










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.111 n.9-10 Pretoria Sep./Oct. 2015
http://dx.doi.org/10.17159/SAJS.2015/20140242
RESEARCH ARTICLE
Comparative morphometric analysis of the proximal femur of African hominids and felids
Andrew Gallagher
Centre for Anthropological Research (CfAR), University of Johannesburg, Johannesburg, South Africa
ABSTRACT
Size and shape of the mammalian proximal femur and taxon-specific distinctions in the relative proportions of the proximal articulation, the femoral neck and the proximal femoral diaphysis, are critical determinants in its adaptation to differential biomechanical stresses and observed locomotor habitus in different taxa. The morphometrics of the proximal femur are considered equally critical in the assessment of locomotor habitus of extinct fossil mammals, particularly extinct Miocene anthropoids and Plio-Pleistocene hominins. Analyses of size and shape of k=10 dimensions of the proximal femur were undertaken for a large sample series of two extant mammal families - the Felidae and Hominidae - using conventional multivariate statistical procedures, commonly used size-correction methods, and post-hoc tests of significance. While significant differences in form do exist, there are equally striking convergences in the functional morphology of extant hominid and felid taxa. Multivariate and bivariate allometric analyses confirm that the proximal femur of these two mammalian families share a common underlying structure manifest in a shared first common principal component. Nevertheless, while considerable convergences in general form of the proximal femur of African hominids and large-bodied felids are apparent, there exist equally discreet distinctions which are consistent with the differential structural demands imposed by their distinct locomotor and behavioural habitus.
Keywords: size and shape; proximal femoral morphology; biomechanics
Introduction
The proximal femur of mammals is extremely generalised in its external and internal form1-3, yet the unique mode of posture and progression in living humans and their extinct Plio-Pleistocene relations is manifest as a suite of consistent structural modifications4-6. The principles of osteonal remodelling, and that of its cartilaginous precursors, are universally accepted7-9 and the adult form of the proximal femur retains considerable potential as a source of quantitative evidence concerning taxonomy, function and phylogeny.4-6,10
All quadrupedal vertebrates are essentially hindlimb driven and, the unusual 'diagonal sequence' gait typical of primates notwithstanding, hindlimb drive is employed in recognition of the simple fact that the pelvic and proximal hindlimb musculature is the primary source of initial power input at commencement of the gait cycle from a static position of rest.11-14 The principal function of the musculoskeletal components of the hip joint are akin to the stroke cycle of a piston rod (or main rod) which powers the main driving wheels in the propulsion of steam-powered locomotives.15,16 Cyclical rotations and angular excursions of the spherical femoral head within its ball-and-socket joint and the shaft of the femur during contact phases of gait, particularly at optimal and peak velocities, are critical in maintaining momentum and direction of the centre of mass.14,17,18 From dynamic and static perspectives, the mammalian proximal femur differs from the main rod of a steam locomotive in that it must be 'engineered', for want of a more appropriate descriptor, to withstand potentially critical stresses induced by ground reaction forces which exceed body mass by a considerable magnitude at peak velocities.9,19
The femoral neck of mammals varies in its absolute (anatomical) length, its superoinferior angulation, and its degree of anteroposterior anteversion, yet despite observed intraspecific and interspecific functional variance in these parameters, it is generally accepted that the femoral neck performs the function of a cantilever subject to bending in the superioinferior plane.20-23 These stresses are greatest at the locus of the neck-shaft junction and are structurally mediated by the overall length of the cantilever (anatomical/functional length), its proportions in the primary plane of bending and in the orthogonal neutral axes (x,y) and, by extension, its geometric conformity.6,21,22,24
The primary function of the proximal femoral articulation in terrestrial vertebrates lies in the transfer of force, propulsive and reactive, engendered by mass of the organism during posture and ambulation and, to a greater degree, during high-energy ground contact at moderate-to-peak velocities of the gait cycle.12,19,25,26 The magnitude of these forces ultimately determines the absolute and relative size of the articular surface, which is greater in terrestrial bipeds.21,27 Absolute length of the neck is a critical determinant of the lever arm of the external and intrinsic hip musculature in quadrupeds and bipeds5,21 and in terrestrial hominins the neck-shaft junction is subject to considerably elevated compressive stresses compared with those of more arboreal species like Gorilla and Pan6,10. Proportions of the proximal and distal osseous components of the limbs and the digits (manus and pes) attest to considerable modifications to habitual structural demands, whether as levers increasing mechanical efficiency with respect to specific functional demands (i.e. suspension versus support; increased inertia in specialised high-velocity taxa), or in resistance to flexure and ultimate failure during peak loading cycles.13,28-30
The extant African hominids (Gorilla, Pan and Homo) and large-bodied felids (Panthera and Acinonyx) offer a unique prospectus to assess patterns of variance in the general morphology of the proximal femur and its modification to disparate structural demands. Both mammalian families exhibit considerable size variance that was likely exceeded in some extinct Plio-Pleistocene representatives31,32 and both include one highly specialised extant taxon, namely Homo and Acinonyx. However, there is good reason to designate the fore- and hindlimb anatomy of Acinonyx to being merely functionally derived, rather than unique, whereas the lower limb of Homo has no true extant comparator. Living humans were thus excluded from this analysis, which was focused upon the elucidation of distinctions and general underlying convergences in the proximal femoral morphology of other hominids and felids.
This contribution explicitly assesses patterns of underlying convergences and disparities in proximal femoral form of these two phyletically discreet families of extant African mammals. If the proximal femur of mammals is conservative in its general form, it follows logically that extant African hominids and large-bodied African felids will share a common generalised vector of size and shape (Ho 1). However, given the distinct postural repertoires and locomotor modes of extant African hominids and large-bodied felids, whereby the total mass of the individual is supported to a greater extent by the proximal femoral articulation in the former, an equally logical expectation is that any discreet morphometric distinctions of the proximal femur will be consistent with the mechanical demands of their observed disparate posture and locomotor modes.
Materials and methods
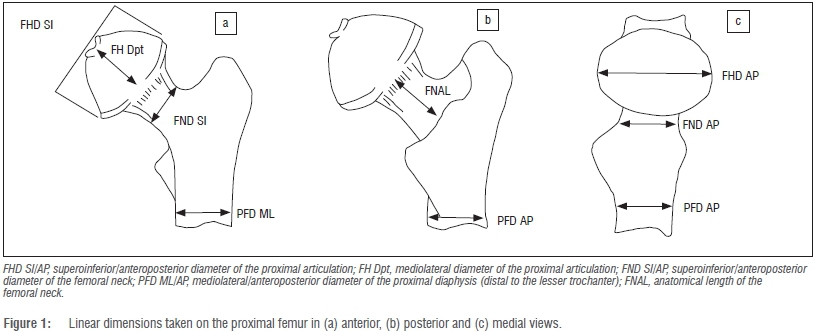
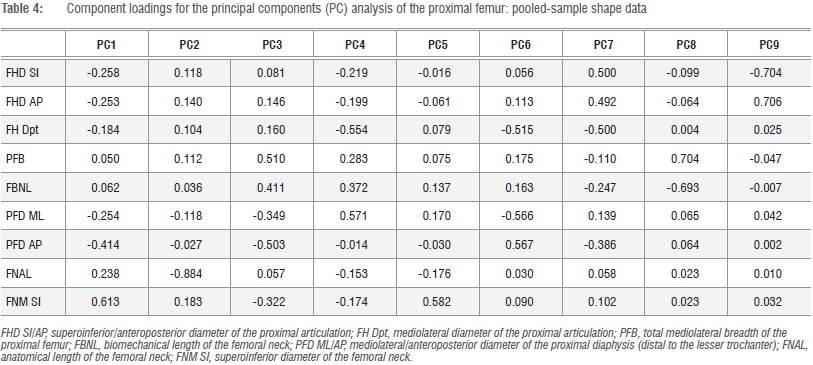
A total of 213 specimens of African hominid species and sub-species of the extant genera Gorilla (gorillas) and Pan (chimpanzees and bonobos) were included in the analysis (Supplementary table 1 of the online supplementary material). The extant large-bodied felid samples included Panthera - Panthera tigris (tigers), Panthera leo (lions) and Panthera pardus (leopards) - and Acinonyx (Acinonyx jubatus; cheetahs) and comprised 69 individuals. Only adult specimens evidencing complete fusion of the postcranial epiphyses were used in this study. A series of k=10 linear dimensions of the proximal femur used in conventional appraisals of the taxonomic and functional affinities of extinct Plio-Pleistocene hominin genera (Australopithecus, Paranthropus and early Homo)33were taken using a pair of Mitutoyo™ digital calipers (Figure 1). As can be seen, these linear dimensions effectively describe the general form (form=size+shape)34 of the proximal articulation (superoinferiorly, anteroposteriorly, mediolaterally), the proportions of the femoral neck and proximal femoral diaphysis (mediolaterally and anteroposteriorly).
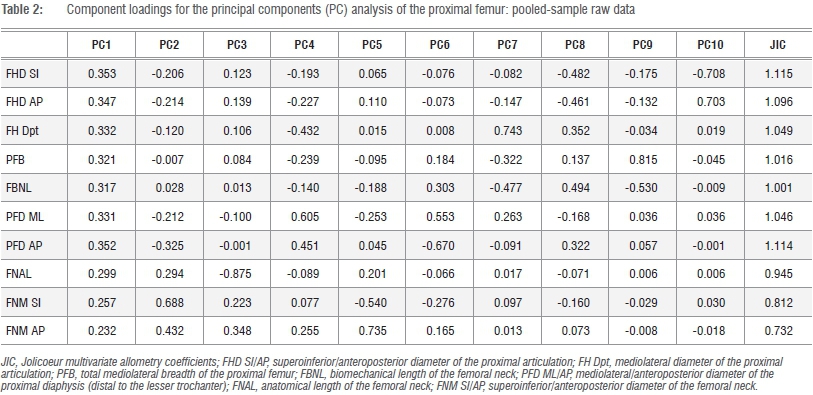
All linear data were log-transformed (base e) in MS Excel and subsequently size corrected using the approach outlined by Mosimann35, whereby each individual k linear dimension for a particular specimen is expressed as a proportion (not a percentage) of the geometric mean of all k=10 dimensions. The variates generated using this approach are effectively uncorrelated with size and facilitate both bivariate and multivariate perspectives on relative size distinctions between series of forms.35 Following the rationale of Jolicoeur, the most appropriate mathematical descriptor of size and shape is the first principal component (PC) of a log-transformed covariance matrix.35,36 If size is dominant, then the first PC (PC1) should account for an overwhelming proportion of the total observed variance, leaving little residual variance assigned to successive orthogonal (and uncorrelated) PC axes.37,38 If this prospectus holds, then the loadings of individual k variates on this axis can be expressed as multivariate allometry coefficients standardised around a value of 1, the theoretical coefficient of isometry.34-36 This is simply calculated as 1/sqrt(k)=0.316 in the case of our k=10 dimensions and all log-transformed raw variates should yield positive scores on the first PC.
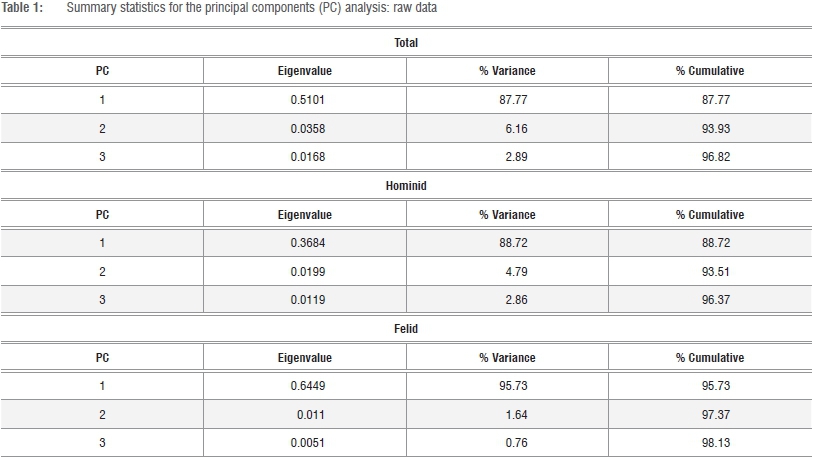
All principal components analyses (PCA) were performed on the variance-covariance matrix (hereafter VCV), as opposed to the correlation matrix, using PAST version 339, which extracts a series of components commensurate to the k-variate series,37,38 irrespective of the magnitude of the variance explained by successive PCs, that effectively diminish to a total cumulative 100%. If PC1 does constitute a 'generalised size and shape vector', then successive PCs may be effectively negligible.34-38 In the first instance, PCAs were performed on the log-transformed raw data of the total sample (pooled hominid and felid) and on the individual family series of hominids and large-bodied felids. Jolicoeur multivariate allometry coefficients were subsequently calculated for the component loadings of the k=10 linear variate series on PC1 alone.35,36
While the derivation of Jolicoeur multivariate allometry coefficients provides a computationally effective means of assessment of distinctions in n=2 (or n=>2) samples for a given battery of k linear dimensions, explicit tests of underlying structural commonalities in sample-specific VCV matrices were explored via the Flury hierarchy of common principal components analysis which assesses for equality, proportionality and sequential shared common PCs.40,41 The common principal components hierarchy is an effective test of underlying correspondences and disparities in matrix structure beyond a simple test of equality of n-group matrices. These tests were performed using the program written in MS Dos for Windows by Dr Patrick C. Phillips.
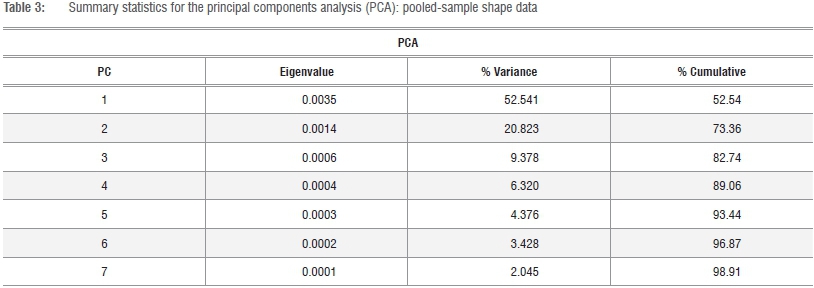
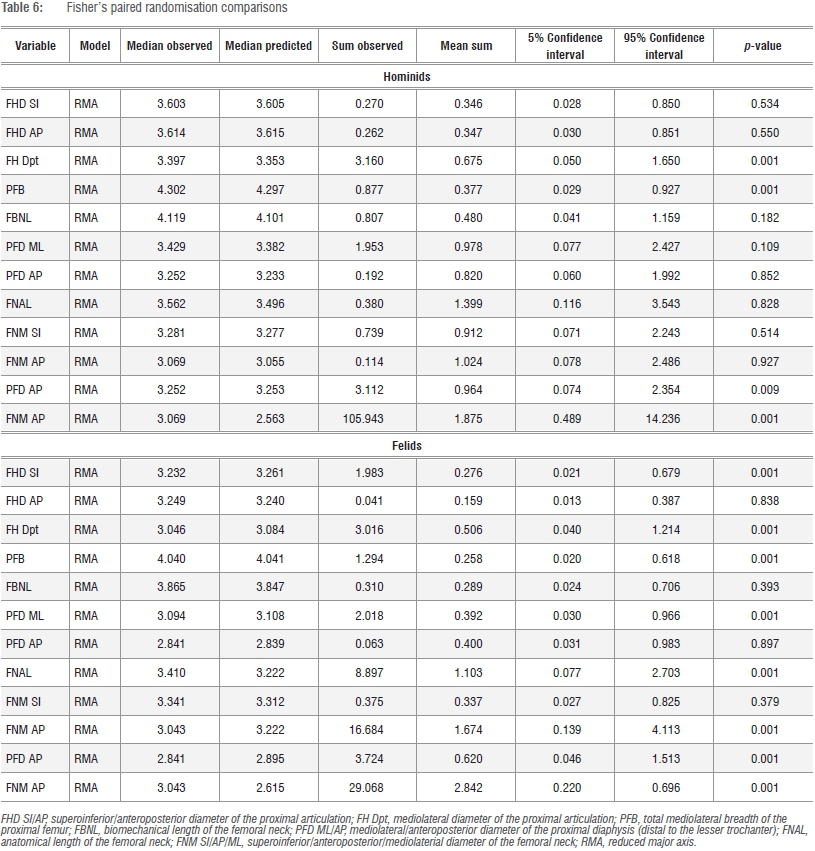
A second PCA was performed on the total series data set using the Mosimann shape variates of the k=10 dimensions in order to explicitly examine patterns of relative size disparities in the proximal femur of African hominids and large-bodied felids. Model II (reduced major axis) bivariate allometric equations were calculated in post-hoc assessments either as sample-specific (i.e. hominid or felid) or total-sample models using PAST v.3. Post-hoc significance tests of estimates of y (the dependent variate) on x (the geometric mean) were determined via Fisher's paired randomisation test performed in Rundom Projects 2.42,43
Results
Summary data for the PCAs of the total and individual sample series, PC loadings and multivariate allometry coefficients for the total sample series are given in Tables 1 and 2. The PC loadings and multivariate allometry coefficients for the individual series (African hominid, large-bodied felid) are given in the online supplementary material (Supplementary tables 2 and 3). In the three analyses of the log-transformed raw data, the first PC is a generalised size vector accounting for about 88% and 96% of the total variance and is greater in felids than in hominids (Table 1). Pearson's r between the PC1 scores and geometric means of the k=10 linear variates is perfect in both felids and hominids (r=1.000; Supplementary figure 1). Nevertheless, subtle distinctions do exist in the sample-specific multivariate allometry coefficients, particularly in dimensions reflecting absolute size of the proximal femur, proximal femoral diaphysis and femoral neck. Yet in both hominids and felids, the articulation is approximately isometric and is greater in the superoinferior plane (Supplementary tables 2 and 3).
As can be seen in Figure 2a, significant contrasts exist in the proximal femur of hominids and large-bodied felids on PC2, which primarily reflects differential loadings of the femoral head on the one hand and dimensions of the neck on the other. An interesting point of note is that the proximal femur of Pn. paniscus (bonobos) appears to be intermediate in form between extant felids and the main scatter of Pn. troglodytes (chimpanzees) and Gorilla on axis 2, reflecting correspondences in the general form of the femoral neck. The anatomical length, the superoinferior height and anteroposterior depth, load positively on this axis and are greater in felids than in hominids. An explicit assessment of commonality of underlying structure in the VCV matrices of log-transformed raw data using the Flury hierarchy confirms that both series share a first, but not a second, common PC. The VCV matrices are neither equivalent nor proportional, and significant contrasts in proximal femoral morphology in hominids and large-bodied felids are confirmed, irrespective of whether the 'jump-up' or 'step-up' approach is preferred (Supplementary tables 4 and 5). 41
The observed contrasts in the proximal femur of extant African hominids and large-bodied felids on the one hand, and Pn. paniscus on the other, are further exemplified in multivariate analyses of the Mosimann shape variates (Figure 3). As size has been effectively removed, five PCs are required to account for approximately 93% of the total variance and component loadings of the k=10 variates range from negative to positive (Tables 3 and 4; Figure 3). Clear distinctions exist between felids (+ scores) and hominids (- scores) on the first, but not the second, PC axis, which reflects contrasts in the relative proportions of the proximal articulation and proximal diaphysis, which are greater in Gorilla and Pan, and the femoral neck (Table 4). As in the PCA of the raw data series, the principle morphometric distinctions in the femoral neck of felids and hominids lie in the proportions of the femoral neck, particularly its relative area (superoinferior and anteroposterior diameters). The striking convergence in proximal femoral morphology of Pn. paniscus and large-bodied felids is equally apparent in Figure 3a, yet in Figure 3b the equally stark contrasts in absolute size of the head and total and functional length of the proximal femur are demonstrable (Table 4; Figure 3).
In order to further assess the significance of the observed distinctions in relative proportions of the proximal femur in African hominids and large-bodied felids, reduced major axis regression solutions were derived for either pooled-sample or family-specific models, depending upon expectations of a single or independent allometric trend. The geometric mean of the k=10 dimensions was used as the independent (x) variate in all comparisons with the exception of the proportions of the neck and proximal diaphysis, in which contrasts in relative anteroposterior/ mediolateral proportions were considered (Table 5). With few exceptions, the linear variates of the proximal femur attest to positive allometry in hominids and felids. Bivariate scatter plots of three variates that apparently contribute to effective discrimination of the proximal femur of hominids and large-bodied felids in multivariate space are shown in Figure 4. Only the absolute size of the proximal articulation exhibits a sound bivariate relationship with minimal scatter in the data and further attests consistent proportional distinctions between felids and hominids (Figure 4a). Although there is a general tendency for Pa. pardus and A. jubatus proximal femoral diaphyses to fall outside the ranges of Pan with respect to their femoral neck anteroposterior/mediolateral diaphyseal proportions in their lower size ranges, this tendency is less consistent in their upper ranges and scarcely holds for Pa. leo and Pa. tigris with Gorilla (Figure 4b,c). Despite its functional significance and consistency in the multivariate analyses, relative anatomical length of the neck exhibits inordinate variance. Data in Figure 4c confirm that the major difference in the proximal femur of Pn. paniscus compared with Pn. troglodytes and Gorilla lies in their proportionally shorter, rather than elongate, femoral neck.
A similar overlap is observed in a great majority of other variates regressed against the geometric mean (GM), yet there is some consistency in Pa. leo and Pa. tigris in the relative proportions of the proximal femoral diaphysis compared with gorillas of equivalent size (Supplementary figures 2-4). These data are generally consistent with post-hoc tests (Table 6) which confirm general distinctions in the relative size of the proximal articulation between hominids and felids and in the proportions of the femoral neck and proximal diaphysis. Nevertheless, comparisons of observed and expected series in felids (medians are presented as a guide) reveal subtle distinctions in the proximal femur of these taxa, irrespective of a general convergence in overall form (Tables 5 and 6; Figures 3 and 4; Supplementary figures 2-4).
Discussion and conclusions
Segmental mass distribution in primates differs from quadrupedal mammals primarily in a proportionally greater allocation of the hindlimb musculature.12,30,44,45 Nonetheless, even among living anthropoids, the terrestrial knuckle-walking gaits of Gorilla and Pan are unique in that the centre of mass is more distally located towards the hip joints than it is in terrestrial quadrupedal monkeys.12,13,30,32,45,46 More crucially, irrespective of differences in absolute size, the distribution of mass during forelimb contact in the gait cycle is only marginally higher in Gorilla than in Pan and does not exceed 20%.13 If large-bodied felids are accepted as suitable mammalian comparatives, then increased joint-reaction forces generated during normal posture and progression in extant African hominids is sufficient explanative for the observed proportional distinctions in their proximal femoral epiphyses.13,30,32,45,46 Despite the pooled-sample least-squares regression slope of greater than 1.00, the hip joints in intraspecific comparisons of Gorilla and Pan likely approximate isometry, and the observed proportional differences between African hominids and felids cannot be simply dismissed as an allometric consequence of extreme size in Gorilla.
Given their immense size and exponentially increased hip joint reaction forces, gorillas might be logically expected to exhibit proportionally greater proximal femoral articulations than chimpanzees, particularly in light of the fact that their femora are proportionally shorter relative to the length of the humerus.13,28,29,32,42,47 Humeral length increases isometrically with body mass (α=0.333) within the order Primates,47 so the elongate humerus of Gorilla is entirely expected for an anthropoid of its size. Following this reasoning, the reduced length of the femur, and not the elongation of the humerus, is imperative in their lower humero-femoral indices compared with Pan.13,28,29,32,47 Reduction in the length of the femur in Gorilla and their extreme midshaft diaphyseal cross-sectional areas29,32 compared with Pan, reflect necessary structural modifications to their increased bulk. Resultant increases in joint-reaction forces at the hip and knee of the lower limb in Gorilla and critical bending stresses at the femoral midshaft may be offset by a reduction in absolute and relative femur length, yet absolute length of the femur may independently mediate absolute size of the proximal and distal epiphyses in adults.29,32,33
The observed contrasts in proximal femoral morphology of Pn. paniscus and other extant African hominids in this analysis is deserving of consideration. The convergence in proximal femoral form of Pn. paniscus with the extant felids reflects subtle distinctions in the relative length and superoinferior/anteroposterior proportions of the femoral neck, which are greater than in Pn. troglodytes and Gorilla. On the one hand, these observed morphometric contrasts in the proximal femur of Pn. paniscus and Pn. troglodytes spp./Gorilla likely correspond with subtle, yet crucial, distinctions in extrinsic hip musculature architecture (M. Gluteus medius, M. Ischiofemoral) and moment arms (M. Gluteus medius) in bonobos and their emphasis in increased hip-flexion during climbing activities, which are more influential in the bonobo's lifestyle.48,49 Nevertheless, no significant contrasts in the general proportions of the neck of Pn. paniscus and Pn. troglodytes were found in previous geometric morphometric analyses50,51; the discrepancies between this analysis and that of Holliday and colleagues51 may simply reflect sampling constraints (larger samples in this study). However, divergences from the results of Harmon's 50 findings cannot be explained by such factors and require more detailed consideration.
Traditional morphometric methods differ both qualitatively and quantitatively from geometric morphometric approaches in that size standardisation exemplifies, rather than partitions, linear dimensions that may, or may not, approximate inter-landmark distances in 2D/3D registration space. In contrast, generalised Procrustes rotation (translation, rotation and scaling) does distort linear inter-landmark distances, which may potentially mask real morphometric distinctions in small-scale, localised regions of interest.52 Further to this, direct post-hoc comparisons of Asian and African apes in Harmon's50 study did reveal significant contrasts in the relative length of the femoral neck, which is proportionally greater in the former, yet no explicit post-hoc assessment of morphological distinctions in Pan was undertaken. At a basic level, these two methodologies are not directly comparable and any potential conflict with previously published analyses does not compromise the consistent contrasts witnessed in this study, nor their significance.
Despite substantial variation in body mass (from <4 kg to nearly 200 kg), extant felids do not exhibit large-scale, size-dependent changes in either limb posture or angular joint excursion during normal progression.53 Results of these analyses confirm that the proximal femoral morphology of living large-bodied felids exhibits striking consistencies in general form, which is particularly evident in multivariate appraisals of Mosimann shape variates with considerable overlap among taxa. This finding is somewhat unexpected, given the highly specialised gait of living cheetahs (A. jubatus) and only the relative mediolateral proportions of the proximal diaphysis attests taxon-specific distinctions when expressed as a function of the GM. Surprisingly, it is Pa. pardus, rather than Acinonyx, which differs from the morphology of Pa. leo and Pa. tigris by virtue of their relatively broad proximal shafts in the mediolateral plane. The most logical explanation for this finding is that the elongated anatomical length of the femoral neck in Pa. pardus increases joint excursion and yields greater mediolateral bending moments in the proximal femoral diaphysis during normal loading20-22 - a possible morphological function of increased tree-climbing behaviours observed in this species.54 Compared with thoroughbred greyhounds, cheetahs evidence considerable hypertrophy and increased momentarms of their hip-joint flexors and extensors.55-58
The hypertrophied hip flexors of Acinonyx, and presumably of other felid taxa, are critical in the initial power input to the stroke cycle of the proximal segment of the hindlimb during deceleration of the leading limb prior to foot contact, whereas the extensors of the hip and knee ('stifle') joints are equally critical immediately after initial ground contact and the spike in the vertical component of the ground reaction force as the entire limb is extended. The extrinsic muscles of the felid hip, particularly the gluteals, have a broad insertion across the posterior aspect of the proximal femoral diaphysis,1,2,55,56,58 whereas a principal intrinsic hip flexor, the M. Psoas muscle, is extremely well developed in Acinonyx compared with greyhounds. The M. Psoas muscle and the extrinsic hip-flexors are critical components of the accelerating leading limb during the areal phase of peak velocity galloping. Hypertrophy of the hip flexors and extensors associated with the proximal femoral diaphysis of felids, coupled with an elongate anatomical neck to increase their mechanical advantage,18,55-58 would effectively reduce, rather than increase, anteroposterior bending moments in the proximal shaft and femoral neck and would account for the distinctive 'pinching' compared with the morphology of Gorilla and Pan. Functional and structural factors thus mediate the limited morphological distinctions observable in the proximal femur of these two extant mammalian families.
While the biomechanical significance of the proximal femur and its associated musculature cannot be overstated, it is equally critical to remember that osseous structures, particularly the diarthroidal joints, are neither designed nor precision engineered15,16,21,24,26,27, they emerge via a complex process of axial cellular differentiation at the growth plate7-10,59. Recent advances have shed considerable light on the developmental emergence, postnatal growth and underpinning cellular signalling mechanisms that regulate the emerging growth plate and its complexities.7,9,10,59 Among living mammals, the emerging proximal femoral metaphysis comprises a singular basal growth plate, with discreet secondary ossification centres, either singular or dual, for the articulation and the greater trochanter.10,60Pan and Homo both share separate ossification centres for the articular head and greater trochanter, yet the cellular dynamics of the intervening proximal aspect of the femoral neck are subject to functionally distinct loading patterns which ultimately mediates absolute thickness of the cortex in this region.6,10 The role of androgens and oestrogens in the cellular dynamics of the proximal femur are also likely critical in the observed sex-specific variance in absolute and relative neck length in humans and other mammals and, by extension, the reduced mechanical integrity and susceptibility to fracture with advanced age.59,60 Results of this study suggest that the femoral neck of the living African apes is a worthy target for future research.
Results of this study confirm the prospectus that the proximal femur of two distinct families of mammals is extremely generalised in its external form; morphometric consistencies in extant African hominids and large-bodied felids considerably outweigh their observed distinctions. Further to this finding, these results underscore the necessity of a broader comparative context in understanding the nature of form and function in the mammalian postcranium. Nevertheless, where significant differences in proximal femoral form do occur - primarily in the relative size of the proximal articulation and in the anteroposterior/superoinferior and anteroposterior/mediolateral proportions of the neck and proximal diaphysis, respectively - these structural disparities are consistent with the mechanical requirements of their distinct locomotor modes and concomitant habitual stresses upon the proximal hindlimb. Further research into the morphology and functional significance of observed convergences and disparities in proximal femoral forms in other mammalian taxa can only increase our understanding of the underlying mechanisms and adaptive potential of this region of the vertebrate skeleton.
Acknowledgements
I would like to thank the following individuals and Institutions for their assistance and courtesy during the collection of data used in this study: Ms D. Hamerton (Iziko Museum of Natural History, Cape Town); Dr W. van Neer, Dr E. Gilissen and Mr W. Wendelin (Royal Museum of Central Africa, Tervuren); Dr Y Haile-Selassie, Dr B. Latimer and Mr L. Jellema (Cleveland Museum of Natural History, Cleveland); and Dr S. Badenhorst (Ditsong Museum of Natural History, Tshwane), and the various Collections Managers and the Trustees (Powell-Cotton Museum, Birchington) for access to the specimens in their care and for their generous hospitality. This research was funded by the Natural Environment Research Council (UK), The Ford Foundation (UK), The Palaeontology Scientific Trust (RSA), the Faculty of Health Sciences, University of the Witwatersrand (RSA), and the Faculty of Humanities, University of Johannesburg (RSA). I would also like to acknowledge Prof. Alan G. Morris and three anonymous reviewers for their helpful and thoughtful critique which improved this submission.
References
1. Wagner R. Elements of the comparative anatomy of the vertebrates. New York: JS Redfield; 1845. [ Links ]
2. Mivart G. St. The cat. An introduction to the study of backboned animals, especially mammals. New York: Schribner & Sons; 1881. [ Links ]
3. Reynolds SH. The vertebrate skeleton. Cambridge: Cambridge University Press; 1897. [ Links ]
4. Robinson JT. Early hominid posture and locomotion. Chicago, IL: Chicago University Press; 1972. [ Links ]
5. Lovejoy CO, Heiple KG, Burstein AH. The gait of Australopithecus. Am J Phys Anthropol. 1973;38:757-780. http://dx.doi.org/10.1002/ajpa.1330380315 [ Links ]
6. Ohman JC, Krochta TJ, Lovejoy CO, Mensforth RB, Latimer B. Cortical distribution of the femoral neck in hominoids: Implications for the locomotion of Australopithecus afarensis. Am J Phys Anthropol. 1997;104:117-131. http://dx.doi.org/10.1002/(SICI)1096-8644(199709)104:1<117::AID-AJPA8>3.0.CO;2-O [ Links ]
7. Murray PDF. Bones. Cambridge: Cambridge University Press; 1936. [ Links ]
8. Pearson OM, Lieberman DE. The aging of Wolff's 'law': Ontogeny and responses to mechanical loading in cortical bone. Ybk Phys Anthropol. 2004;47:63-99. http://dx.doi.org/10.1002/ajpa.20155 [ Links ]
9. Currey JD. Bones: Structure and mechanics. Princeton NJ: Princeton University Press; 2006. [ Links ]
10. Lovejoy CO, Meindl RS, Ohman JC, Heiple KG, White TD. The Maka femur and its bearing on the antiquity of human walking: Applying contemporary concepts of morphogenesis to the human fossil record. Am J Phys Anthropol. 2002;119:97-133. http://dx.doi.org/10.1002/ajpa.10111 [ Links ]
11. Hildebrand M. Symmetrical gaits of Primates. Am J Phys Anthropol. 1967;26:119-130. http://dx.doi.org/10.1002/ajpa.1330260203 [ Links ]
12. Demes B, Larson SG, Stern JT Jr, Jungers WL, Biknevicius A, Schmitt D. The kinetics of Primate quadrupedalism: 'Hindlimb drive' reconsidered. J Hum Evol. 1994;26:353-374. http://dx.doi.org/10.1006/jhev.1994.1023 [ Links ]
13. Preuschoft H. Mechanisms for the acquisition of habitual bipedality: Are there biomechanical reasons for the acquisition of upright bipedal posture? J Anat. 2004;204:363-384. http://dx.doi.org/10.1111/j.0021-8782.2004.00303.x [ Links ]
14. Schmitt D, Cartmill M, Griffin TM, Hanna JB, Lemelin P. Adaptive value of ambling gaits in Primates and other mammals. J Exp Biol. 2006;209:2042-2049. http://dx.doi.org/10.1242/jeb.02235 [ Links ]
15. Edwards E. Modern American locomotive engines: Their design, construction, and management. Philadelphia, PA: Carey Baird & Co; 1883 [ Links ]
16. Meyer JGA. Modern locomotive construction. New York: John Wiley & Sons; 1892. [ Links ]
17. Usherwood JR, Wilson AM. No force limit on greyhound sprint speed. Nature. 2005;438:753-754. http://dx.doi.org/10.1038/438753a [ Links ]
18. Hudson PE, Corr SA, Wilson AM. High-speed galloping in the cheetah (Acinonyx jubatus) and the racing greyhound (Canis familiaris): Spatiotemporal and kinetic characteristics. J Exp Biol. 2012;215:2425-2434. http://dx.doi.org/10.1242/jeb.066720 [ Links ]
19. Nigg BM, Herzog W. Biomechanics of the musculo-skeletal system. 2nd ed. New York: John Wiley & Sons; 1999. [ Links ]
20. Pauwels F. Biomechanics of the locomotor apparatus: Contributions to the functional anatomy of the locomotor apparatus. Berlin: Springer; 1980. [ Links ]
21. Ruff CB. Biomechanics of the hip and birth in early Homo. Am J Phys Anthropol. 1995;98:527-574. http://dx.doi.org/10.1002/ajpa.1330980412 [ Links ]
22. Rafferty K. Structural design of the femoral neck in Primates. J Hum Evol. 1998;34:361-383. http://dx.doi.org/10.1006/jhev.1997.0202 [ Links ]
23. Beer FP Johnston ER, DeWolf JT, Mazurek DF. Mechanics of materials. 6th ed. New York: McGraw Hill; 2012. [ Links ]
24. Lovejoy CO, Burstein AH, Heiple KG. The biomechanical analysis of bone strength: A method and its application to platycnemia. Am J Phys Anthropol. 1976;44:489-506. http://dx.doi.org/10.1002/ajpa.1330440312 [ Links ]
25. Paul JP. Forces transmitted by joints in the human body. Proc Inst Mech Engin. 1967;181:8-15. [ Links ]
26. Paul JP. Approaches to design: Force action transmitted by joints in the human body. Proc R Soc Lond B-Bio 1976;192:163-172. http://dx.doi.org/10.1098/rspb.1976.0004 [ Links ]
27. Jungers WL. Relative joint size and hominoid locomotor adaptations with implications for the evolution of hominid bipedalism. J Hum Evol. 1988;17:247-265. http://dx.doi.org/10.1016/0047-2484(88)90056-5 [ Links ]
28. Schultz AH. The skeleton of the trunk and limbs of higher primates. Hum Biol. 1930;3:303-321. [ Links ]
29. Schultz AH. Proportions, variability, and asymmetries of the long bones of the limbs and the clavicles in man and apes. Hum Biol. 1930;9:281-328. [ Links ]
30. Demes B, Günther MM. Biomechanics and allometric scaling in Primate locomotion and morphology. Folia Primatol. 1989;53:125-141. http://dx.doi.org/10.1159/000156412 [ Links ]
31. Eisenberg JF. The mammalian radiations: An analysis of trends in evolution, adaptation, and behaviour. Chicago, IL: Chicago University Press; 1990. [ Links ]
32. Fleagle JG. Primate adaptation and evolution. 2nd ed. London: Academic Press; 1999. [ Links ]
33. McHenry HM, Corruccini RS. The femur in early human evolution. Am J Phys Anthropol. 1978;49;473-488. http://dx.doi.org/10.1002/ajpa.1330490407 [ Links ]
34. Gould SJ. Allometry and size in ontogeny and phylogeny. Biol Rev. 1966;41:587-640. http://dx.doi.org/10.1111/j.1469-185X.1966.tb01624.x [ Links ]
35. Mosimann JE. Size allometry: Size and shape variables with characterisations of the lognormal and gamma distributions. J Am Statist Ass. 1970;65:930-945. http://dx.doi.org/10.1080/01621459.1970.10481136 [ Links ]
36. Jolicoeur PF. The multivariate generalisation of the allometry equation. Biometrics. 1963;19:497-499. http://dx.doi.org/10.2307/2527939 [ Links ]
37. Blackith RE, Reyment RA. Multivariate morphometrics. New York: Academic Press; 1970. [ Links ]
38. Joliffe IT. Principal components analysis. 2nd ed. New York: Springer; 2002. [ Links ]
39. Hammer 0, Ryan PD, Harper DAT. PAST: Palaeontological statistics software package for education and data analysis. Palaeont Electr. 2001;4(1), 9 pages. Available from: http://folk.uio.no/ohammer/past [ Links ]
40. Flury BK. Common principal components and related multivariate procedures. New York: Wiley; 1988 [ Links ]
41. Phillips PC, Arnold SJ. Hierarchical comparison of genetic variance-covariance matrices. I. Using the Flury hierarchy. Evolution. 1999;53:506-1515. http://dx.doi.org/10.2307/2640896 [ Links ]
42. Fisher RA. The design of experiments. 8th ed. Edinburgh: Oliver and Boyd; 1966. [ Links ]
43. Jadwisczcak FP Rundom Projects 2; 2003. Available from: http://pjadw.tripod.com [ Links ]
44. Isler K, Payne RC, Günther MM, Thorpe SKS, Yu L, Savage R, et al. Inertial properties of hominoid limb segments. J Anat. 2006;209:201-218. http://dx.doi.org/10.1111/j.1469-7580.2006.00588.x [ Links ]
45. Raichlen DA, Pontzer H, Shapiro LJ, Sockol MD. Understanding hindlimb weight support in chimpanzees with implications for the evolution of Primate locomotion. Am J Phys Anthropol. 2009;138:395-402. http://dx.doi.org/10.1002/ajpa.20952 [ Links ]
46. Crompton RH, Vereecke EE, Thorpe SKS. Locomotion and posture from the common hominoid ancestor to fully modern hominins, with special reference to the last common panin/hominin ancestor. J Anat. 2008;212:501-543. http://dx.doi.org/10.1111/j.1469-7580.2008.00870.x [ Links ]
47. Jungers WL. Aspects of size and scaling in Primate biology with special reference to the locomotor skeleton. Am J Phys Anthropol. 1984;27:73-97. http://dx.doi.org/10.1002/ajpa.1330270505 [ Links ]
48. Payne RC, Crompton RH, Isler K, Savage R, Vereecke EE, Günther MM, et al. Morphological analysis of the hindlimb in apes and humans. I. Muscle architecture. J Anat. 2006;208:709-724. [ Links ]
49. Payne RC, Crompton RH, Isler K, Savage R, Vereecke EE, Günther MM, et al. Morphological analysis of the hindlimb in apes and humans. II. Moment arms. J Anat. 2006;208:725-742. http://dx.doi.org/10.1111/j.1469-7580.2006.00564.x [ Links ]
50. Harmon EH. The shape of the hominoid proximal femur: A geometric morphometic analysis. J Anat. 2007;210:170-175. http://dx.doi.org/10.1111/j.1469-7580.2006.00688.x [ Links ]
51. Holliday TW, Hutchinson VT, Morrow MB, Livesay GE. Geometric morphometric analyses of hominid proximal femora: Taxonomic and phylogenetic considerations. Homo - J Comp Hum Biol. 2010;61:3-15. http://dx.doi.org/10.1016/j.jchb.2010.01.001 [ Links ]
52. Von Cramon-Taubadel N, Frazier BC, Lahr MM. The problem of assessing landmark error in geometric morphometrics: Theory, methods, and modifications. Am J Phys Anthropol. 2007;134:24-35. http://dx.doi.org/10.1002/ajpa.20616 [ Links ]
53. Day LM, Jayne BC. Interspecific scaling of the morphology and posture of the limbs during the locomotion of cats (Felidae). J Exp Biol. 2007;210:642-654. http://dx.doi.org/10.1242/jeb.02703 [ Links ]
54. Brain CK. The hunters or the hunted? Chicago, IL: Chicago University Press; 1981. [ Links ]
55. Hildebrand M, Goslow GC. Analysis of vertebrate structure. New York: John Wiley and Sons; 2002. [ Links ]
56. Kardong KV. Vertebrates: Comparative anatomy, function, and evolution. 6th ed. New York: McGraw Hill; 2009. [ Links ]
57. Williams SB, Tan H, Usherwood JR, Wilson AM. Pitch then power: Limitations to acceleration in quadrupeds. Biol Lett. 2009;5:610-613. http://dx.doi.org/10.1098/rsbl.2009.0360 [ Links ]
58. Hudson PE, Corr SA, Payne-Davis RA, Clancy SN, Lane E, Wilson AM. Functional anatomy of the cheetah hindlimb. J Anat. 2011;218:363-374. http://dx.doi.org/10.1111/j.1469-7580.2010.01310.x [ Links ]
59. Vanderschueren D, Vandenput L, Boonen S, Lindberg SK, Bouillon R, Ohlsson C. Androgens and bone. End Rev. 2004;25:389-425. http://dx.doi.org/10.1210/er.2003-0003 [ Links ]
60. Serrat MA, Reno PL, McCollum MA, Meindl RS, Lovejoy CO. Variation in mammalian proximal femoral development: Comparative analysis of two distinct ossification patterns. J Anat. 2007;210:249-258. http://dx.doi.org/10.1111/j.1469-7580.2007.00694.x [ Links ]
 Correspondence:
Correspondence:
Andrew Gallagher
Centre for Anthropological
Research, Department of
Anthropology and Development
Studies, University of Johannesburg
Auckland Park 2006, Johannesburg
South Africa
Email: agal1815@gmail.com
Received: 18 July 2014
Revised: 30 Oct. 2014
Accepted: 04 Jan. 2015
Note: This article is supplemented with online only material.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}