




![Dichloro(bis[diphenylthiourea])cadmium complex as a precursor for HDA-capped CdS nanoparticles and their solubility in water](/img/en/next.gif)





Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.111 n.7-8 Pretoria Jul./Aug. 2015
http://dx.doi.org/10.17159/SAJS.2015/20140369
REVIEW ARTICLE
Pancreatic islet regeneration: Therapeutic potential, unknowns and controversy
Ingrid L. Cockburn; William F. Ferris
Division of Endocrinology, Department of Medicine, Faculty of Medicine and Health Sciences, Stellenbosch University, Tygerberg, South Africa
ABSTRACT
Glucose homeostasis in mammals is primarily maintained by the insulin-secreting β-cells contained within pancreas-resident islets of Langerhans. Gross disruption of this glucose regulation as a result of pancreatic dysfunction frequently results in diabetes, which is currently a major health concern in South Africa, as well as globally. For many years, researchers have realised that the pancreas, and specifically the islets of Langerhans, have a regenerative capacity, as islet mass has frequently been shown to increase following induced pancreatic injury. Given that gross β-cell loss contributes significantly to the pathogenesis of both type 1 and type 2 diabetes, endogenous pancreatic islet regeneration has been investigated extensively as a potential β-cell replacement therapy for diabetes. From the extensive research conducted on pancreatic regeneration, opposing findings and opinions have arisen as to how, and more recently even if, pancreatic regeneration occurs following induced injury. In this review, we outline and discuss the three primary mechanisms by which pancreatic regeneration is proposed to occur: neogenesis, β-cell replication and transdifferentiation. We further explain some of the advanced techniques used in pancreatic regeneration research, and conclude that despite the technologically advanced research tools available to researchers today, the mechanisms governing pancreatic regeneration may remain elusive until more powerful techniques are developed to allow for real-time, live-cell assessment of morphology and gene expression within the pancreas.
Keywords: diabetes; islets of Langerhans; β-cells; neogenesis; transdifferentiation
Diabetes: Therapies and challenges
The prevalence of diabetes and its comorbidities, such as cardiovascular disease, is increasing rapidly both globally and in South Africa.1 It is currently estimated that 347 million people worldwide suffer from the disease and the prevalence of diabetes is predicted to double between 2005 and 2030.2 Indeed, in a recent comprehensive survey on health and nutrition in South Africa, diabetes was diagnosed in 9.6% of the survey participants (aged >l5 years),3 which, based on South Africa's current population, equates to ~5 million people living with the disease. Of particular concern is that the prevalence in some demographic groups far exceeds the national average: diabetes was diagnosed in as much as 30.7% of the Asian / Indian study participants.3 This malady exerts a considerable burden of disease, which will increase with its rapidly escalating prevalence.
Diabetes is commonly subdivided into two types, with type 1 diabetes (T1D) believed to account for only 5-10% of all diabetes cases4, although little data are currently available regarding the prevalence of T1D in South Africa5. The insulin deficiency associated with T1D is caused by an autoimmune destruction of insulin-producing pancreatic β-cells.6 The hyperglycaemia and ketosis resulting from gross β-cell depletion in T1D patients are currently treated with insulin replacement,7 and research into potential T1D therapeutics is therefore commonly focused on developing strategies to eliminate the dependence of patients on exogenous insulin. One such approach, which represents a major advancement in diabetes therapy, is islet or pancreas transplantation. Transplantation of either whole pancreata or isolated islets as a means to regain pancreatic endocrine function has been successfully used to reverse T1D; however, limitations - including an insufficient number of donor organs, poor cell viability and undesirable effects of the accompanying immunosuppressive drugs - have meant that transplantation is currently not a feasible and sustainable solution.8,9 Although type 2 diabetes (T2D) is generally characterised by hyperglycaemia primarily resulting from insulin resistance, insufficient insulin production as a result of the loss of β-cells, as in T1D, is also important in the aetiology of the disease10, as β-cell function and mass are known to be reduced in T2D patients11.
To address the limitations associated with the management of diabetes, such as the exogenous insulin-dependence of patients as well as the shortcomings of islet or pancreas transplantation, possible alternative sources of β-cells for replacement therapy have been investigated. These alternatives include the stimulation of the pancreas to promote the endogenous regeneration of viable β-cells. With a view to future pharmacological or cell therapy interventions, researchers have looked to in-utero pancreatic organogenesis to ascertain which molecular pathways may be important for generating increased islet mass. In particular, the temporal expression of transcription factors involved in pancreatic cell fate determination has been investigated and characterised.
In-utero pancreatic development
Knowledge of the in-utero development of the mammalian pancreas, and in particular of the origin and development of the islets of Langerhans and β-cells, provides a starting point for investigations into potential regenerative processes in the adult pancreas. All exocrine and endocrine cell types of the pancreas originate from a common pool of progenitor cells in the gut endoderm of the embryo. A pancreatic bud forms from the endoderm, and subsequently expands and forms branched structures which eventually form the pancreatic ducts.12 During the extension of these branches, clusters of endocrine cells bud off and aggregate to form the islets of Langerhans.13 A brief overview of the in-utero development of islet cells and the transcription factors involved in this process is depicted in Figure 1.

The transcription factor Ipf1 (insulin promoter factor 1, Pdx1 in rodents) is known to play a major role in pancreatic specification and growth in early embryonic development, and indeed, lineage-tracing experiments have shown that all pancreatic cells, both exocrine and endocrine, arise from Ipf1-expressing progenitor cells.14 Further along in pancreatic development, the endocrine portion of the pancreas (islets of Langerhans) is formed from a subset of pancreatic duct endothelial cells expressing a second transcription factor central to islet development, namely neurogenin 3 (Ngn3): Ngn3-expressing progenitors differentiate into the five endocrine cell types (α-, β-, δ-, ε- and PP-cells), which subsequently separate from the endothelium and cluster to form the islets of Langerhans.15,16 Of the five islet cell types, α- and β-cells are the most abundant17; and while murine islets consist of a β-cell core surrounded by a mantle of α- and δ-cells, the α-, β- and δ-cells are dispersed throughout human and non-human primate islets18.
The development of β-cells, in particular from Ngn3+ cells, is regulated by the Pax4 gene19, a member of the homeobox transcription factor gene family which comprises a large and diverse group of genes that play an important role in embryonic development20. Pax4 expression is known to peak during in-utero β-cell development21, and, in a knockout study, Pax4-deficient mice showed significantly diminished β-cell development19. The absence of Pax4 expression in mature islet cells is indicative of the importance of Pax4 expression to β-cell development, in particular, rather than β-cell function or maintenance.21
Pancreatic regeneration
Discovery and models
Since the 1960s, many reports have demonstrated in-vivo manipulation of the pancreas by mechanical stress (injury) to stimulate the regeneration of damaged tissue and, importantly, also an increase in islet mass in the pancreas. Ligation or partial occlusion of the main pancreatic duct22-25 as well as incomplete pancreatectomy26,27 have been shown to result in pancreatic regeneration - observed as increased mass of the endocrine portion (islets) of pancreata following induced injury. In the 1970 study by Boquist and Edstrom22, ligation of the main pancreatic duct in rats resulted in degeneration of acinar cells, with no signs of regeneration of this exocrine moiety of the pancreas. There was, however, an increase in endocrine cells following duct ligation, specifically by endocrine clusters seen to bud off from proliferating ductules and develop into islets. Similarly, Rosenberg and colleagues23 observed the formation of new islets from hyperplasic (proliferating) ductules after partial occlusion of the pancreatic ducts of hamsters by wrapping a thin cellophane strip around the head of the pancreas. In more significant pancreatic injury (90% pancreatectomy in rats), pronounced regeneration of both the endocrine and exocrine pancreatic tissues was observed 8 weeks subsequent to surgeries.26 All these studies involved subjecting the pancreata of study animals to prolonged mechanical stress; however, it was later shown that even brief occlusion of the main pancreatic duct (by gently squeezing the pancreas for 60 s) in rats results in duct cell proliferation and signs of islet regeneration24, as well as overall increased endocrine mass (by 80%) 56 days after surgery, suggestive of islet neogenesis25. The fact that apparent islet neogenesis was observed even after only very brief occlusion of the pancreatic duct suggested that the signalling events triggering these regenerative processes in response to pancreatic injury occur immediately at the initiation of injury.
Proposed mechanisms of regeneration
The observations described above lead into key and currently unresolved questions regarding the mechanisms and processes governing islet regeneration. Islet regeneration, and specifically regeneration of β-cells following pancreatic injury, has generally been attributed to one of three mechanisms: transdifferentiation of non-endocrine cells into β-cells, neogenesis of β-cells from progenitor cells, or replication of existing β-cells (Figure 2); however, contradictory opinions and data surrounding these theories are plentiful.
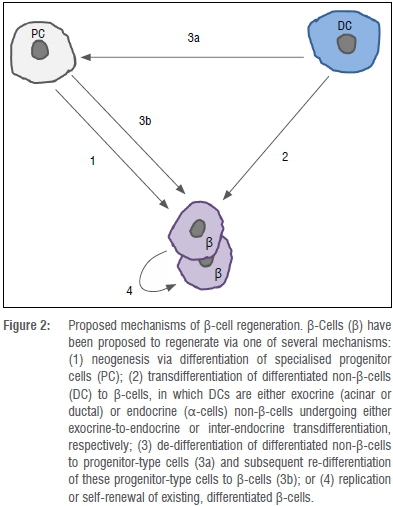
Transdifferentiation
As islet cells originate from a subset of duct cells during pancreatic organogenesis (as described above), transdifferentiation has been investigated as a possible mechanism by which islet regeneration occurs in the adult pancreas. In adult rats subjected to pancreatic duct ligation, transdifferentiation has been identified in the pancreas in the form of cells co-expressing markers from more than one terminally differentiated cell type, indicating a mixed lineage. These cells include those expressing both epithelial and β-cell markers; cells co-expressing epithelial and α-cell markers; duct cells expressing GLUT-2, the β-cell-specific glucose transporter protein28; or intra- and extra-islet cells co-expressing acinar cell and β-cell markers29. In these and other studies,30-32 increased islet cell number following pancreatic injury was thus attributed mainly to the transdifferentiation of non-endocrine (acinar or duct) cells to endocrine (α- or β-) cells. In an in-vitro study, AR42J acinar-derived amylase-secreting cells were converted to insulin-secreting cells by treatment with the growth factors betacellulin and activin A,33 and although insulin secretion by these cells does not confirm their identity as β-cells, the observed conversion is suggestive of exocrine-to-endocrine transdifferentiation. In a more recent lineage-tracing study on transgenic mice subjected to dipthera toxin-induced β-cell ablation, inter-endocrine transdifferentiation was also suggested to occur: new β-cells were identified as arising from α-cells.34
Desai and colleagues35 refuted the hypothesis that exocrine acinar cells transdifferentiate into endocrine β-cells based on the findings of their in vivo lineage-tracing experiments in which the acinar cells of transgenic mice were genetically labelled. This labelling strategy allowed for the progeny of these cells to be identified, thereby enabling the investigators to identify cells of acinar origin. Various models of pancreatic injury were used to induce regeneration, after which a lack of labelled endocrine cells was observed, leading the authors to conclude that 'acinar cells do not normally transdifferentiate into islet beta cells in vivo in adult mice'35. The findings of another genetic labelling-based study further countered the notion of transdifferentiation being the mechanism by which islet regeneration occurs by indicating that β-cells only arise from duct epithelial cells during embryogenesis, and that these epithelial cells do not significantly contribute to endocrine or acinar cell populations after birth.36
The findings described here clearly demonstrate that reports regarding transdifferentiation as a mechanism by which pancreatic regeneration occurs are as contradictory as they are plentiful.
Neogenesis from specialised progenitors
A population of specialised progenitor cells in the pancreas that can give rise to new β-cells would be extremely valuable for the development of endogenous cell therapies for the treatment of diabetes-associated β-cell depletion. Such a cell population could conceivably either be isolated, expanded ex vivo and used for transplantation or alternatively be stimulated to produce new β-cells in vivo. Thus far, however, whether or not such a specialised progenitor cell population exists in the pancreas remains unclear despite a number of reports in support of the existence of such cells. In corroboration with the existence of progenitor cells within the pancreas, flow cytometry has been used to identify a side population of cells in the murine pancreas which has been described as a putative stem cell population. These cells, identified as stem cells based on their ability to expel the Hoechst 33342 DNA-binding dye, were shown to undergo hyperplasia in vivo after β-cell or pancreas injury and, upon induction, the cells proliferated and differentiated in vitro giving rise to endocrine cells that exhibited glucose-stimulated insulin secretion.37 These findings led the authors to conclude that progenitor cells that give rise to endocrine cells exist within the pancreas, a conclusion also reached by other researchers who identified endogenous β-cell progenitors on the basis of the expression of the islet cell-specific transcription factor Ngn3: Ngn3-expressing cells located in the ductal lining of the pancreas were shown to be multi-potent progenitor cells with the ability to give rise to new glucose-responsive β-cells both in situ and in vitro.38 Very small embryonic-like stem cells (VSELs), a novel type of pluripotent stem cell reported to exist in various adult murine organs including the pancreas,39 were recently reported to mobilise to the pancreas following partial pancreatectomy in mice. These VSELs reportedly differentiated into progenitor cells expressing Pdx1 (the transcription factor expressed by progenitor cells that differentiate into all pancreatic cell types during organogenesis), potentially giving rise to new acinar and islet cells.40
Based on the findings of another study using a partial pancreatectomy model in rats, islet regeneration has been suggested to occur via initial de-differentiation of duct cells to progenitor-type cells, followed by re-differentiation of these cells, which 'recapitulate aspects of embryonic pancreas differentiation' to facilitate pancreatic regeneration41 - a mechanism which can be classed as neogenesis or transdifferentiation, or indeed a combination of the two.
A report opposing the hypothesis that β-cell regeneration following pancreatic injury occurs via neogenesis was recently published: the results of lineage-tracing experiments carried out using various models of β-cell loss in the adult murine pancreas led the authors to conclude that little to no β-cell neogenesis occurs in the adult pancreas under normal and pathological (partial pancreatectomy, duct ligation or treatment with β-cell-specific toxins) conditions.42
β-cell replication
The controversies and conflicting opinions within the field of islet regeneration are clearly demonstrated by the literature on β-cell replication as the mechanism driving β-cell maintenance and/or regeneration: 'Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation'43; 'β-cell replication is the primary mechanism subserving the postnatal expansion of β-cell mass in humans'44 and 'β-cell growth and regeneration: Replication is only part of the story'45 are the titles of just some of these publications.
It was initially believed that significant β-cell proliferation does not occur after the initial periods of β-cell mass growth during the neonatal and infancy stages of development, and that β-cell mass is not replenished or maintained by β-cell proliferation.46-48 Studies on β-cell replication have thus generally been carried out to assess not only β-cell replenishment following pancreatic injury, but also increases and maintenance of β-cell mass under normal physiological conditions. Similar to the lineage-tracing experiments carried out by Desai and colleagues35 (described previously), Dor et al.43 made use of a transgenic mouse model and a tamoxifen-inducible Cre/lox system to integrate genes into the genome by which β-cells were specifically labelled with a histochemically detectable, heritable label. Subsequent to removal of the initial 'pulse' (tamoxifen treatment), only pre-labelled β-cells and progeny of such cells would carry the detectable label, allowing newly formed β-cells to be identified as being β-cell-derived or non-β-cell-derived. These genetic labelling experiments led the authors to conclude that, during normal adult life or following pancreatectomy in the murine pancreas, adult β-cells are formed or replenished by replication rather than by islet neogenesis or differentiation of stem cells.43 In agreement with the conclusion reached by Dor and colleagues, a later human study in which β-cell mass was assessed using computer tomography techniques revealed that β-cell mass expansion during infancy, the period during which β-cell growth rates were found to be highest, occurs primarily via β-cell replication44; and in a transgenic mouse model, near-complete β-cell ablation was followed by full recovery of the pancreas, which was described to occur via β-cell replication within existing islets rather than islet neogenesis49. In an extensive review by Bonner-Weir and colleagues45, β-cell replication and neogenesis as mechanisms of pancreatic regeneration or postnatal islet growth are described as not being mutually exclusive. The authors go on to review the many reports on pancreatic regeneration and conclude that both neogenesis and β-cell replication contribute to β-cell mass maintenance, and that both these mechanisms have the potential to be harnessed for therapeutic applications.45
Doubt shed on regeneration
Controversy surrounding β-cell regeneration has recently extended from differing opinions on the mechanisms allowing for regeneration, to doubts being raised as to whether regeneration following pancreatic injury does in fact occur at all: Rankin et al.50 recently reported on extensive experiments carried out in an adult mouse model which, in agreement with the report by Xiao and colleagues42, show that adult β-cells do not develop from specialised pancreas-resident progenitor cells following pancreatic injury and, importantly, also that new β-cells are not generated following pancreatic duct ligation.50 The authors of this report attribute the apparent regeneration that has frequently been described for similar pancreatic injury models to quantitative artefacts and variable recovery of pancreatic tissue; an extensive morphometric assessment of the entire murine pancreas in Rankin et al.'s study indicates that β-cell mass is unaltered in the ligated pancreas compared with sham-operated tissue and therefore that injury of the adult murine pancreas does not induce β-cell regeneration.50
Both Xiao et al.42 and Rankin et al.50 conclude that β-cell neogenesis does not occur following pancreatic injury; however, in both these studies, the expression of the islet-specific transcription factor Ngn3 was shown to be increased following pancreatic injury. Kushner and colleagues attributed the observed induction of Ngn3 expression by ligation in their study to an 'artefact due to differences in RNA recovery from injured compared with uninjured pancreas'50, while damage to the injured portion of the pancreas (specifically exocrine cell contents and various inflammatory factors) was proposed to induce an up-regulation of Ngn3 expression in existing β-cells by Xiao and colleagues42.
Perspective
In this brief review we have presented the diverse and often contradictory findings of some of the many investigations into pancreatic regeneration carried out over the last five decades. It is clear from these studies that putative islet regeneration is a complex, poorly understood and controversial research area, but the potential benefits of understanding and possibly harnessing the processes involved are immense. Despite recent reports contesting the existence of β-cell regeneration, a lot of unknowns in this field remain to be clarified: Why do different studies obtain vastly different results when investigating the same models and systems? If pancreatic regeneration does not take place, why is Ngn3 up-regulated following pancreatic injury?
Although sophisticated techniques available to us today, such as genetic lineage-tracing technology, are extremely powerful, they too are limited. In the case of investigations into pancreatic injury-induced events, the ideal scenario would be one in which cells can be monitored in real time within the pancreas, simultaneously assessing both morphological changes and gene expression. Although three-dimensional microscopy and the culturing of whole or partial fragments of tissues have advanced considerably in recent years, this ideal is still beyond our capabilities. Until such time that technological advances will allow for such assessments to be carried out, definitive mechanisms that potentially stimulate an increase in β-cell mass triggered by pancreatic injury remain elusive.
Acknowledgements
We thank the Harry Crossley Foundation for funding our research on pancreatic islet neogenesis.
Authors' contributions
I.C. researched and wrote the review; W.F. was involved in planning the review outline and edited drafts of the manuscript.
References
1. Danaei G, Finucane MM, Lu Y Singh GM, Cowan MJ, Paciorek CJ, et al. National, regional, and global trends in fasting plasma glucose and diabetes prevalence since 1980: Systematic analysis of health examination surveys and epidemiological studies with 370 country-years and 2.7 million participants. Lancet. 2011;378(9785):31-40. http://dx.doi.org/10.1016/S0140-6736(11)60679-X [ Links ]
2. World Health Organization. Diabetes programme [homepage on the Internet]. No date [cited 2014 Oct 21]. [ Links ] Available from: http://www.who.int/diabetes/en/
3. Shisana O, Labadarios D, Rehle T, Simbayi L, Zuma K, Dhansay A, et al. South African National Health and Nutrition Examination Survey (SANHANES-1). Cape Town: HSRC Press; 2013. [ Links ]
4. American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care. 2009;32(Suppl 1):S62-S67. [ Links ]
5. Mbanya JCN, Motala AA, Sobngwi E, Assah FK, Enoru ST. Diabetes in sub-Saharan Africa. Lancet. 2010;375(9733):2254-2266. http://dx.doi.org/10.1016/S0140-6736(10)60550-8 [ Links ]
6. Atkinson MA, Maclaren NK. The pathogenesis of insulin-dependent diabetes mellitus. N Engl J Med. 1994;331(21):1428-1436. http://dx.doi.org/10.1056/NEJM199411243312107 [ Links ]
7. Van Belle TL, Coppieters KT, Von Herrath MG. Type 1 diabetes: Etiology, immunology, and therapeutic strategies. Physiol Rev. 2011;91(1):79-118. http://dx.doi.org/10.1152/physrev.00003.2010 [ Links ]
8. Noguchi H. Stem cells for the treatment of diabetes. Endocr J. 2007;54(1):7-16. http://dx.doi.org/10.1507/endocrj.KR-86 [ Links ]
9. Haller MJ, Viener H-L, Wasserfall C, Brusko T, Atkinson MA, Schatz DA. Autologous umbilical cord blood infusion for type 1 diabetes. Exp Hematol. 2008;36(6):710-715. http://dx.doi.org/10.1016/j.exphem.2008.01.009 [ Links ]
10. Ahmad LA, Crandall JP. Type 2 diabetes prevention: A review. Clin Diabetes. 2010;28:53-59. http://dx.doi.org/10.2337/diaclin.28.2.53 [ Links ]
11. Maedler K, Donath MY Beta-cells in type 2 diabetes: A loss of function and mass. Horm Res. 2004;62(Suppl 3):67-73. http://dx.doi.org/10.1159/000080503 [ Links ]
12. Shih HP, Wang A, Sander M. Pancreas organogenesis: From lineage determination to morphogenesis. Annu Rev Cell Dev Biol. 2013;29:81-105. http://dx.doi.org/10.1146/annurev-cellbio-101512-122405 [ Links ]
13. Suckale J, Solimena M. Pancreas islets in metabolic signaling - Focus on the beta-cell. Front Biosci J Virtual Libr. 2008;13:7156-7171. http://dx.doi.org/10.2741/3218 [ Links ]
14. Murtaugh LC. Pancreas and beta-cell development: From the actual to the possible. Dev Camb Engl. 2007;134(3):427-438. http://dx.doi.org/10.1242/dev.02770 [ Links ]
15. Desgraz R, Herrera PL. Pancreatic neurogenin 3-expressing cells are unipotent islet precursors. Development. 2009;136(21):3567-3574. http://dx.doi.org/10.1242/dev.039214 [ Links ]
16. Benitez CM, Goodyer WR, Kim SK. Deconstructing pancreas developmental biology. Cold Spring Harb Perspect Biol. 2012;4(6):a012401. http://dx.doi.org/10.1101/cshperspect.a012401 [ Links ]
17. Bosco D, Armanet M, Morel P Niclauss N, Sgroi A, Muller YD, et al. Unique arrangement of α- and β-cells in human islets of Langerhans. Diabetes. 2010;59(5):1202-1210. http://dx.doi.org/10.2337/db09-1177 [ Links ]
18. Brissova M, Fowler MJ, Nicholson WE, Chu A, Hirshberg B, Harlan DM, et al. Assessment of human pancreatic islet architecture and composition by laser scanning confocal microscopy. J Histochem Cytochem. 2005;53(9):1087-1097. http://dx.doi.org/10.1369/jhc.5C6684.2005 [ Links ]
19. Sosa-Pineda B, Chowdhury K, Torres M, Oliver G, Gruss P. The Pax4 gene is essential for differentiation of insulin-producing beta cells in the mammalian pancreas. Nature. 1997;386(6623):399-402. http://dx.doi.org/10.1038/386399a0 [ Links ]
20. Holland PWH, Booth HAF, Bruford EA. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007;5:47. http://dx.doi.org/10.1186/1741-7007-5-47 [ Links ]
21. Smith SB, Ee HC, Conners JR, German MS. Paired-homeodomain transcription factor PAX4 acts as a transcriptional repressor in early pancreatic development. Mol Cell Biol. 1999;19(12):8272-8280. [ Links ]
22. Boquist L, Edstrom C. Ultrastructure of pancreatic acinar and islet parenchyma in rats at various intervals after duct ligation. Virchows Arch A. 1970;349(1):69-79. http://dx.doi.org/10.1007/BF00548522 [ Links ]
23. Rosenberg L, Brown RA, Duguid WP. A new approach to the induction of duct epithelial hyperplasia and nesidioblastosis by cellophane wrapping of the hamster pancreas. J Surg Res. 1983;35(1):63-72. http://dx.doi.org/10.1016/0022-4804(83)90127-0 [ Links ]
24. Ferris WF, Woodroof CW, Louw J, Wolfe-Coote SA. Brief occlusion of the main pancreatic duct rapidly initiates signals which lead to increased duct cell proliferation in the rat. Cell Biol Int. 2001;25(1):113-117. http://dx.doi.org/10.1006/cbir.2000.0683 [ Links ]
25. Woodroof CW, De Villiers C, Page BJ, Van der Merwe L, Ferris WF. Islet neogenesis is stimulated by brief occlusion of the main pancreatic duct. S Afr Med J. 2004;94(1):54-57. http://dx.doi.org/10.1080/22201009.2004.10872330 [ Links ]
26. Brockenbrough JS, Weir GC, Bonner-Weir S. Discordance of exocrine and endocrine growth after 90% pancreatectomy in rats. Diabetes. 1988;37(2):232-236. http://dx.doi.org/10.2337/diab.37.2.232 [ Links ]
27. Bonner-Weir S, Baxter LA, Schuppin GT, Smith FE. A second pathway for regeneration of adult exocrine and endocrine pancreas. A possible recapitulation of embryonic development. Diabetes. 1993;42(12):1715-1720. http://dx.doi.org/10.2337/diab.42.12.1715 [ Links ]
28. Wang RN, Kloppel G, Bouwens L. Duct- to islet-cell differentiation and islet growth in the pancreas of duct-ligated adult rats. Diabetologia. 1995;38(12):1405-1411. http://dx.doi.org/10.1007/BF00400600 [ Links ]
29. Bertelli E, Bendayan M. Intermediate endocrine-acinar pancreatic cells in duct ligation conditions. Am J Physiol. 1997;273(5 Pt 1):C1641-C1649. [ Links ]
30. Gu D, Lee MS, Krahl T, Sarvetnick N. Transitional cells in the regenerating pancreas. Dev Camb Engl. 1994;120(7):1873-1881. [ Links ]
31. Gu D, Arnush M, Sarvetnick N. Endocrine/exocrine intermediate cells in streptozotocin-treated Ins-IFN-gamma transgenic mice. Pancreas. 1997;15(3):246-250. http://dx.doi.org/10.1097/00006676-199710000-00005 [ Links ]
32. Lardon J, Huyens N, Rooman I, Bouwens L. Exocrine cell transdifferentiation in dexamethasone-treated rat pancreas. Virchows Arch Int J Pathol. 2004;444(1):61-65. http://dx.doi.org/10.1007/s00428-003-0930-z [ Links ]
33. Mashima H, Ohnishi H, Wakabayashi K, Mine T, Miyagawa J, Hanafusa T, et al. Betacellulin and activin A coordinately convert amylase-secreting pancreatic AR42J cells into insulin-secreting cells. J Clin Invest. 1996;97(7):1647-1654. http://dx.doi.org/10.1172/JCI118591 [ Links ]
34. Thorel F, Népote V Avril I, Kohno K, Desgraz R, Chera S, et al. Conversion of adult pancreatic α-cells to β-cells after extreme β-cell loss. Nature. 2010;464(7292):1149-1154. http://dx.doi.org/10.1038/nature08894 [ Links ]
35. Desai BM, Oliver-Krasinski J, De Leon DD, Farzad C, Hong N, Leach SD, et al. Preexisting pancreatic acinar cells contribute to acinar cell, but not islet beta cell, regeneration. J Clin Invest. 2007;117(4):971-977. http://dx.doi.org/10.1172/JCI29988 [ Links ]
36. Solar M, Cardalda C, Houbracken I, Martin M, Maestro MA, De Medts N, et al. Pancreatic exocrine duct cells give rise to insulin-producing beta cells during embryogenesis but not after birth. Dev Cell. 2009;17(6):849-860. http://dx.doi.org/10.1016/j.devcel.2009.11.003 [ Links ]
37. Banakh I, Gonez LJ, Sutherland RM, Naselli G, Harrison LC. Adult pancreas side population cells expand after β cell injury and are a source of insulin-secreting cells. PLoS ONE. 2012;7(11):e48977. http://dx.doi.org/10.1371/journal.pone.0048977 [ Links ]
38. Xu X, D'Hoker J, Stangé G, Bonne S, De Leu N, Xiao X, et al. β cells can be generated from endogenous progenitors in injured adult mouse pancreas. Cell. 2008;132(2):197-207. http://dx.doi.org/10.1016/j.cell.2007.12.015 [ Links ]
39. Zuba-Surma EK, Kucia M, Wu W, Klich I, Lillard JW, Ratajczak J, et al. Very small embryonic-like stem cells are present in adult murine organs: ImageStream-based morphological analysis and distribution studies. Cytom Part J Int Soc Anal Cytol. 2008;73A(12):1116-1127. http://dx.doi.org/10.1002/cyto.a.20667 [ Links ]
40. Bhartiya D, Mundekar A, Mahale V, Patel H. Very small embryonic-like stem cells are involved in regeneration of mouse pancreas post-pancreatectomy. Stem Cell Res Ther. 2014;5(5):106. http://dx.doi.org/10.1186/scrt494 [ Links ]
41. Li W-C, Rukstalis JM, Nishimura W, Tchipashvili V Habener JF, Sharma A, et al. Activation of pancreatic-duct-derived progenitor cells during pancreas regeneration in adult rats. J Cell Sci. 2010;123(16):2792-2802. http://dx.doi.org/10.1242/jcs.065268 [ Links ]
42. Xiao X, Chen Z, Shiota C, Prasadan K, Guo P El-Gohary Y et al. No evidence for β cell neogenesis in murine adult pancreas. J Clin Invest. 2013;123(5):2207-2217. http://dx.doi.org/10.1172/JCI66323 [ Links ]
43. Dor Y Brown J, Martinez OI, Melton DA. Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation. Nature. 2004;429(6987):41-l6. http://dx.doi.org/10.1038/nature02520 [ Links ]
44. Meier JJ, Butler AE, Saisho Y, Monchamp T, Galasso R, Bhushan A, et al. β-cell replication is the primary mechanism subserving the postnatal expansion of β-cell mass in humans. Diabetes. 2008;57(6):1584-1594. http://dx.doi.org/10.2337/db07-1369 [ Links ]
45. Bonner-Weir S, Li W-C, Ouziel-Yahalom L, Guo L, Weir GC, Sharma A. β-cell growth and regeneration: Replication is only part of the story. Diabetes. 2010;59(10):2340-2348. http://dx.doi.org/10.2337/db10-0084 [ Links ]
46. Bonner-Weir S. Life and death of the pancreatic beta cells. Trends Endocrinol Metab. 2000;11(9):375-378. http://dx.doi.org/10.1016/S1043-2760(00)00305-2 [ Links ]
47. Brennand K, Melton D. Slow and steady is the key to β-cell replication. J Cell Mol Med. 2009;13(3):472. http://dx.doi.org/10.1111/j.1582-4934.2008.00635.x [ Links ]
48. Granger A, Kushner JA. Cellular origins of β-cell regeneration: A legacy view of historical controversies. J Intern Med. 2009;266(4):325-338. http://dx.doi.org/10.1111/j.1365-2796.2009.02156.x [ Links ]
49. Cano DA, Rulifson IC, Heiser PW, Swigart LB, Pelengaris S, German M, et al. Regulated β-cell regeneration in the adult mouse pancreas. Diabetes. 2007;57(4):958-966. http://dx.doi.org/10.2337/db07-0913 [ Links ]
50. Rankin MM, Wilbur CJ, Rak K, Shields EJ, Granger A, Kushner JA. β-cells are not generated in pancreatic duct ligation-induced injury in adult mice. Diabetes. 2013;62(5):1634-1645. http://dx.doi.org/10.2337/db12-0848 [ Links ]
 Correspondence:
Correspondence:
William F. Ferris
Division of Endocrinology
Department of Medicine
Faculty of Medicine and
Health Sciences, Stellenbosch
University, PO Box 19063
Tygerberg 7505
South Africa
Email: Wferris@sun.ac.za
Received: 22 Oct. 2014
Revised: 10 Nov. 2014
Accepted: 15 Nov. 2014

