










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.111 no.3-4 Pretoria Mar./Abr. 2015
http://dx.doi.org/10.17159/sajs.2015/20140309
RESEARCH LETTER
Youngest dinocephalian fossils extend the Tapinocephalus Zone, Karoo Basin, South Africa
Michael O. DayI; Saniye GüvenI; Fernando AbdalaI; Sifelani JirahI; Bruce RubidgeI; John AlmondII
IEvolutionary Studies Institute, School of Geosciences, University of the Witwatersrand, Johannesburg, South Africa
IINatura Viva, Cape Town, South Africa
ABSTRACT
The dinocephalians (Synapsida, Therapsida) were one of the dominant tetrapod groups of the Middle Permian (Guadalupian Epoch, ~270-260 million years ago) and are most abundantly recorded in the Tapinocephalus Assemblage Zone (AZ) of the Main Karoo Basin, South Africa. Dinocephalians are thought to have become extinct near the top of the Abrahamskraal Formation of the Beaufort Group and their disappearance is one criterion used to define the base of the overlying Pristerognathus AZ. Because of the abundance of fossils in the Karoo, the Beaufort Group biozones form the biostratigraphic standard for later Permian terrestrial tetrapod ecosystems, so their stratigraphic delineation is of great importance to Permian palaeobiology. We report two new specimens of the rare tapinocephalid dinocephalian Criocephalosaurus from the lowermost Poortjie Member, which makes them the youngest dinocephalians known from the Main Karoo Basin and extends the Tapinocephalus AZ from the Abrahamskraal Formation up into the Teekloof Formation. The extension of the Tapinocephalus AZ relative to the lithostratigraphy potentially affects the biozone or biozones to which a fossil species can be attributed; this extension has implications for biostratigraphic correlations within the Main Karoo Basin as well as with other basins across Gondwana. These discoveries also indicate that a population of herbivorous tapinocephalids survived as rare constituents of the tetrapod fauna after most generic richness within the clade had already been lost.
Keywords: Tapinocephalus; Middle Permian; Abrahamskraal; biostratigraphy; Pristerognathus
Introduction
The Dinocephalia are a clade of mostly large basal therapsids1 that were widely distributed around Pangaea in the Guadalupian. The group is known from Middle Permian basins in Russia, Central Asia, China, Brazil, Tanzania, Zambia and Zimbabwe2-10, but is best represented in the Abrahamskraal Formation of the South African Beaufort Group11 (Figure 1). Three dinocephalian families have been found in South Africa: the Anteosauridae, the Titanosuchidae and the Tapinocephalidae. The clade is most abundant and taxonomically diverse in the Tapinocephalus Assemblage Zone (AZ) - which has yielded all known species of titanosuchids and 18 of the 22 described species of tapinocephalids - but they are also found in the underlying Eodicynodon AZ in which they are represented by only two species, Tapinocaninus pamelae and Australosyodon nyaphuli.12,13 Together, the Eodicynodon and Tapinocephalus AZs are the oldest two biozones of the Beaufort Group.11
Although dinocephalians were an important constituent of early therapsid faunas, they disappear from the fossil record at the top of the Middle Permian Tapinocephalus AZ11,14,15 and in the Main Karoo Basin their last occurrence is a major criterion defining the base of the overlying Pristerognathus AZ16. Lithostratigraphically, the Tapinocephalus AZ corresponds to the upper Abrahamskraal Formation, apart from its uppermost strata, while the overlying Pristerognathus AZ is considered to extend through the uppermost part of the Abrahamskraal Formation and the Poortjie Member of the Teekloof Formation.17 Changes in the lithostratigraphic position of the contact between the Tapinocephalus and Pristerognathus AZs have consequences for stratigraphic correlation, as well as palaeobiodiversity studies, because the biozones of the Beaufort Group represent a global reference for terrestrial tetrapod faunas from the Middle Permian to the Early Triassic.18-21 Furthermore, radiometric dates have recently been determined for several zircon-bearing volcanic ash horizons within the Beaufort Group,22 so accurate chronology of tetrapod evolutionary patterns in the Permian is dependent on good biostratigraphic resolution relative to the lithostratigraphy.
Two dinocephalian crania, one with associated postcranial fragments including two articulated vertebrae with a high neural spine, have recently been recovered from the basal Poortjie Member of the Teekloof Formation. These discoveries provide new biostratigraphic data that necessitates a shift in the lithostratigraphic extent of the Tapinocephalus AZ.
Institutional Abbreviations
BP, Evolutionary Studies Institute (formerly Bernard Price Institute for Palaeontological Research), Johannesburg; CGS, Council for Geosciences, Pretoria; KM, McGregor Museum, Kimberley; NHMUK, Natural History Museum, London, UK; SAM, Iziko: South African Museum, Cape Town.
Material
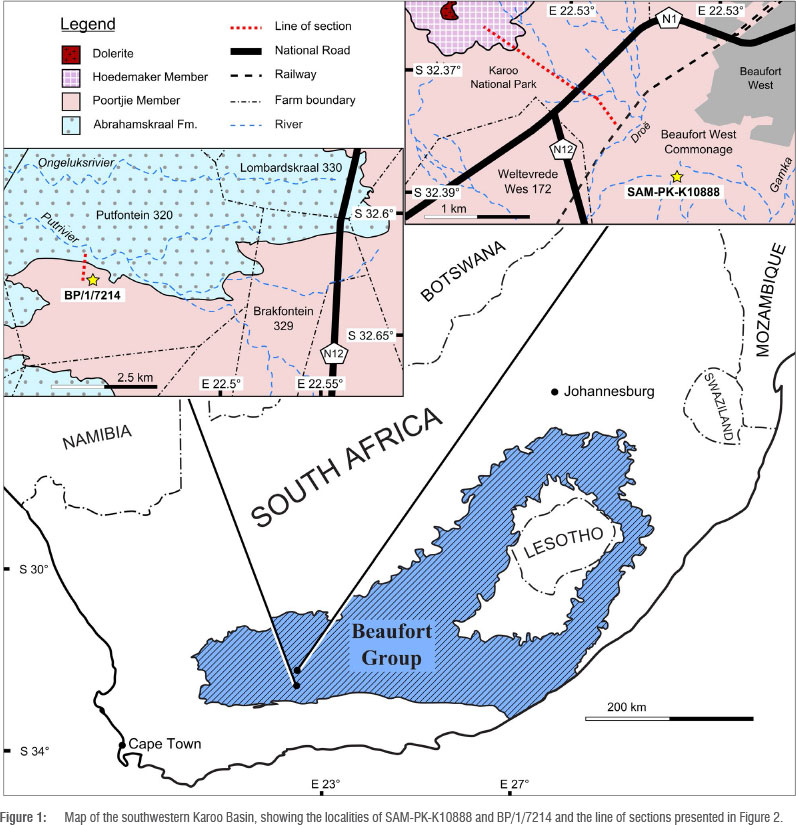
SAM-PK-K10888 is a partial skull and some associated postcranial elements that were discovered by J.A. and Madelon Tusenius on the Beaufort West commonage, Western Cape Province, to the southwest of that town (32° 23.25' S, 22° 33.24' E). Although the specimen was found ex situ, the low relief of the area and the sub-horizontal strata constrain the provenance of the specimen to a mudrock horizon exposed on the southern side of the valley (Figure 1). Reference to the published 1:250 000 geological map for the area (1979, Beaufort West Sheet, Geological Survey of South Africa 1:250000 Series), combined with stratigraphic fieldwork in the area by the authors, shows that this locality is within the lower Poortjie Member of the Teekloof Formation. As the combination of low relief and poor rock exposure was not conducive to measuring a stratigraphic section between the locality and the base of the Poortjie Member, a stratigraphic section was instead measured between the locality and the base of the overlying Hoedemaker Member, using a Jacob's staff and Abney level. This section indicated that the specimen occurred in a horizon 116 m below the top of the Poortjie Member (Figure 2). The Poortjie Member in the Beaufort West area is 160-182 m thick,23,24 in which case this specimen can be constrained to 40-70 m above the base of the Poortjie Member. This position is consistent with altitude readings for the locality and the mapped base of the Teekloof Formation (1979, Beaufort West Sheet, Geological Survey of South Africa 1:250000 Series), which provides an estimate of 50-60 m, although this may be affected by minor local folding.
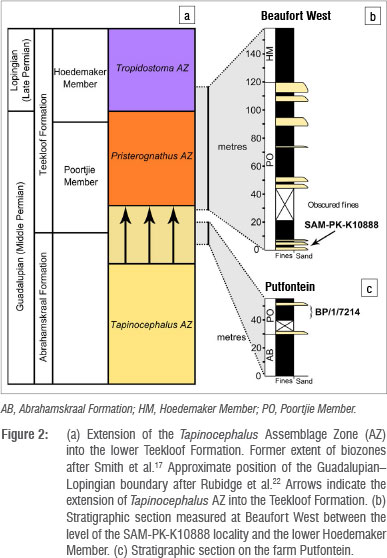
The second specimen, BP/1/7214, is also a partial skull and was found by M.D. and Christen Shelton, 70 km further south of Beaufort West on the farm Putfontein, Beaufort West district (32° 37.644' S, 22° 26.366' E). Like SAM-PK-K10888, BP/1/7214 was found ex situ in a stream bed but its provenance can be constrained to a 10-m thick mudrock horizon immediately below the second major sandstone body of the Poortjie Member. The specimen therefore originated 10-20 m above the base of the Poortjie Member (Figure 2).
Description
SAM-PK-K10888 and BP/1/7214 have very similar morphologies (Figure 3). Both comprise the posterior two-thirds of a skull roof up to the posterior border of the orbits and the occiput, although BP/1/7214 is more extensively weathered. SAM-PK-K10888 also preserves the basicranium (Figure 3d). In dorsal view, both specimens display a broadly elliptical shape resulting from the broad frontals being bordered by the narrow interorbital region anteriorly and the narrow temporal roof posteriorly (Figure 3a, 3e).
Although both specimens lack the anterior portion of the skull, they are clearly identified as tapinocephalid dinocephalians by the anteroventral rotation of the occiput and by the presence of pronounced cranial pachyostosis. Furthermore, SAM-PK-K10888 and BP/1/7214 present a set of characters that allow their referral to the tapinocephalid genus Criocephalosaurus (replacement name for Criocephalus since 2002).25,26 These include the close proximity of the orbits to one another and their anterior position on the skull compared to other tapinocephalids, a long pineal canal, and an acute angle between the median line of the occiput and the skull roof.9'25'27,28 Compared with other specimens of Criocephalosaurus, the cranial roof of both SAM-PK-K10888 and BP/1/7214 displays moderate pachyostosis that increases the cranial width across the frontals and deepens the parietal along the pineal canal, as is the case in the genotype (KM5138) and the referred specimen (NHMUK R36626). However, the preserved portion of the orbital region in both SAM-PK-K10888 and BP/1/7214 also shows only minor thickening of the prefrontal dorsally and of the postorbital posteriorly (Figure 3b, 3c, 3f and 3g), unlike other examples of Criocephalosaurus. This difference may be related to their size, as the two new specimens are the smallest representatives of the genus, with postorbital skull lengths (from the posterior border of the orbit to the posterior tip of the temporal roof) of 160 mm (SAM-PK-K10888) and 169 mm (BP/1/7214).
The parietal in both specimens is a small but deep bone extending across the posteriorly narrowing intertemporal region. In this latter character, SAM-PK-K10888 and BP/1/7214 differ from KM5138, CGP/1/846 and NHMUK R36626, in which the intertemporal region is broad throughout. In SAM-PK-K10888, the posterior portion of the cranium is fractured, creating a roughly sagittal section through the cranial roof, allowing the long pineal canal to be seen (Figure 3h). The canal is orientated parallel to the long axis of the cranium, as in NHMUK R36626. The posteriorly facing pineal opening is large and is surrounded by a thick rim, which is elevated from the parietal surface as in KM5138, CGP/1/846 and NHMUK R36626; this rim contributes to the conical appearance of the posterior end of the skull. The preserved portion of the occiput of SAM-PK-K10888 is formed by the tabular and postparietal bones (Figure 3d), which are elongated dorsoventrally as in the only other specimen of Criocephalosaurus that possesses a comparably well-preserved occiput (NHMUK R36626).
Discussion and conclusions
The genus Criocephalosaurus has two described species: C. vanderbyli Broom 1928, the holotype of which is a weathered cranial roof, and C. gunyankaensis Boonstra 1968, which was based on four cranial roofs briefly described by Boonstra9 and only later referred to Criocephalus.28 The latter specimens are now all missing. A few other incomplete crania were referred to the genus by Boonstra28, but a survey of museum collections has yielded only four specimens that can reliably be identified as Criocephalosaurus: KM5138 (holotype), CGP/1/846, NHMUK R36626 and BP/1/1582. Although specimens SAM-PK-K10888 and BP/1/7214 can be confidently identified as Criocephalosaurus, their poor state of preservation does not allow their identification to species level.
Both SAM-PK-K10888 and BP/1/7214 fall within the lower size range of tapinocephalids and are the smallest known examples of Criocephalosaurus. Both specimens show only a moderate degree of pachyostosis of the postorbital bar and cranial roof. The latter is steadily reduced posteriorly from the level of the mid-temporal fenestra leading to the conical appearance of the back of the skulls (best illustrated in Figure 3e). Both the overall lesser extent of pachyostosis and the pronounced conical form of the skull differentiate SAM-PK-K10888 and BP/1/7214 from larger specimens, although CGP/1/846 demonstrates a slight tapering of the posterior skull roof towards the pineal opening. Because the degree of pachyostosis is associated with ontogeny in tapinocephalids,29,30 it is possible that these two specimens represent sub-adults; however, their incompleteness prevents the definitive identification of unambiguous juvenile morphology (e.g. unfused sutures). The fact that both specimens bearing the more gracile morphology occur in the lower Poortjie Member, whereas all larger existing specimens occur in the underlying Abrahamskraal Formation, may also be of significance but this can only be elucidated by the discovery of further material.
The stratigraphic position of the Tapinocephalus-Pristerognathus AZ boundary relies on the current definition of the Pristerognathus AZ, which is described as 'low diversity dominated by [the dicynodont] Diictodon in association with [the therocephalian] Pristerognathus and the absence of dinocephalian fossils that are a prominent component of the underlying fauna'16. Because both Diictodon and Pristerognathus are known from the underlying Tapinocephalus AZ and the dominance of Diictodon continues into the overlying Tropidostoma AZ,17 the Pristerognathus AZ can only maintain its integrity when described as an 'interval zone'31 between the last appearance of dinocephalians and the first appearance of the dicynodont Tropidostoma. This lack of new taxa in comparison with the Tapinocephalus AZ has in fact led to doubt over the validity of the Pristerognathus AZ.21
Although it is acknowledged that a biostratigraphic review is required for this interval, the work for which has already begun,32 the last occurrence of the previously dominant dinocephalians remains a biostratigraphically important event. Despite reservations over the definition of the Pristerognathus AZ, we therefore recognise a biostratigraphic distinction between the dinocephalian bearing Tapinocephalus AZ and the dinocephalian deficient assemblages above, at least within the Main Karoo Basin. The discovery of tapinocephalids in the lowermost Poortjie Member demonstrates that the dinocephalian record extends into the Teekloof Formation of the Beaufort Group (Figure 2a), and this notion is supported by a probable titanosuchid (BP/1/7184) from the lowermost Poortjie Member near Sutherland.32 Consequently, the Tapinocephalus AZ should be extended by a commensurate degree into the lower Poortjie Member, meaning its upper boundary is therefore correspondingly younger than previously understood.
Regionally, an upward shift in the stratigraphic position of the Tapinocephalus-Pristerognathus AZ boundary has implications for the biozone designation of fossil material in existing collections and thus of individual genera. For instance, the extension of the Tapinocephalus AZ up into the lower Poortjie Member could constrain the therocephalian Scylacosaurus (through specimen SAM-PK-10530) and varanopid synapsids (through specimen SAM-PK-K10407) to the Tapinocephalus AZ.32,33 Globally, this shift could potentially impact palaeobiological studies of Permian tetrapods because biozones are often used as approximations of 'time bins' in calculations of taxonomic diversity trends and those of other biological variables.17,3437 Because they represent the youngest dinocephalian fossils from the Main Karoo Basin, the new fossils may also help to elucidate the mode by which dinocephalians became extinct, and, by extension, the cause of the global disappearance of the clade.38 In particular, the presence of Criocephalosaurus in the Teekloof Formation, above the range of any other dinocephalian genus, suggests that a loss of species richness within dinocephalians preceded their extinction.
Acknowledgements
We thank the Palaeontological Scientific Trust (PAST) and its Scatterlings of Africa programmes, and the African Origins platform of the NRF for the generous funding which enabled this research to be undertaken. The support of the DST/NRF Centre of Excellence in Palaeosciences (CoE in Palaeosciences) towards this research is hereby acknowledged. Opinions expressed and conclusions arrived at are those of the authors and are not necessarily to be attributed to the CoE in Palaeosciences. We also thank Christen Shelton for his role in finding the second half of BP/1/7214, Madelon Tusenius for the co-discovery of SAM-PK-K10888 and Roger Smith who led the post-conference fieldtrip during which specimen BP/1/7214 was discovered. We are grateful to Spencer Lucas and an anonymous reviewer for comments that helped improve the manuscript.
Authors' contributions
M.O.D. determined the stratigraphic position of the fossils; S.G. described and identified the fossil material; B.R. was the project leader, provided funding for and intellectual input to the study and co-supervised S.G.; F.A. co-supervised S.G.; J.A. and M.D. discovered the new specimens of Criocephalosaurus examined in the study; M.D. and S.J. measured the stratigraphic sections; M.D. and S.G. wrote the first draft of the manuscript; and all authors contributed to the preparation of the final draft.
References
1. Rubidge BS, Sidor CA. Evolutionary patterns among Permo-Triassic therapsids. Annu Rev Ecol Syst. 2001;32:449-480. http://dx.doi.org/10.1146/annurev.ecolsys.32.081501.114113 [ Links ]
2. Battail B, Surkov MV. Mammal-like reptiles from Russia. In: Benton MJ, Shishkin MA, Unwin DM, editors. The age of dinosaurs in Russia and Mongolia. Cambridge: Cambridge University Press; 2000. p. 86-119. [ Links ]
3. Cheng Z, Li J. First record of a primitive anteosaurid dinocephalian from the Upper Permian of Gansu, China. Vertebrat Palasiatic. 1997;34:123-134. [ Links ]
4. Cheng Z, Li J. A new genus of primitive dinocephalian - The third report on Late Permian Dashankou lower tetrapod fauna. Vertebrat Palasiatic. 1997;35:35-43. [ Links ]
5. Langer MC. The first record of dinocephalians in South America, Late Permian (Rio do Rosto Formation) of the Parana Basin, Brazil. Neues Jahrb Geol Palãontol Abh. 2000;215:69-95. [ Links ]
6. Cisneros JC, Abdala F, Atayman-Güven S, Rubidge BS, Şengör AMC, Schultz CL. Carnivorous dinocephalian from the Middle Permian of Brazil and tetrapod dispersal in Pangaea. Proc Natl Acad Sci USA. 2012;109:1584-1588. http://dx.doi.org/10.1073/pnas.1115975109 [ Links ]
7. Simon RV, Sidor CA, Angielczyk KD, Smith RMH. First record of a tapinocephalid (Therapsida: Dinocephalia) from the Ruhuhu Formation (Songea Group) of southern Tanzania. J Vertebr Paleontol. 2010;30:1289-1293. http://dx.doi.org/10.1080/02724634.2010.483549 [ Links ]
8. Sidor CA, Angielczyk KD, Smith RM, Goulding AK, Nesbitt SJ, PeecookBR, et al. Tapinocephalids (Therapsida, Dinocephalia) from the Permian Madumabisa Mudstone Formation (Lower Karoo, Mid-Zambezi Basin) of southern Zambia. J Vertebr Paleontol. 2014;34:980-986. http://dx.doi.org/10.1080/02724634.2013.826669 [ Links ]
9. Boonstra LD. Report on some reptilian fossils from Gunyanka's Kraal, Busi Valley. Trans Rhodesia Sci Assoc. 1946;41:1-4. [ Links ]
10. Lepper J, Raath MA, Rubidge BS. A diverse dinocephalian fauna from Zimbabwe. S Afr J Sci. 2000;96:403-405. [ Links ]
11. Rubidge BS. Re-uniting lost continents - Fossil reptiles from the ancient Karoo and their wanderlust. S Afr J Geol. 2005;108:135-172. [ Links ]
12. Rubidge BS. A new primitive dinocephalian mammal-like reptile from the Permian of South Africa. Palaeontology. 1991;34:547-559. [ Links ]
13. Rubidge BS. Australosyodon, the first primitive anteosaurid dinocephalian from the Upper Permian of Gondwana. Palaeontology. 1994;37:579-594. [ Links ]
14. Boonstra LD. The fauna of the Tapinocephalus zone (Beaufort beds of theKaroo). Ann S Afr Mus. 1969;56:1-73. [ Links ]
15. Keyser AW, Smith RHM. Vertebrate biozonation of the Beaufort Group with special reference to the Western Karoo Basin. Ann Geol Surv S Afr. 1979;12:1-36. [ Links ]
16. Smith RMH, Keyser AW. Biostratigraphy of the Pristerognathus Assemblage Zone. In: Rubidge BS, editor. Biostratigraphy of the Beaufort Group (Karoo Supergroup), South African Committee for Stratigraphy, Biostratigraphic Series 1. Pretoria: Council for Geoscience; 1995. p. 13-17. [ Links ]
17. Smith RHM, Rubidge BS, Van Der Walt MVM. Therapsid biodiversity patterns and palaeoenvironments of the Karoo Basin, South Africa. In: Chinsamy-Turan A, editor. Forerunners of mammals. Bloomington and Indianapolis: Indiana University Press; 2012. p. 31-64. [ Links ]
18. Benton MJ. No gap in the Middle Permian record of terrestrial vertebrates. Geology. 2012;40:339-342. http://dx.doi.org/10.1130/G32669.1 [ Links ]
19. Fröbisch J. Global taxonomic diversity of anomodonts (Tetrapoda, Therapsida) and the terrestrial rock record across the Permian-Triassic boundary. PLoS ONE. 2008;3(11):e3733. http://dx.doi.org/10.1371/journal.pone.0003733 [ Links ]
20. Lucas SG. Global Triassic tetrapod biostratigraphy and biochronology. Palaeogeogr Palaeocl. 1998;143:347-384. http://dx.doi.org/10.1016/S0031-0182(98)00117-5 [ Links ]
21. Lucas SG. Global Permian tetrapod biostratigraphy and biochronology. Geol Soc Spec Publ. 2006;265:65-93. http://dx.doi.org/10.1144/GSL.SP.2006.265.01.04 [ Links ]
22. Rubidge BS, Erwin D, Ramezani J, Browning SA, De Klerk, WJ. High-precision temporal calibration of Late Permian vertebrate biostratigraphy: U-Pb zircon constrains from the Karoo Supergroup, South Africa. Geology. 2013;41(3):363-366. http://dx.doi.org/10.1130/G33622.1 [ Links ]
23. Cole DI, Wipplinger PE. Sedimentology and molybdenum potential of the Beaufort Group in the main Karoo Basin, South Africa. Counc Geosci Mem. 2001;80:1-225. [ Links ]
24. Stear WM. The sedimentary environment of the Beaufort Group uranium province in the vicinity of Beaufort West [PhD thesis]. Port Elizabeth:University of Port Elizabeth; 1980. [ Links ]
25. Broom R. On Tapinocephalus and two other dinocephalians. Ann S Afr Mus. 1928;22:427-438. [ Links ]
26. Kammerer C, Sidor CA. Replacement names for the therapsid genera Criocephalus Broom 1928 and Olivieria Brink 1965. Palaeontologia Africana. 2002;38:71-72. [ Links ]
27. Boonstra LD. Diversity within the South African Dinocephalia. S Afr J Sci. 1963;59:196-206. [ Links ]
28. Boonstra LD. The braincase, basicranial axis and median septum in the Dinocephalia. Ann S Afr Mus. 1968;50(10):195-273. [ Links ]
29. Broom R. On some new South African Permian reptiles. Proc Zool Soc Lond. 1911;81(4):1073-1082. http://dx.doi.org/10.1111/j.1096-3642.1911.tb01976.x [ Links ]
30. Gregory WK. The skeleton of Moschops capensis Broom, a dinocephalian reptile from the Permian of South Africa. Bull Am Mus Nat Hist. 1926;56(3):179-251. [ Links ]
31. Murphy MA, Salvador A, editors. International Subcommission on Stratigraphic Classification (ISSC). International Stratigraphic Guide - An abridged edition. Episodes. 1999;22(4):255-271. [ Links ]
32. Day MO. Middle Permian continental biodiversity change as reflected in the Beaufort Group of South Africa: A bio- and lithostratigraphic review of the Eodicynodon, Tapinocephalus and Pristerognathus Assemblage Zones [PhDthesis]. Johannesburg: University of the Witwatersrand; 2013. [ Links ]
33. Modesto SP Smith RMH, Campione NE, Reisz RR. The last "pelycosaur": A varanopid synapsid from the Pristerognathus Assemblage Zone, Middle Permian of South Africa. Naturwissenschaften. 2011;98:1027-1034. http://dx.doi.org/10.1007/s00114-011-0856-2 [ Links ]
34. Huttenlocker AK. Body size reductions in non-mammalian eutheriodont therapsids (Synapsida) during the End-Permian mass extinction. PLoS ONE. 2014;9(2):e87553. http://dx.doi.org/10.1371/journal.pone.0087553 [ Links ]
35. Irmis RB, Whiteside JH. Delayed recovery of non-marine tetrapods after the end-Permian mass extinction tracks global carbon cycle. Proc R Soc B. 2012;279:1310-1318. http://dx.doi.org/10.1098/rspb.2011.1895 [ Links ]
36. Irmis RB, Whiteside JH, Kammerer KF. Non-biotic controls of observed diversity in the paleontologic record: An example from the Permo-Triassic Karoo Basin of South Africa. Palaeogeogr Palaeocl. 2013;372:62-77. http://dx.doi.org/10.1016/j.palaeo.2012.07.014 [ Links ]
37. Fröbisch J. Vertebrate diversity across the end-Permian mass extinction- Separating biological and geological signals. Palaeogeogr Palaeocl. 2013;372:50-61. http://dx.doi.org/10.1016/j.palaeo.2012.10.036 [ Links ]
38. Lucas SG. Timing and magnitude of tetrapod extinctions across the Permo-Triassic boundary. J Asian Earth Sci. 2009;36:491-502. http://dx.doi.org/10.1016/j.jseaes.2008.11.016 [ Links ]
 Correspondence:
Correspondence:
Michael Day
Evolutionary Studies Institute
University of the Witwatersrand
Private Bag 3, Johannesburg 2050
South Africa
Email: michael.day@wits.ac.za
Received: 04 Sep. 2014
Revised: 15 Dec. 2014
Accepted: 03 Jan. 2015


{kind=link}