










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.111 n.11-12 Pretoria Nov./Dec. 2015
http://dx.doi.org/10.17159/sajs.2015/20150261
RESEARCH LETTER
Insecticide resistance in the malaria vector Anopheles arabiensis in Mamfene, KwaZulu-Natal
Basil D. BrookeI, II; Leanne RobertsonI; Maria L. KaiserI, II; Eric RaswiswiIII; Givemore MunhengaI, II; Nelius VenterI, II; Oliver R. WoodI, II; Lizette L. KoekemoerI, II
ICentre for Opportunistic, Tropical & Hospital Infections, National Institute for Communicable Diseases, Johannesburg, South Africa
IIWits Research Institute for Malaria, School of Pathology, Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa
IIIKwaZulu-Natal Department of Health, KwaZulu-Natal Provincial Government, Jozini, South Africa
ABSTRACT
The control of malaria vector mosquitoes in South Africa's affected provinces is primarily based on indoor spraying of long-lasting residual insecticides. The primary vectors in South Africa are Anopheles arabiensis and An. funestus. South Africa's National Malaria Control Programme has adopted a malaria elimination agenda and has scaled up vector control activities accordingly. However, despite these plans, local transmission continues and is most likely because of outdoor feeding by populations of An. arabiensis. An outdoor Anopheles surveillance system has been set up in three sections of the Mamfene district in northern KwaZulu-Natal in order to assess the extent of outdoor resting An. arabiensis in Mamfene and to assess the current insecticide susceptibility status of this population. According to WHO criteria, the An. arabiensis samples tested showed evidence of resistance to deltamethrin (pyrethroid), DDT (organochlorine) and bendiocarb (carbamate), and full susceptibility to the organophosphates pirimiphos-methyl and fenitrothion. Pre-exposure to piperonyl butoxide completely nullified the deltamethrin resistance otherwise evident in these samples, supporting previous studies implicating monooxygenase-based detoxification as the primary mechanism of pyrethroid resistance. The data presented here affirm the presence of pyrethroid and DDT resistance previously detected in this population and also indicate the comparatively recent emergence of resistance to the carbamate insecticide bendiocarb. These data show that special attention and commitment needs to be given to the principles of insecticide resistance management as well as to investigations into alternative control techniques designed to target outdoor-resting An. arabiensis in northern KwaZulu-Natal.
Keywords: malaria vector control; malaria elimination; vector surveillance; South Africa; resistance management
Introduction
The control of malaria vector mosquitoes in South Africa's affected provinces is primarily based on indoor spraying of long-lasting residual insecticides.1 The indoor residual spraying (1RS) method has been the mainstay of malaria vector control in South Africa since the late 1940s and has remained effective owing to carefully co-ordinated 1RS programmes in South Africa's Limpopo, Mpumalanga and KwaZulu-Natal Provinces.2
Only Anopheles mosquitoes can transmit human malaria parasites and the primary vectors in South Africa are Anopheles arabiensis and An. funestus.1Of these, An. funestus is almost entirely anthropophilic (human biting), endophagic (indoor feeding) and endophilic (indoor resting).3 These characteristics make this species especially susceptible to control by 1RS, assuming that the insecticide employed for this purpose is effective against the target An. funestus population. Control by 1RS means that the mosquitoes must retain complete or near complete susceptibility to the insecticide class being used, which can only be ascertained by regular monitoring and surveillance. The malaria epidemic experienced in South Africa during the period 1996-2000 was largely the result of the development of resistance to pyrethroid insecticides in populations of this species in northern KwaZulu-Natal and Mpumalanga which led to vector control failure.1,2 Control was re-established using a mosaic resistance management system which was later drafted into a World Health Organization (WHO) document -- the Global Plan for Insecticide Resistance Management (GPIRM).4 South Africa currently subscribes to the principles outlined in GPIRM as part of its malaria elimination agenda.5 However, despite these plans and the scaling up of vector control activities in South Africa, local transmission continues, most likely because of outdoor transmission by populations of An. arabiensis. Unlike An. funestus, An. arabiensis has evolved substantial behavioural plasticity and will feed and rest indoors and outdoors, and will feed on humans as well as livestock, especially bovines.3Anopheles arabiensis is therefore substantially less susceptible to control by IRS.
Recently, a project was launched to assess the feasibility of the sterile insect technique for malaria vector control in South Africa, with special emphasis on controlling outdoor transmission by An. arabiensis.6As part of the baseline survey linked to this project, an outdoor Anopheles surveillance system has been set up in three sections of the Mamfene district in northern KwaZulu-Natal. This surveillance system has enabled recent assessments of insecticide resistance in outdoor-resting An. arabiensis in Mamfene as a follow-up to the discovery of pyrethroid resistance in this region in 2005.7
Methods
In order to assess the current insecticide susceptibility status of outdoor resting An. arabiensis in Mamfene, adult Anopheles mosquitoes were collected from outdoor-placed ceramic pots8 and modified plastic buckets deployed in 20 households in Mamfene Sections 2, 8 and 9 during March and April 2015. These collections were transported live to the Botha De Meillon insectary facility at the National Institute for Communicable Diseases (NICD) in Johannesburg. Blood-fed female specimens were individually placed in egg-laying vials so that eggs could be harvested and reared by family. All wild-caught female individuals, including those that produced eggs, were identified to species group using morphological keys9,10 and to species by standard PCR11. A total of 35 families identified as An. arabiensis was pooled and the F1 progeny were reared to adults under standard insectary conditions of 25 °C and 80% relative humidity.12
Samples of 2-5-day-old female adult F1 progeny were assessed for their susceptibility to diagnostic concentrations of a range of insecticides according to the standard WHO bioassay method.13 Controls included samples of 2-4-day-old F1 male adults exposed to untreated papers. In addition, a subset of samples was used to assess the effect of preexposure to the insecticide synergist piperonyl butoxide (PBO) on the expression of pyrethroid resistance according to a method previously described by Brooke et al.14
Results and discussion
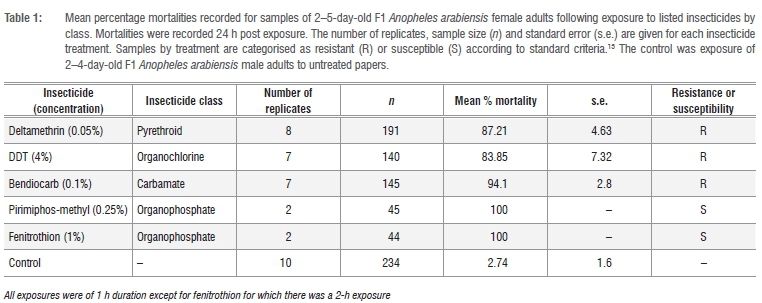
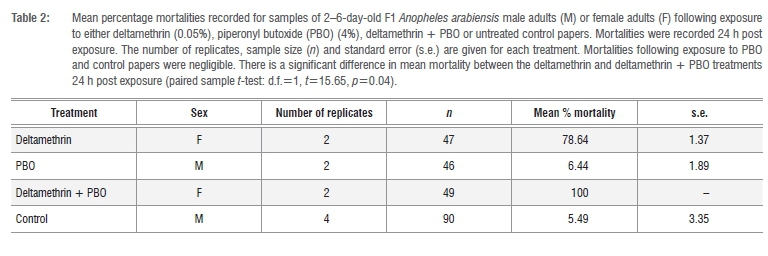
According to WHO criteria,13 the F1 An. arabiensis samples tested showed evidence of resistance to deltamethrin (pyrethroid), DDT (organo-chlorine) and bendiocarb (carbamate), and full susceptibility to the organophosphates pirimiphos-methyl and fenitrothion (Table 1). Preexposure to PBO completely nullified the deltamethrin resistance otherwise evident in these samples (paired sample f-test: d.f. = 1, t = 15.65, p= 0.04) (Table 2).
The first assessments of resistance in An. arabiensis at Mamfene were conducted in 1996 and no resistance phenotypes were recorded.15 However, subsequent samples collected in 2002 indicated the emergence of resistance to DDT16 which was again recorded in 2005 together with the first indication of pyrethroid resistance7. The 2015 data presented here affirm the presence of pyrethroid and DDT resistance in this population, albeit at a low frequency, and also indicate the comparatively recent emergence of resistance to the carbamate insecticide bendiocarb. The PBO exposure data support previous analyses implicating monooxygenase-mediated detoxification as the primary mode of pyrethroid resistance in An. arabiensis at Mamfene,7,17,18 because PBO enhances insecticide toxicity by providing an alternative substrate for monooxygenases. PBO pre-exposure assays can therefore be used to elucidate monooxygenase-based resistance mechanisms.
Conclusion
South Africa's 1996 to 2000 malaria epidemic illustrates the effect that a single insecticide resistance phenotype (pyrethroid resistance in An. funestus) can have on an IRS-based vector control programme.2,19,20 The occurrence of multiple vector species carrying multiple resistance mechanisms coupled to ongoing outdoor transmission in northern KwaZulu-Natal means that special attention and commitment needs to be given to the principles of insecticide resistance management as outlined in the GPIRM document4 as well as to investigations into alternative control techniques designed to target outdoor-resting An. arabiensis.
Acknowledgements
We thank Mr Sifiso Ngxongo and Mr Jabulani Zikhali for their assistance with the collection of specimens and Prof. Maureen Coetzee for her comments on the manuscript. This work was funded by the Nuclear Technologies in Medicine and the Bioscience Initiatives (NTeMBI), a national platform developed and managed by the South African Nuclear Energy Corporation and supported by the Department of Science and Technology. Funding was also provided in part by the National Research Foundation and by the International Atomic Energy Agency ((Contract no SAF16780 (under the G34002)) as well as by a CDC/GDD (Global Diseases Detection programme) grant to B.D.B. (U19GH000622-01 MAL01).
Authors' contributions
B.D.B. assisted with data analysis and interpretation and produced the manuscript. L.R., N.V. and O.R.W. assisted with specimen collection, species identification and experimental procedures. M.L.K. and G.M. assisted with the experimental procedures and data analysis. E.R. assisted with data interpretation and the drafting of the manuscript. L.L.K. conceived the project, assisted with experimental procedures, data analysis and interpretation and the drafting of the manuscript. All authors read and approved the final draft of the manuscript.
References
1. Brooke B, Koekemoer L, Kruger P Urbach J, Misiani E, Coetzee M. Malaria vector control in South Africa. S Afr Med J. 2013;103(10 Suppl 2):784-788. http://dx.doi.org/10.7196/samj.7447 [ Links ]
2. Coetzee M, Kruger P Hunt RH, Durrheim DN, Urbach J, Hansford CF. Malaria in South Africa: 110 years of learning to control the disease. S Afr Med J. 2013;103(10 Suppl 2):770-778. http://dx.doi.org/10.7196/samj.7446 [ Links ]
3. Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: Occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117. http://dx.doi.org/10.1186/1756-3305-3-117 [ Links ]
4. World Health Organization (WHO). Global Plan for insecticide resistance management. Geneva: WHO; 2012. Available from: http://www.who.int/malaria/vector_control/ivm/gpirm/en/ [ Links ]
5. Maharaj R, Morris N, Seocharan I, Kruger P Moonasar D, Mabuza A, et al. The feasibility of malaria elimination in South Africa. Malar J. 2012;11:423. http://dx.doi.org/10.1186/ 1475-2875-11-423 [ Links ]
6. Munhenga G, Brooke BD, Chirwa TF, Hunt RH, Coetzee M, Koekemoer LL. Evaluating the potential of the sterile insect technique for malaria control: Relative fitness and mating compatibility between laboratory colonized and a wild population of Anopheles arabiensis from the Kruger National Park, South Africa. Parasit Vectors. 2011;4:208. http://dx.doi.org/10.1186/1756-3305-4-208 [ Links ]
7. Mouatcho JC, Munhenga G , Hargreaves K, Brooke BD, Coetzee M, Koekemoer LL. Pyrethroid resistance in a major African malaria vector Anopheles arabiensis from Mamfene, northern Kwazulu-Natal, South Africa. S Afr J Sci. 2009;105:127-131. [ Links ]
8. Brooke B, Hargreaves K, Spillings B, Raswiswi E, Koekemoer L, Coetzee M. Novel malaria control: Can traditional clay pots be used to deliver entomopathogenic fungi to malaria vectors in northern Kwazulu-Natal? Communicable Diseases Surveillance Bulletin. 2012;10:13-15. [ Links ]
9. Gillies MT, De Meillon B. The Anophelinae of Africa south of the Sahara. Johannesburg: South African Institute for Medical Research; 1968. [ Links ]
10. Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara. Johannesburg: South African Institute for Medical Research; 1987. [ Links ]
11. Scott JA, Brogdon WG, Collins FH. Identification of single specimens of the Anopheles gambiae complex by the polymerase chain reaction. Am J Trop Med Hyg.1993;49:520-529. [ Links ]
12. Hunt RH, Brooke BD, Pillay C, Koekemoer LL, Coetzee M. Laboratory selection for and characteristics of pyrethroid resistance in the malaria vector Anopheles funestus. Med Vet Entomol. 2005;19:271-275. http://dx.doi.org/10.1111/j.1365-2915.2005.00574.x [ Links ]
13. World Health Organization (WHO). Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. Geneva: WHO; 2013. http://www.who.int/malaria/publications/atoz/9789241505154/en/ [ Links ]
14. Brooke BD, Kloke G, Hunt RH, Temu EA, Koekemoer LL, Taylor ME, et al. Bioassay and biochemical analyses of insecticide resistance in southern African Anopheles funestus. Bull Ent Res. 2001;91:265-272. http://dx.doi.org/10.1079/BER2001108 [ Links ]
15. Gericke A, Govere JM, Durrheim DN. Insecticide susceptibility in the South African malaria mosquito Anopheles arabiensis. S Afr J Sci. 2002;98:205-208. [ Links ]
16. Hargreaves K, Hunt RH, Brooke BD, Mthembu J, Weeto MM, Awolola TS, et al. Anopheles arabiensis and An. quadrianmiatus resistance to DDT in South Africa. Med Vet Entomol. 2003;17:417-422. http://dx.doi.org/10.1111/j.1365-2915.2003.00460.x [ Links ]
17. Munhenga G, Koekemoer LL. Differential expression of P450 genes in Anopheles arabiensis following permethrin selection. Afr J Biotechnol. 2011;10:12711-12716. [ Links ]
18. Nardini L, Christian RN, Coetzer N, Ranson H, Coetzee M, Koekemoer LL. Detoxification enzymes associated with insecticide resistance in laboratory strains of Anopheles arabiensis of different geographic origin. Parasit Vectors. 2012;5:113. http://dx.doi.org/10.1186/1756-3305-5-113 [ Links ]
19. Maharaj R, Mthembu DJ, Sharp BL. Impact of DDT re-introduction on malaria transmission in KwaZulu-Natal. S Afr Med J. 2005;95:871-874. [ Links ]
20. Maharaj R, Raman J, Morris N, Moonasar D, Durrheim DN, Seocharan I, et al. Epidemiology of malaria in South Africa: From control to elimination. S Afr Med J. 2013;103(10 Suppl 2):779-783. http://dx.doi.org/10.7196/samj.7441 [ Links ]
 Correspondence:
Correspondence:
Basil Brooke
Vector Control Reference Laboratory, National Institute for Communicable Diseases
Private Bag X4, Sandringham 2131
South Africa
Email: basilb@nicd.ac.za


{kind=link}
{kind=link}