vitro and in infected lung tissue is dependent on invading neutrophils and influenced by hydrophobin RodA. PLoS Pathog. 2010;6(4), e1000873, 18 pages. http://dx.doi.org/10.1371/journal.ppat.1000873 [ Links ]
36. Wartha F, Beiter K, Albiger B, Fernebro J, Zychlinsky A, Normark S, et al. Capsule and D-alanylated lipoteichoic acids protect Streptococcus pneumoniae against neutrophil extracellular traps. Cell Microbiol. 2007;9(5):1162-1171. http://dx.doi.org/10.1111/j.1462-5822.2006.00857.x [ Links ]
37. Bianchi M, Niemiec MJ, Siler U, Urban CF, Reichenbach J. Restoration of anti-Aspergillus defense by neutrophil extracellular traps in human chronic granulomatous disease after gene therapy is calprotectin-dependent. J Allergy Clin Immunol. 2011;127(5):1243-1252e7. http://dx.doi.org/10.1016/j.jaci.2011.01.021 [ Links ]
38. Shan Q, Dwyer M, Rahman S, Gadjeva M. Distinct susceptibilities of corneal Pseudomonas aeruginosa clinical isolates to neutrophil extracellular trap-mediated immunity. Infect Immun. 2014;82(10):4135-4143. http://dx.doi.org/10.1128/IAI.02169-14 [ Links ]
39. Yoo DG, Floyd M, Winn M, Moskowitz SM, Rada B. NET formation induced by Pseudomonas aeruginosa cystic fibrosis isolates measured as release of myeloperoxidase-DNA and neutrophil elastase-DNA complexes. Immunol Lett. 2014;160(2):186-194. http://dx.doi.org/10.1016/jJmlet.2014.03.003 [ Links ]
40. Thammavongsa V, Missiakas DM, Schneewind O. Staphylococcus aureus degrades neutrophil extracellular traps to promote immune cell death. Science. 2013;342(6160):863-866. http://dx.doi.org/10.1126/science.1242255 [ Links ]
41. Lappann M, Danhof S, Guenther F, Olivares-Florez S, Mordhorst IL, Vogel U. In vitro resistance mechanisms of Neisseria meningitidis against neutrophil extracellular traps. Mol Microbiol. 2013;89(3):433-449. http://dx.doi.org/10.1111/mmi.12288 [ Links ]
42. Juneau RA, Stevens JS, Apicella MA, Criss AK. A thermonuclease of Neisseria gonorrhoeae enhances bacterial escape from killing by neutrophil extracellular traps. J Infect Dis. 2015;212(2):316-324. http://dx.doi.org/10.1093/infdis/jiv031 [ Links ]
43. Gunderson CW, Seifert HS. Neisseria gonorrhoeae elicits extracellular traps in primary neutrophil culture while suppressing the oxidative burst. MBio. 2015;6(1), e02452-14, 10 pages. http://dx.doi.org/10.1128/mBio.02452-14 [ Links ]
44. Seper A, Hosseinzadeh A, Gorkiewicz G, Lichtenegger S, Roier S, Leitner DR, et al. Vibrio cholerae evades neutrophil extracellular traps by the activity of two extracellular nucleases. PLoS Pathog. 2013;9(9), e1003614, 15 pages. http://dx.doi.org/10.1371/journal.ppat.1003614 [ Links ]
45. Guimaraes-Costa AB, DeSouza-Vieira TS, Paletta-Silva R, Freitas-Mesquita AL, Meyer-Fernandes JR, Saraiva EM. 3'-nucleotidase/nuclease activity allows Leishmania parasites to escape killing by neutrophil extracellular traps. Infect Immun. 2014;82(4):1732-1740. http://dx.doi.org/10.1128/IAI.01232-13 [ Links ]
46. Abi Abdallah DS, Lin C, Ball CJ, King MR, Duhamel GE, Denkers EY. Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps. Infect Immun. 2012;80(2):768-777. http://dx.doi.org/10.1128/IAI.05730-11 [ Links ]
47. Baker VS, Imade GE, Molta NB, Tawde P, Pam SD, Obadofin MO, et al. Cytokine-associated neutrophil extracellular traps and antinuclear antibodies in Plasmodium falciparum infected children under six years of age. Malar J. 2008;7, Art. #41, 12 pages. http://dx.doi.org/10.1186/1475-2875-7-41 [ Links ]
48. Urban CF, Reichard U, Brinkmann V, Zychlinsky A. Neutrophil extracellular traps capture and kill Candida albicans yeast and hyphal forms. Cell Microbiol. 2006;8(4):668-676. http://dx.doi.org/10.1111/j.1462-5822.2005.00659.x [ Links ]
49. Kamoshida G, Kikuchi-Ueda T, Tansho-Nagakawa S, Nakano R, Nakano A, Kikuchi H, et al. Acinetobacter baumannii escape from neutrophil extracellular traps (NETs). J Infect Chemother. 2015;21(1):43-49. http://dx.doi.org/10.1016/j.jiac.2014.08.032 [ Links ]
50. Rocha JD, Nascimento MT, Decote-Ricardo D, Corte-Real S, Morrot A, Heise N, et al. Capsular polysaccharides from Cryptococcus neoformans modulate production of neutrophil extracellular traps (NETs) by human neutrophils. Sci Rep. 2015;5, Art. #8008, 10 pages. http://dx.doi.org/10.1038/srep08008 [ Links ]
51. Menegazzi R, Decleva E, Dri P. Killing by neutrophil extracellular traps: Fact or folklore? Blood. 2012;119(5):1214-1216. http://dx.doi.org/10.1182/blood-2011-07-364604 [ Links ]
52. Farrera C, Fadeel B. Macrophage clearance of neutrophil extracellular traps is a silent process. J Immunol. 2013;191(5):2647-2656. http://dx.doi.org/10.4049/jimmunol.1300436 [ Links ]
53. Wang H, Wang C, Zhao MH, Chen M. Neutrophil extracellular traps can activate alternative complement pathways. Clin Exp Immunol. 2015;181(3):518-527. http://dx.doi.org/10.1111/cei.12654 [ Links ]
54. Saitoh T, Komano J, Saitoh Y, Misawa T, Takahama M, Kozaki T, et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host Microbe. 2012;12(1):109-116. http://dx.doi.org/10.1016/j.chom.2012.05.015 [ Links ]
55. Yost CC, Cody MJ, Harris ES, Thornton NL, McInturff AM, Martinez ML, et al. Impaired neutrophil extracellular trap (NET) formation: A novel innate immune deficiency of human neonates. Blood. 2009;113(25):6419-6427. http://dx.doi.org/10.1182/blood-2008-07-171629 [ Links ]
56. Hazeldine J, Harris P, Chapple IL, Grant M, Greenwood H, Livesey A, et al. Impaired neutrophil extracellular trap formation: A novel defect in the innate immune system of aged individuals. Aging Cell. 2014;13(4):690-698. http://dx.doi.org/10.1111/acel.12222 [ Links ]
57. Yu Y, Su K. Neutrophil extracellular traps and systemic lupus erythematosus. J Clin Cell Immunol. 2013;4(2), Art. #139, 7 pages. http://dx.doi.org/10.4172/2155-9899.1000139 [ Links ]
58. Leffler J, Gullstrand B, Jonsen A, Nilsson JA, Martin M, Blom AM, et al. Degradation of neutrophil extracellular traps co-varies with disease activity in patients with systemic lupus erythematosus. Arthritis Res Ther. 2013;15(4), R84, 9 pages. http://dx.doi.org/10.1186/ar4264 [ Links ]
59. Leffler J, Martin M, Gullstrand B, Tyden H, Lood C, Truedsson L, et al. Neutrophil extracellular traps that are not degraded in systemic lupus erythematosus activate complement exacerbating the disease. J Immunol. 2012;188(7):3522-3531. http://dx.doi.org/10.4049/jimmunol.1102404 [ Links ]
60. Hakkim A, Furnrohr BG, Amann K, Laube B, Abed UA, Brinkmann V, et al. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc Natl Acad Sci USA. 2010;107(21):9813-9818. http://dx.doi.org/10.1073/pnas.0909927107 [ Links ]
61. Carmona-Rivera C, Kaplan MJ. Low-density granulocytes: A distinct class of neutrophils in systemic autoimmunity. Semin Immunopathol. 2013;35(4):455-463. http://dx.doi.org/10.1007/s00281-013-0375-7 [ Links ]
62. Khandpur R, Carmona-Rivera C, Vivekanandan-Giri A, Gizinski A, Yalavarthi S, Knight JS, et al. NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci Transl Med. 2013;5(178):178ra40. http://dx.doi.org/10.1126/scitranslmed.3005580 [ Links ]
63. Papadaki G, Choulaki C, Mitroulis I, Verginis P Repa A, Raptopoulou A, et al. Enhanced release of neutrophil extracellular traps from peripheral blood neutrophils in patients with rheumatoid arthritis. Ann Rheum Dis. 2012;71(Suppl 1):A79. http://dx.doi.org/10.1136/annrheumdis-2011-201238.17 [ Links ]
64. Zawrotniak M, Rapala-Kozik M. Neutrophil extracellular traps (NETs) -Formation and implications. Acta Biochim Pol. 2013;60(3):277-284. [ Links ]
65. Kessenbrock K, Krumbholz M, Schonermarck U, Back W, Gross WL, Werb Z, et al. Netting neutrophils in autoimmune small-vessel vasculitis. Nat Med. 2009;15(6):623-625. http://dx.doi.org/10.1038/nm.1959 [ Links ]
66. Saffarzadeh M, Juenemann C, Queisser MA, Lochnit G, Barreto G, Galuska SP et al. Neutrophil extracellular traps directly induce epithelial and endothelial cell death: A predominant role of histones. PLoS One. 2012;7(2), e32366, 14 pages. http://dx.doi.org/10.1371/journal.pone.0032366 [ Links ]
67. Fuchs TA, Brill A, Wagner DD. Neutrophil extracellular trap (NET) impact on deep vein thrombosis. Arterioscler Thromb Vasc Biol. 2012;32(8):1777-1783. http://dx.doi.org/10.1161/ATVBAHA.111.242859 [ Links ]
68. Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD Jr., et al. Extracellular DNA traps promote thrombosis. Proc Natl Acad Sci USA. 2010;107(36):15880-15885. http://dx.doi.org/10.1073/pnas.1005743107 [ Links ]
69. Saffarzadeh M, Preissner KT. Fighting against the dark side of neutrophil extracellular traps in disease: Manoeuvres for host protection. Curr Opin Hematol. 2013;20(1):3-9. http://dx.doi.org/10.1097/MOH.0b013e32835a0025 [ Links ]
70. Martinod K, Wagner DD. Thrombosis: Tangled up in NETs. Blood. 2014;123(18):2768-2776. http://dx.doi.org/10.1182/blood-2013-10-463646 [ Links ]
71. Brill A, Fuchs TA, Savchenko AS, Thomas GM, Martinod K, De Meyer SF, et al. Neutrophil extracellular traps promote deep vein thrombosis in mice. J Thromb Haemost. 2012;10(1):136-144. http://dx.doi.org/10.1111/j.1538-7836.2011.04544.x [ Links ]
72. Von Bruhl ML, Stark K, Steinhart A, Chandraratne S, Konrad I, Lorenz M, et al. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J Exp Med. 2012;209(4):819-835. http://dx.doi.org/10.1084/jem.20112322 [ Links ]
73. Savchenko AS, Martinod K, Seidman MA, Wong SL, Borissoff JI, Piazza G, et al. Neutrophil extracellular traps form predominantly during the organizing stage of human venous thromboembolism development. J Thromb Haemost. 2014;12(6):860-870. http://dx.doi.org/10.1111/jth.12571 [ Links ]
74. Marcos V Zhou Z, Yildirim AO, Bohla A, Hector A, Vitkov L, et al. CXCR2 mediates NADPH oxidase-independent neutrophil extracellular trap formation in cystic fibrosis airway inflammation. Nat Med. 2010;16(9):1018-1023. http://dx.doi.org/10.1038/nm.2209 [ Links ]
75. Cheng OZ, Palaniyar N. NET balancing: A problem in inflammatory lung diseases. Front Immunol. 2013;4, Art. #1, 13 pages. http://dx.doi.org/10.3389/fimmu.2013.00001 [ Links ]
76. Dworski R, Simon HU, Hoskins A, Yousefi S. Eosinophil and neutrophil extracellular DNA traps in human allergic asthmatic airways. J Allergy Clin Immunol. 2011;127(5):1260-1266. http://dx.doi.org/10.1016/j.jaci.2010.12.1103 [ Links ]
77. Grabcanovic-Musija F, Obermayer A, Stoiber W, Krautgartner WD, Steinbacher P, Winterberg N, et al. Neutrophil extracellular trap (NET) formation characterises stable and exacerbated COPD and correlates with airflow limitation. Respir Res. 2015;16, Art. #59, 12 pages. http://dx.doi.org/10.1186/s12931-015-0221-7 [ Links ]
78. Dushianthan A, Grocott MP, Postle AD, Cusack R. Acute respiratory distress syndrome and acute lung injury. Postgrad Med J. 2011;87(1031):612-622. http://dx.doi.org/10.1136/pgmj.2011.118398 [ Links ]
79. Summers C, Singh NR, White JF, Mackenzie IM, Johnston A, Solanki C, et al. Pulmonary retention of primed neutrophils: A novel protective host response, which is impaired in the acute respiratory distress syndrome. Thorax. 2014;69(7):623-629. http://dx.doi.org/10.1136/thoraxjnl-2013-204742 [ Links ]
80. Khatri IA, Bhaskar KR, Lamont JT, Sajjan SU, Ho CK, Forstner J. Effect of chondroitinase ABC on purulent sputum from cystic fibrosis and other patients. Pediatr Res. 2003;53(4):619-627. http://dx.doi.org/10.1203/01.PDR.0000054780.11755.B9 [ Links ]
81. Papayannopoulos V, Staab D, Zychlinsky A. Neutrophil elastase enhances sputum solubilization in cystic fibrosis patients receiving DNase therapy. PLoS One. 2011;6(12), e28526, 7 pages. http://dx.doi.org/10.1371/journal.pone.0028526 [ Links ]
82. Dwyer M, Shan Q, D'Ortona S, Maurer R, Mitchell R, Olesen H, et al. Cystic fibrosis sputum DNA has NETosis characteristics and neutrophil extracellular trap release is regulated by macrophage migration-inhibitory factor. J Innate Immun. 2014;6(6):765-779. http://dx.doi.org/10.1159/000363242 [ Links ]
83. Lapponi MJ, Carestia A, Landoni VI, Rivadeneyra L, Etulain J, Negrotto S, et al. Regulation of neutrophil extracellular trap formation by anti-inflammatory drugs. J Pharmacol Exp Ther. 2013;345(3):430-437. http://dx.doi.org/10.1124/jpet.112.202879 [ Links ]
84. Bozdag M, Dreker T, Henry C, Tosco P, Vallaro M, Fruttero R, et al. Novel small molecule protein arginine deiminase 4 (PAD4) inhibitors. Bioorg Med Chem Lett. 2013;23(3):715-719. http://dx.doi.org/10.1016/j.bmcl.2012.11.102 [ Links ]
85. Bakker EM, Volpi S, Salonini E, Van der Wiel-Kooij EC, Sintnicolaas CJ, Hop WC, et al. Improved treatment response to dornase alfa in cystic fibrosis patients using controlled inhalation. Eur Respir J. 2011;38(6):1328-1335. http://dx.doi.org/10.1183/09031936.00006211 [ Links ]
 Correspondence:
Correspondence:
Jan Nel
Department of Haematology, Faculty of Health Sciences, University of Pretoria
Private Bag X323, Arcadia 0007, South Africa
Jan.Nel@up.ac.za
Received: 20 Feb. 2015
Revised: 08 June 2015
Accepted: 18 June 2015


{kind=link}
{kind=link}
{kind=link}