










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.110 no.9-10 Pretoria oct. 2014
http://dx.doi.org/10.1590/sajs.2014/20130369
RESEARCH ARTICLE
Long bone cross-sectional geometric properties of Later Stone Age foragers and herder-foragers
Michelle E. CameronI, II; Susan PfeifferII, III
IDepartment of Archaeology and Anthropology, University of Cambridge, Cambridge, United Kingdom
IIDepartment of Anthropology, University of Toronto, Toronto, Ontario, Canada
IIIDepartment of Archaeology, University of Cape Town, Cape Town, South Africa
ABSTRACT
Diaphyseal cross-sectional geometry can be used to infer activity patterns in archaeological populations. We examined the cross-sectional geometric (CSG) properties of adult Later Stone Age (LSA) herder-forager long bones from the inland lower Orange River Valley of South Africa (n=5 m, 13 f). We then compared their CSG properties to LSA forager adults from the coastal fynbos (n=23 m, 14 f) and forest (n=17 m, 19 f) regions, building on a previous report (Stock and Pfeiffer, 2004). The periosteal mould method was used to quantify total subperiosteal area, torsional strength, bilateral asymmetry and diaphyseal circularity (Imax/Imin) at the mid-distal (35%) location of upper arms (humeri) and the mid-shaft (50%) location of upper legs (femora). Maximum humerus and femur lengths were similar among the three samples, suggesting that adult stature was similar in all three regions. When compared to the previous study, CSG property values obtained using the periosteal mould method correlated well, and there were no significant differences between data collected using the different methods. No statistically significant differences were found among the humerus or femur CSG properties from the different regions. This finding suggests that all individuals undertook similar volitional habitual activities in regard to their upper limbs, and also had similar degrees of terrestrial mobility. These results indicate relative behavioural homogeneity among LSA foragers and herder-foragers from South Africa. The small degree of regional variation apparent among the three samples may reflect local ecology and the subsistence demands affecting populations in these different regions.
Keywords: South Africa; Holocene; bioarchaeology; regional variation; habitual physical activity.
Introduction
Reconstruction of physical activity among Later Stone Age foragers
Holocene Later Stone Age (LSA) forager skeletons have been recovered archaeologically most often from the southern and southwestern coasts and coastal forelands of the South African Cape (Figure 1). They date from ca. 10 000 bp to the historical era.12 While these LSA populations share artefact traditions23, there are some pertinent ecological differences between coastal regions that appear to have been consistent throughout the Holocene2.
Along the southwestern Cape coast, fynbos ecology predominates (Figure 1), featuring succulents, a diverse plant base of geophytes and fruits, and a Mediterranean climate with rainfall of 200-600 mm per annum. Small browsers, with some larger grazing ungulates, dominate the faunal assemblages. Along the southern coast, afro-montane forest conditions predominate, although fynbos vegetation also persists (Figure 1). While the climate is similar to the fynbos, the evergreen canopy in the forest is continuous and rainfall is higher (500-1200 mm per annum). The forest has small browsers as well, although some larger species inhabited this area during the Holocene. Diverse marine resources are available in both regions.12 The southwestern region will be referred to as the 'fynbos' and the southern region will be referred to as the 'forest,' following the geographical definitions provided by Morris1.
The size and shape of adult LSA forager skeletons have interested many researchers. Studying a sample available 20 years ago, Smith et al.4 examined changes in LSA forager shaft diameter, cortical bone thickness, maximum humerus length and maximum femur length to detect if changes in diet and lifestyle associated with contact between forager and pastoralist groups may have impacted forager health and activity patterns between the hypothesised pre- and post-contact periods. They noted that cortical bone thickness and maximum bone lengths increased in the post-contact period, particularly in forager men, although these increases only reached significance for a few of the measures considered. About 15 years ago, Churchill and Morris5 examined differences in the habitual activities of LSA foragers from the fynbos, forest and savannah regions of South Africa by examining musculoskeletal stress markers. They predicted that physical activity and musculoskeletal stress markers would be higher in the forest, reflecting higher resource abundance. They reported a stronger pattern among men than among women. Ledger et al.6 used computed tomographic (CT) scans to compare long bone diaphyseal cross-sectional geometric (CSG) properties of LSA foragers to a sample of historical skeletons from Cape Town and a modern cadaveric collection. They found that male and female foragers had strong lower limbs and appeared to have been highly mobile and active compared to the more recent populations. Stock and Pfeiffer7 also examined variation in the habitual physical activity patterns of male and female LSA foragers from the fynbos and forest by analysing long bone diaphyseal CSG properties.
Cross-sectional geometric properties are presumed to reflect the strength and shape of the long bone. These properties develop during life, and are influenced by the intensity and frequency of physical activities undertaken. Consequently, they reflect the habitual activities and biomechanical patterns of an individual.8 Stock and Pfeiffer7 found differences in upper and lower limb strength between the men and women of the forest and fynbos samples. They concluded that there was a greater disparity in the intensity of manual labour between forest men and women than between fynbos men and women. Bilateral asymmetry differences between the men of these two regions suggested that forest men may have more commonly used spears for hunting and fynbos men may have more frequently used light-draw bows.7 These conclusions built on previous research into upper limb strength asymmetries in past populations and contemporary athletes.9,10 A conclusion of spear use, which is reflected in greater limb strength asymmetry, is consistent with the artefacts documented from the forest region.9,10 Small points, typical of fynbos assemblages, are consistent with the type of bow-and-arrow hunting ethnographically associated with the San foragers of southern Africa.3,7 Men from both the forest and fynbos displayed femur CSG properties consistent with levels of terrestrial mobility higher than those of women. Femur data further showed that women from the forest had CSG properties consistent with higher levels of terrestrial mobility than women from the fynbos.7
That study of the fynbos and forest regions has subsequently been used to compare habitual physical activities among diverse Holocene foragers. Stock11 compared the upper and lower limb CSG properties of LSA foragers to an early historical Andaman Island sample, as Andaman Islanders represent another small-bodied foraging population. He found that LSA foragers displayed greater lower limb CSG property values, likely as a result of greater terrestrial mobility, while Andaman Islanders displayed higher upper limb CSG property values, which was attributed to a high proportion of water-based activities, such as canoeing and swimming. He also compared LSA CSG properties to those of Yahgan foragers from Tierra del Fuego in South America. The Yahgan foragers displayed higher upper limb CSG property values, which may also reflect substantial watercraft use and swimming activities. Carlson et al.12 compared the CSG properties of Australian Aborigine foragers to those of the LSA. Australian Aborigines displayed generally weaker upper and lower limb CSG property values, suggesting that LSA foragers pursued terrestrial mobility-based foraging activities at higher intensities.12
In sum, LSA foragers appear to have been more terrestrially mobile than other groups, yet do not appear to have regularly engaged in activities requiring intense upper limb strength. With the exception of male arm strength, physical activity patterns were relatively consistent between two regions. To elaborate on this line of research, a LSA skeletal sample from a third region - the lower Orange River Valley - will be assessed. The inclusion of this sample will help to establish the range of variation in habitual physical activities that may have existed among geographically diverse Holocene populations of South Africa.
Later Stone Age herder-foragers of the lower Orange River Valley
The lower Orange River Valley is both geographically and ecologically distinct from the two more southerly coastal areas (Figure 1). The lower Orange River Valley will be referred to as the 'inland' region, as distinct from the fynbos and forest regions. The inland area is semi-arid, with less predictable rainfall and more extreme temperatures than the Cape. Ecologically, it is marginal between the Sweet Grassveld and Karoo types.13 Vegetation is relatively bushy and uneven, although it is capable of sustaining livestock if there is sufficient rainfall.13 Geophytic plants dominate the edible plant species. The lower Orange River Valley has a high proportion of browsers relative to areas with heavier rainfall where grazers may be more common.14 Therefore, inland foragers may have had distinctive diets, reflecting the ecological differences among the three areas being compared. Inland population density appears to have been much lower, likely because of sporadic rainfall and low availability of reliable resources.2
LSA skeletons have been recovered from Koffiefontein and Augrabies Falls, both located in the inland lower Orange River Valley.14 These sites date to the southern African proto-historical period, with one burial near Koffiefontein dating to 390 ¹ 50 bp (uncalibrated) radiocarbon years.14 Many skeletons from this region were found near Type-R settlements.1415 These settlements are associated with what has been characterised as a relatively sedentary lifestyle in which food resources were acquired through pastoralism as well as foraging.14 During this period, Khoe pastoralist and Bantu-speaking (black African) agriculturalists also occupied the lower Orange River Valley. Type-R settlement sites show some evidence of close contact and trade with Bantu-speaking agropastoralists.14
Morris14 studied the craniometric and dental characteristics of LSA herder-foragers from the lower Orange River Valley. Herder-foragers from this region display craniometric characteristics consistent with other LSA and San populations, implying that, genetically, inland men and women remained a unique population despite the presence of non-foraging groups in this area.14 Morphological evidence of some Bantu admixture was also noted, suggesting intermarriage with Bantuspeaking agriculturalists.14
An analysis of lower Orange River Valley (inland) LSA herder-forager postcranial skeletons may help to clarify the types of habitual activities undertaken by this group, and determine if their activities were distinct from other LSA foragers as a result of cultural differences, such as the incorporation of pastoralism. As herder-foragers may supplement their foraging efforts with provisions supplied by animal herds, lower mobility demands may have been placed on the legs, reflecting less searching for food resources. Upper limb strength could also be less crucial compared to other LSA groups, if less vigorous food processing was required. Examining whether inland herder-foragers had habitual physical activities similar to those of related foragers from the other regions may also help clarify whether contact with non-foraging populations affected herder-foragers' habitual activities.2,16-18
Biomechanical analyses of long bone shafts
Studies investigating the relationship between physical activity and bone remodelling are based on assumptions of bone functional adaptation. In this framework, bone remodelling is stimulated by the physical deformation of bone tissue through positive and negative feedback. Heighted physical activity and resulting tissue deformation leads to bone tissue deposition, while lessened physical activity leads to the resorption of bone tissue.19 Analyses of CSG properties, which characterise the distribution of bone tissue around long bone diaphyses, have been applied to skeletal materials originating from archaeological contexts, to reconstruct habitual activities undertaken by past human populations.6-8,12,20-22
CSG analyses focus on the application of beam theory to the quantification of long bone CSG properties.8,23-25 Information typically comes from long bone diaphyseal cross-sections in which periosteal and endosteal contours are modelled using CT scanners or a combination of silicone moulds and biplanar radiographs.8,23-26 However, while these analyses are accurate, high-resolution scanners are often unavailable in remote research locales and images may be expensive to produce.8,23-26
In response to these issues, the accuracy of external methods for quantifying CSG properties has been explored by examining the efficacy of relying solely on periosteal contours, represented by periosteal moulds.23-25 Because the strength of a long bone diaphysis is dependent on the amount and distribution of cortical bone present, a greater distance between the periosteal boundary and the diaphyseal centroid indicates a long bone's enhanced capacity to resist bending and torsional loading.8 23-25 Periosteal contours have been shown to have a stronger influence on a bone shaft's biomechanical characteristics than do endosteal contours.8,23-25 Moulding of periosteal contours with silicone impression material is non-invasive, does not damage skeletal tissue and relies on no biomedical technology. Validation of this approach for South African LSA foragers would corroborate other reports indicating that external methods can be used to obtain valid CSG properties, without reliance on images of endosteal contours.21-24
Aims
The accuracy of the periosteal mould method was examined by comparing newly generated values with previous values from the same LSA long bones used in a previous study by Stock and Pfeiffer7. The sample sizes for the fynbos and forest populations were also increased, as more skeletons had become available for analysis since Stock and Pfeiffer's study.7 Then, using this method, a new geographic comparison of CSG properties among LSA forager populations from the fynbos, forest and inland regions of South Africa was undertaken, to look for behavioural patterns among the samples. We sought to determine whether inland herder-foragers display habitual activity patterns comparable to those of foragers that did not practise any pastoralism. The results obtained in this study for the fynbos and forest samples should remain consistent with those of Stock and Pfeiffer7 despite the increase in sample sizes. Variation in habitual activity patterns may be low among and between the three samples, regardless of the incorporation of pastoralism inland, if LSA foraging is characterised by behavioural homogeneity.
Materials and methods
Later Stone Age forager and herder-forager samples
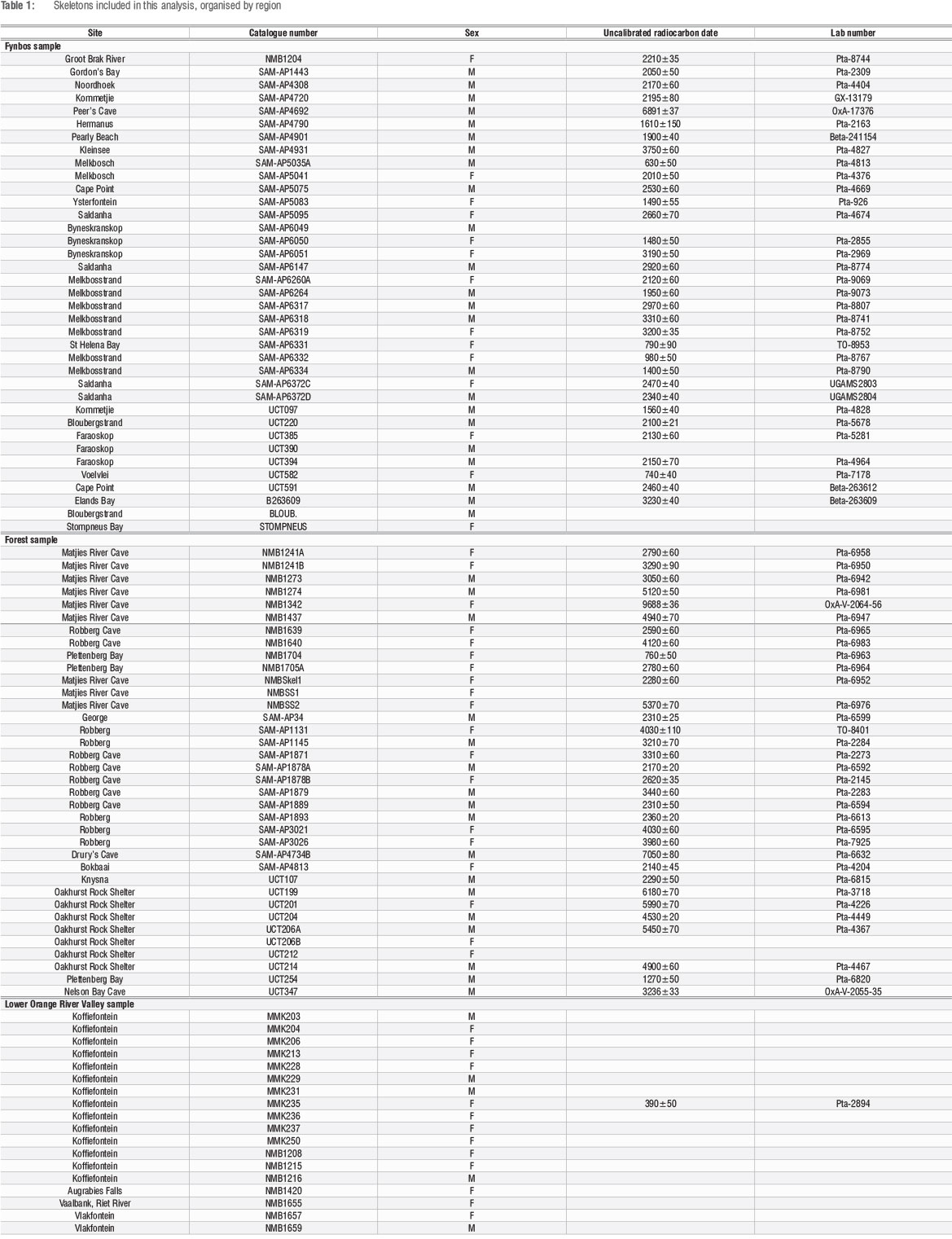
The samples include the humeri and femora of 91 adults (45 males, 46 females; Table 1) originating from archaeological sites in the fynbos (n=23 m, 14 f), forest (n=17 m, 19 f) and inland (n=5 m, 13 f) regions. The skeletons have been radiocarbon dated to between 10 000 bp and 300 bp (uncalibrated) (Table 1). CSG properties of many of the LSA foragers from the fynbos and forest have previously been examined but the sample sizes have increased since then. There are newly added fynbos skeletons (n=14 m, 9 f) and newly added forest skeletons (n=2 m, 1 f). Age and sex estimations are based on skeletal morphology, as observed by collaborating bioarchaeologists L.E. Doyle and C. Merritt who also documented postcranial dimensions.
The craniometric stability and homogeneity observed among LSA skeletons27 indicate that the three samples can be considered as a single population. To confirm postcranial size homogeneity across regions, one-way analyses of variance (ANOVAs) for maximum humerus and femur lengths were conducted for all samples.
Assessment of cross-sectional geometric properties
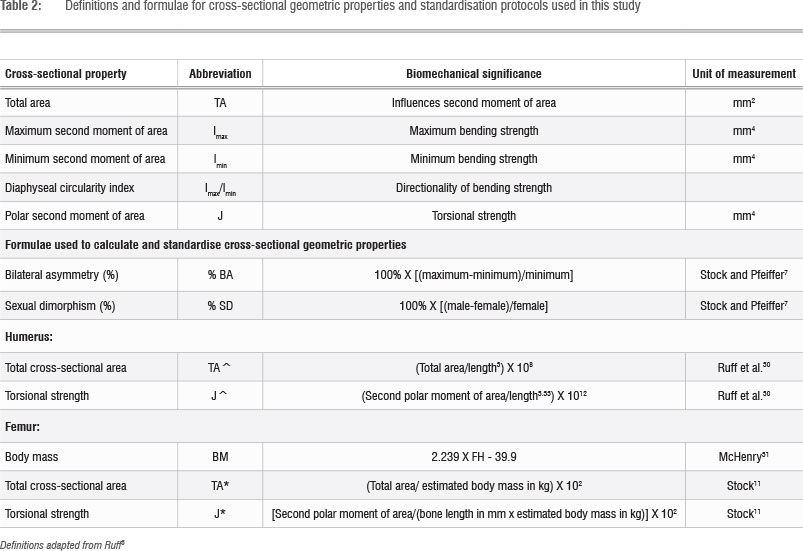
Periosteal moulds were taken using Exaflex Heavy Body silicone impression material (GC America, Alsip, IL, USA) at the mid-distal (35%) location of all humeri (bilaterally) and at the mid-shaft of one femur per individual, with the right femur preferred. Researchers commonly use the 35% location on the humerus shaft, thus avoiding large muscle attachment sites and allowing effective assessment of bilateral asymmetry. The left femur was used when the right was missing or incomplete. Assuming symmetry of the lower legs, values from left femora were included with values from the right femora in the analysis. Once mid-shaft and mid-distal locations had been identified for the femora and humeri, respectively, these locations were marked with string, and then the quick-drying silicone impression material was applied to the external surface of the bone. The moulds were then removed, resulting in a representation of the diaphyseal (shaft) cross section.
Anatomical planes of orientation were marked on each mould during the moulding process to help maintain correct orientation during digitisation. The moulds were digitised using an Epson flatbed scanner, including a ruler for scale. Periosteal contours were estimated by manually tracing along the edges of digitised moulds using a drawing tablet. The traced images were analysed using the ImageJ platform28 with the Moment Macro v1.3 plug-in29. CSG properties were calculated for each bone (Table 2).
The CSG variables examined in this study are listed in Table 2. Total subperiosteal area (TA) is a direct indicator of diaphyseal cross-sectional size. It influences the polar second moment of area, which reflects the bending strength of a long bone.8 TA also represents the most accurate measure that can be derived from the periosteal mould method. Torsional strength (J) represents the robustness of a diaphysis and is the best indicator of a bone's capacity to withstand diverse mechanical loads.8 There is some error associated with J when the periosteal mould method is applied, because of the exclusion of medullary cavity dimensions. This inaccuracy limits the comparability of J calculated using this method to values of J reported in other studies that have generated this value from periosteal and endosteal information.22,24 Values of J calculated from periosteal contours alone, however, correlate highly with values of J calculated from CT scans or direct sectioning.23,25 Periosteally derived J values can therefore be used to compare long bone strength characteristics within a sample. Humerus bilateral asymmetry (BA) in TA and J were quantified as percentage values, as was sexual dimorphism (Table 2).
Diaphyseal shape was assessed by quantifying the ratio of maximum to minimum bending strengths (Imax/Imin) as a measure of diaphyseal circularity. Imax/Imin is a robust indicator of diaphyseal shape, as it compares two non-fixed axes.8 A value of Imax/Imin that is close to unity represents a circular diaphysis, in which uniform loading has occurred around the long axis of the diaphysis, while values greater than one indicate greater loading in the direction of the Imax value.
Different body size standardisation methods were applied to humerus and femur CSG properties (Table 2). While the upper limbs are not weight bearing, humerus CSG values must be scaled to account for differences in bone lengths. Humerus CSG properties were standardised to body size using theoretical equivalents for the product of bone length and body mass based on bone length alone. The powers of 3 for standardising cross-sectional areas and 5.33 for torsional strength were used as they provide a strong correlation between humerus length and body size in Homo sapiens.30
Femur CSG properties must be standardised in a way that incorporates estimated body mass, as mass affects axial, bending and torsional loading in weight-bearing long bones.30 TA and J were standardised to body mass using formulae incorporating body mass and bone length.11,22 Estimated body mass was calculated using the McHenry31 formula in which femur head diameter is used (Table 2). This formula is preferred for assessing body mass in small-bodied samples, like LSA foragers.32,33
To determine the accuracy of the periosteal mould method, data from the Stock and Pfeiffer7 study were compared to data from the present study.
One of the authors (MEC) scanned all periosteal moulds, both those retained from Stock and Pfeiffer's7 research and those that were newly obtained. CSG properties from the retained moulds, calculated by MEC, were compared to the original measurements on file. This calibration exercise assures consistency between the studies. Imax/Imin and J were compared between individuals assessed in both studies using two-tailed paired t-tests. Results from the Stock and Pfeiffer7 study were also regressed using a reduced major axis linear regression to determine the correlation between data obtained from the same long bones using the two methods. After establishing homoscedasticity between samples using Levene's test for the equality of variances, independent samples t-tests (Student's t-tests) were used to compare TA, J and Imax/Imin between men and women within each region for both humeri and femora. To investigate ecological variation, samples from the fynbos, forest and inland regions were compared using one-way ANOVAs. For all statistical comparisons, α was set at 0.05 for detecting significant differences between samples.
Results
Comparison between methods
A strong correlation exists between data from Stock and Pfeiffer7 and data obtained from the same long bones using solely periosteal contours. Both Imax/Imin (r2= 0.82) and JA (r2= 0.91) correlate strongly. When compared using a two-tailed paired samples t-test, there are no significant differences between the Imax/Imin (p=0.174) or J (p=0.972) values obtained using the different methods. This result validates further comparison of CSG properties calculated using periosteal contours, allowing expansion of the sample sizes from the fynbos and forest regions.
Linear dimensions of Later Stone Age adult skeletons
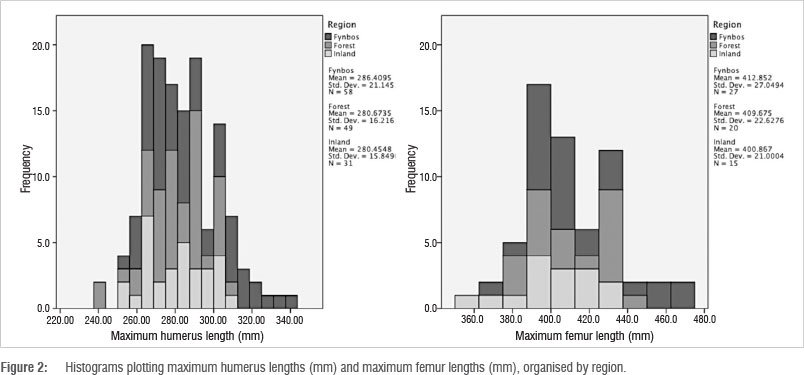
When compared using a one-way ANOVA, no significant differences are evident among the inland, fynbos and forest samples for maximum humerus lengths (p=0.189) and maximum femur lengths (p=0.281) (Figure 2). The samples are homogenous in terms of linear dimensions.
The inclusion of the values from the inland sample expands the prior observation that LSA forager adults are small bodied.34
Humerus cross-sectional geometric properties
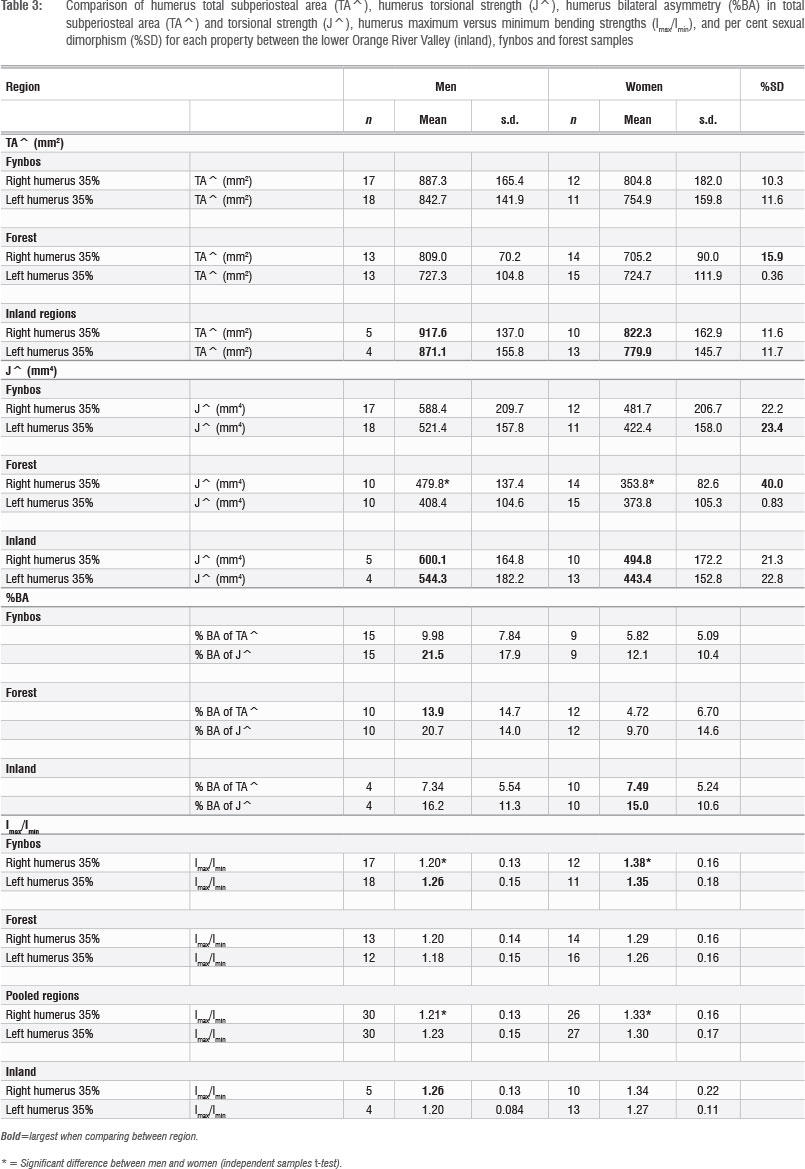
In absolute terms, inland men and women display the highest humerus total area (TA^) values (Table 3). In all three regions, men display higher absolute TA^ humerus values than women but the differences do not reach significance in any region. Comparing the magnitude of sexual dimorphism by region, it is greatest between the right humeri of the forest men and women. The magnitude of dimorphism is lower and is comparable for inland and fynbos men and women. Sexual dimorphism is the lowest between the left humeri of forest men and women.
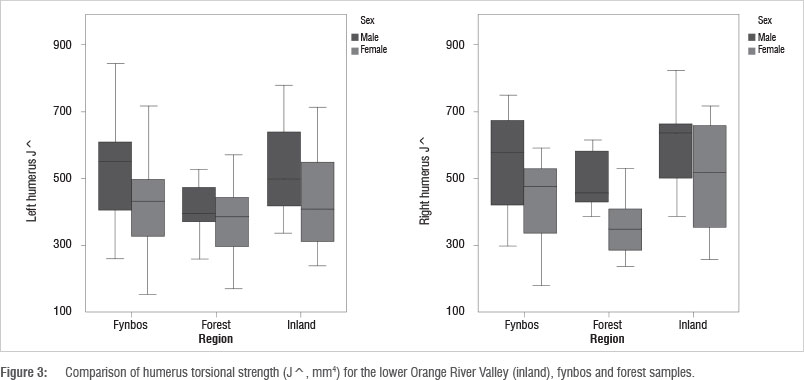
With regard to humerus torsional strength, men show higher J^ than women, and in terms of regions, inland men and women display the highest J^ values (Table 3; Figure 3). There is a significant difference between the right humeri of forest men and women (p=0.00138). Sexual dimorphism is again greatest between the right humeri of forest men and women. Values are lower and comparable for the fynbos and inland men and women, except for left humeri J^, for which forest men and women display the lowest sexual dimorphism. These results suggest that fynbos and inland men and women experienced similar patterns in diaphyseal loading intensity, different from those in the forest.
TA^ and J^ bilateral asymmetry values are similar for inland men and women. This result differs from the fynbos and forest regions, where men and women display different bilateral asymmetry values (Table 3; Figure 4). The difference between the sexes in bilateral asymmetry is trending towards significance for the forest men and women (p=0.089 for TA^ and p=0.068 for J^). Inland women display greater bilateral asymmetry than women from the other two regions, and inland men display lower bilateral asymmetry than other men.
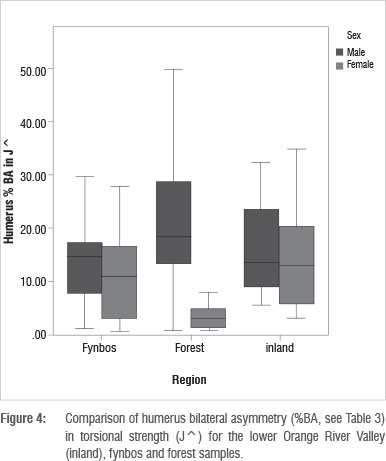
There is a significant difference in the circularity (Imax/Imin) of the right humeri between men and women in the fynbos (p=0.003; Table 3). The difference between the right humeri of men and women when the forest and fynbos samples are pooled is also significant (p=0.002). There are no significant differences between inland men and women for either side. This suggests less differentiation between inland men and women in the distribution of diaphyseal loading, compared to the other regions.
Femur cross-sectional geometric properties
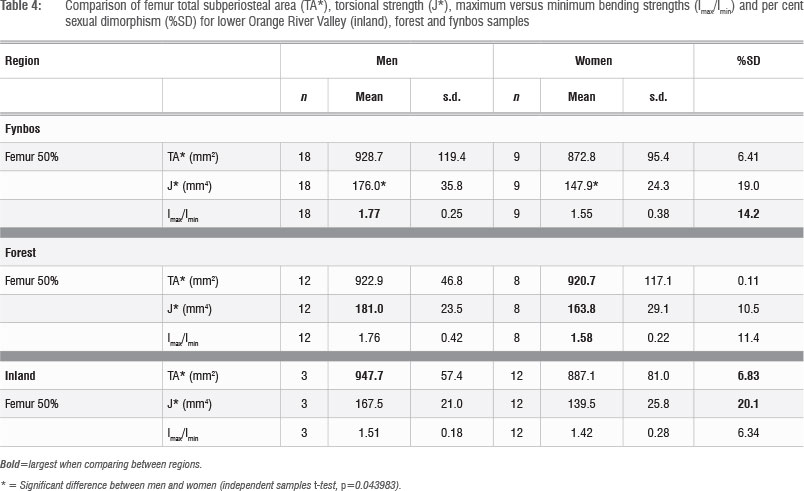
Table 4 documents femur total area (TA*) for all regions. The TA* values of men are higher than women in all regions; however, there are no significant differences between the sexes. The highest male TA* values are found among inland men, while the highest female values are evident among forest women. The degree of sexual dimorphism between men and women is comparable for the inland and fynbos samples, and is lower between forest men and women.
Table 4 and Figure 5 display femur torsional strength (J*) values for all regions. Men have higher J* than women in all regions. Male J* values are similar regardless of regional origin. Forest men and women display the highest J* values when compared to the other regions. Sexual dimorphism is comparably high between the J* values of inland and fynbos men and women: the difference between fynbos male and female J* values reaches statistical significance (p=0.044). Sexual dimorphism is relatively low between forest men and women, because forest women have relatively high J* values when compared to those of inland and fynbos women. These results suggest that forest male and female femora were loaded with relatively similar intensities, while inland and fynbos male femora were loaded at higher intensities than female femora.
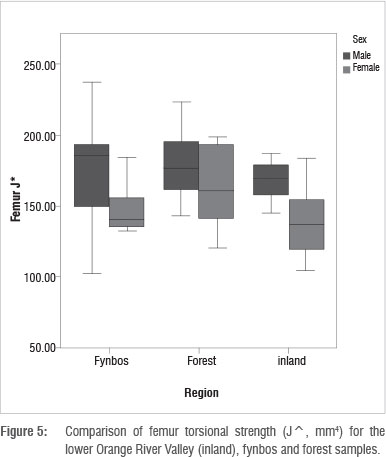
The femur circularity (Imax/Imin) results are displayed in Table 4. Men have higher Imax/Imin (less circular femora) than women in all regions. The Imax/Imin values are comparable between the fynbos and forest samples: men of these two regions have very similar Imax/Imin, as do women. Fynbos men have the highest Imax/Imin (the greatest deviation from circularity) when compared to men from the other regions, while forest women have the highest female Imax/Imin values. Inland men and women, however, display slightly lower values than the other two regions. The inland sample also displays less sexual dimorphism.
Comparisons to foragers from other parts of the world
Because of the somewhat different methods used for body size standardisation and biomechanical strength assessment, only values for humerus and femur Imax/Imin can be readily compared with published values from foragers who lived in other parts of the world. This variable is relatively resistant to observer error and methodological differences, and it reflects bone adaptation to habitual loading.23 Imax /Imin does not need to be standardised based on body size, thereby simplifying comparisons. Australian Aborigine humerus CSG properties could not be compared to the LSA data, as Carlson et al.12 took all measurements at the humerus mid-shaft (50%) rather than mid-distally (35%).
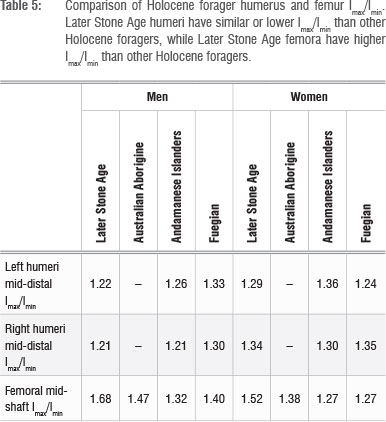
The patterns reported in previous comparisons of LSA CSG properties have been maintained (Table 5). LSA foragers display average to low humerus Imax/Imin when compared to Andaman Islanders and Yahgan foragers, and high femur Imax/Imin when compared to these two samples as well as Australian Aborigines. High femur Imax/Imin values in LSA foragers relative to other mobile forager samples confirm the prior observation that terrestrial mobility was high amongst this group relative to other foraging populations.
Discussion
LSA foragers represent a good sample for this type of analysis, as there is good preservation of discrete individual skeletons. By including skeletons from three regions of South Africa, this work includes a diverse sample that holistically represents active foragers and herder-foragers in South Africa across the past 10 000 years. Achieving humerus CSG property values that correlate well to those of Stock and Pfeiffer7 using the periosteal mould method validates the use of this new, simpler technique. Although there are some differences among and between the three South African regions compared, there is also evidence of behavioural homogeneity within the LSA.
The upper limb
The humerus torsional strength values of forest men and women suggest that forest men pursued activities that relied heavily on their right arms, while the forest women did not. Fynbos men and women also undertook different types of activities from one another. In both scenarios, these observations may be related to the foraging activities of women, including their bilateral use of digging sticks, and the hunting activities of men, which may be more lateralised. The observation that only two humerus measures in men and women differed significantly among the three samples supports the assertion that there is low variation in the activities these three populations undertook using their upper limbs.
Upper limb strength is slightly higher among inland men and women, which suggests that higher loads may have been placed on the diaphyses of this sample. While pastoralism was hypothesised to reduce labour intensity, the opposite effect was observed. Higher intensity workloads may have been associated with the challenge of acquiring resources in the semi-arid environment of the lower Orange River Valley. Herder-foragers may also have increased resource acquisition to cope with competition from non-foraging groups who were also occupying this region. If herder-foragers in the lower Orange River Valley were supplementing the resources of non-foraging groups with foraged foods - a dynamic suggested by Wadley35 based on observations at the Jubilee Shelter in Magaliesburg - herder-foragers may have intensified resource collection to meet these demands.
Inland male and female bilateral asymmetry and diaphyseal shape values are similar, suggesting that men and women participated in similar types of activities. These results differ from those of the forest and fynbos regions, where differences in strength asymmetry and shape were previously attributed to the different activities associated with hunting for men and foraging for women.7 The convergence in bilateral asymmetry and diaphyseal shape between the sexes in the inland sample suggests that men and women from the lower Orange River Valley participated in activities that were more similar than those of foragers from the fynbos and forest. However, the inland sample is small.
The lower limb
The greater lower limb strength values observed in men relative to women are consistent with values reported in previous studies. Lower limb strength is comparable between the men of all three regions, which is unexpected given the differences in terrain: in terms of topographic relief, the forest is the most mountainous, followed by the fynbos, then inland.7 This result suggests that fynbos and inland men must have been very mobile and traversed considerable distances to have acquired similar lower limb strength values as men from the more rugged forest region.
Forest women display femora that are more robust than those of inland and fynbos women. This suggests that they may have been more terrestrially mobile than their fynbos and Karoo counterparts. These results indicate a lower degree of sex-based differentiation in terrestrial mobility in the forest during the LSA as compared to other regions of the southern African Cape. The shared pattern of higher sexual dimorphism in femoral CSG properties between men and women of inland and fynbos regions suggests that a similar sex-based differentiation of terrestrial mobility may have occurred in these two regions. This may reflect a similar division of labour between men and women in terms of mobility. However, limited conclusions can be drawn because of the small sample sizes.
Summary and conclusions
In the inland, forest and fynbos regions, men participated in activities that increased humerus and femur biomechanical loading as compared to women, regardless of local ecology. Women's upper arm bilateral symmetry and shaft non-circularity in the forest and fynbos regions suggests that women participated in low-intensity activities with non-equivalent loading around the shaft. In contrast, men and women of the lower Orange River Valley engaged in activities with heavier upper arm biomechanical loading.
Relatively little behavioural variability is evident among foragers and herder-foragers from the fynbos, forest and inland regions of South Africa, despite the ecological differences of these regions and the probable incorporation of some pastoralism among the inland foragers. The lack of statistically significant differences between the upper and lower limb strength and shape characteristics of these three samples suggests that many similar physical behaviours were commonly undertaken. The inland forager-herder sample from a semi-arid region displays CSG property values that are similar to LSA foragers who did not possess livestock and lived in more resource-rich environments.
The interpretation of results from this study is limited by the small size of the inland herder-forager sample. As the lower Orange River Valley individuals have long bone lengths that are similar to those of the smallbodied foragers from the fynbos and forest, pastoralism does not appear to have affected body size. However, further investigation is needed to explore possible relationships among foraging, pastoralism and body size. As there appear to be meaningful patterns in the CSG properties of these individuals, further investigation into the inland population is warranted. As interaction with non-foraging groups may have also affected the habitual activities of inland populations, the CSG properties of individuals from neighbouring, non-LSA groups could also be examined. This examination would help clarify if inland herder-foragers displayed slightly different manual activities from non-foraging populations. Further investigation of local resource availability, geography, hydrology, and topography in each region could clarify the relationships between observed patterns in terrestrial mobility and these factors.
The similarity between the CSG properties of the three samples supports an assumption of behavioural and physical activity homogeneity among foraging populations, regardless of ecological variation, or the incorporation of pastoralism. These results are generally consistent with those from previous investigations into the physical activities of LSA foragers. They reinforce a growing impression that 'trekking' was a core adaptation that was particularly important among hunter-gatherers of southernmost Africa.
Acknowledgements
We thank L. Elizabeth Doyle and Catherine Merritt for collecting the diaphyseal moulds used in this study and Professor Michael Schillaci for his comments. We also thank the curating institutions for permission to study their collections (National Museum Bloemfontein, Iziko Museums of South Africa and McGregor Museum). In all instances, curatorial procedures of the respective institutions were followed. No permits were required for the described study, which complied with all relevant institutional regulations. The Social Sciences and Humanities Research Council provided funding for this research.
Authors' contributions
S.P arranged for the observations to be made at various curation facilities in South Africa and was responsible for the broad concept on which the research is based; M.E.C. organised and analysed the data; and both authors wrote the manuscript.
References
1. Morris AG. A master catalogue: Holocene human skeletons from South Africa. Johannesburg: Witwatersrand University Press; 1992. [ Links ]
2. Barham L, Mitchell P The first Africans: African archaeology from the earliest Toolmakers to most recent foragers. New York: Cambridge University Press; 2008. [ Links ]
3. Deacon HJ, Deacon J. Human beginnings in South Africa: Uncovering the secrets of the Stone Age. Walnut Creek, CA: Altamira Press; 1999. [ Links ]
4. Smith P Horwitz LK, Kaplan E. Skeletal evidence for population change in the late holocene of the south-western Cape: A radiological study. S Afr Archaeol Bull. 1992;47(156):82-88. http://dx.doi.org/10.2307/3889202 [ Links ]
5. Churchill SE, Morris AG. Muscle marking morphology and labour intensity in prehistoric Khoisan foragers. Int J Osteoarchaeol. 1998;8(5):390-411. http://dx.doi.org/10.1002/(SICI)1099-1212(1998090)8:5<390::AID-OA435>3.0.CO;2-N [ Links ]
6. Ledger M, Holtzhausen LM, Constant D, Morris AG. Biomechanical beam analysis of long bones from a late 18th century slave cemetery in Cape Town, South Africa. Am J Phys Anthropol. 2000;112(2):207-216. http://dx.doi.org/10.1002/(SICI)1096-8644(2000)112:2<2ü7::AID-AJPA7>3.0.CO;2-K [ Links ]
7. Stock J, Pfeiffer S. Long bone robusticity and subsistence behaviour among Later Stone Age foragers of the forest and fynbos biomes of South Africa. J Archaeol Sci. 2004;31 (7):999-1013. http://dx.doi.org/10.1016/j.jas.2003.12.012 [ Links ]
8. Ruff CB. Biomechanical analyses of archaeological human skeletons. In: Katzenberg MA, Saunders SR, editors. Biological anthropology of the human skeleton. 2nd ed. New York: Wiley-Liss, Inc.; 2008. http://dx.doi.org/10.1002/9780470245842.ch6 [ Links ]
9. Trinkaus E, Churchill SE, Ruff CB. Postcranial robusticity in Homo. II : Humeral bilateral asymmetry and bone plasticity. Am J Phys Anthropol. 1994;93:1-34.http://dx.doi.org/10.1002/ajpa.1330930102 [ Links ]
10. Schmitt D, Churchill SE, Hylander WL. Experimental Evidence concerning spear use in Neandertals and early modern humans. J Archaeol Sci. 2003;30(1):103-114. http://dx.doi.org/10.1006/jasc.2001.0814 [ Links ]
11. Stock JT. Climatic and behavioural influences on postcranial robusticity among Holocene foragers. Toronto: University of Toronto; 2002. [ Links ]
12. Carlson KJ, Grine FE, Pearson OM. Robusticity and sexual dimorphism in the postcranium of modern hunter-gatherers from Australia. 2007;23(May):9-23. [ Links ]
13. Dean WR, Milton S. The Karoo: Ecological patterns and processes. New York: Cambridge University Press; 1999. [ Links ]
14. Morris AG. The skeletons of contact: A study of protohistoric burials from the lower Orange River Valley, South Africa. Johannesburg, South Africa: Witwatersrand University Press; 1992. [ Links ]
15. Maggs TMO. Pastoral settlements on the Riet River. S Afr Archaeol Bull. 1971;26(101/102):37-63. http://dx.doi.org/10.2307/3888527 [ Links ]
16. Smith AB. Keeping people on the periphery : The ideology of social hierarchies between hunters and herders. J Anthropol Archaeol. 1998;17:201-15. http://dx.doi.org/10.1006/jaar.1998.0323 [ Links ]
17. Sadr K, Smith A, Plug I, Orton J, Mutti B. Herders and foragers on Kasteelberg: Interim report of excavations 1992-2002. S Afr Archaeol Bull. 2003;58(177):27-32. http://dx.doi.org/10.2307/3889154 [ Links ]
18. Sadr K, Plug INA. Faunal remains in the transition from hunting to herding in southeastern Botswana. S Afr Archaeol Bull. 2001;56(173/174):76-82. http://dx.doi.org/10.2307/3889030 [ Links ]
19. Ruff C, Holt B, Trinkaus E. Who's afraid of the big bad Wolff ?: "Wolff's Law" and bone functional adaptation. Am J Phys Anthropol. 2006;129(4):484-498. http://dx.doi.org/10.1002/ajpa.20371 [ Links ]
20. Stock J, Pfeiffer S. Linking structural variability in long bone diaphyses to habitual behaviors : Foragers from the southern African Later Stone Age and the Andaman Islands. Am J Phys Anthropol. 2001;115(May):337-348. http://dx.doi.org/10.1002/ajpa.1090 [ Links ]
21. Ogilvie MD, Hilton CE. Cross-sectional geometry in the humeri of foragers and farmers from the prehispanic American Southwest: Exploring patterns in the sexual division of labor. Am J Phys Anthropol. 2011;144:11-21. http://dx.doi.org/10.1002/ajpa.21362 [ Links ]
22. Nikita E, Siew YY Stock J, Mattingly D, Lahr MM. Activity patterns in the Sahara Desert: An interpretation based on cross-sectional geometric properties. Am J Phys Anthropol. 2011 ;146(3):423-434. http://dx.doi.org/10.1002/ajpa.21597 [ Links ]
23. Stock JT, Shaw CN. Which measures of diaphyseal robusticity are robust? A comparison of external methods of quantifying the strength of long bone diaphyses to cross-sectional geometric properties. Am J Phys Anthropol. 2007;134(3):412-423. http://dx.doi.org/10.1002/ajpa.20686 [ Links ]
24. Sparacello VS, Pearson OM. The importance of accounting for the area of the medullary cavity in cross-sectional geometry: A test based on the femoral midshaft. Am J Phys Anthropol. 2010;143(4):612-624. http://dx.doi.org/10.1002/ajpa.21361 [ Links ]
25. Macintosh AA, Davies TG, Ryan TM, Shaw CN, Stock JT. Periosteal versus true cross-sectional geometry: A comparison along humeral, femoral, and tibial diaphyses. Am J Phys Anthropol. 2013;150(3):442-452. http://dx.doi.org/10.1002/ajpa.22218 [ Links ]
26. Davies TG, Shaw CN, Stock JT. A test of a new method and software for the rapid estimation of cross-sectional geometric properties of long bone diaphyses from 3D laser surface scans. Archaeol Anthropol Sci. 2012;4(4):277-290. http://dx.doi.org/10.1007/s12520-012-0101-8 [ Links ]
27. Stynder DD. Craniometric evidence for South African Later Stone Age herders and hunter-gatherers being a single biological population. J Archaeol Sci. 2009;36(3):798-806. http://dx.doi.org/10.1016/j.jas.2008.11.001 [ Links ]
28. Rasband WS. Image J. Bethesda, MD: US National Institutes of Health. [ Links ]
29. Ruff CB. MomentMacro for NIH Image and Image J. 2006. http://dx.doi.org/10.1002/ajpa.1330910103 [ Links ]
30. Ruff CB, Trinkaus E, Walker A, Larsen CS. Postcranial robusticity in Homo. I : Temporal trends and mechanical interpretation. Am J Phys Anthropol. 1993;91(1):21-53. [ Links ]
31. McHenry HM. Body size and proportions in early hominids. Am J Phys Anthropol. 1992;87:407-431. http://dx.doi.org/10.1002/ajpa.1330870404 [ Links ]
32. Auerbach BM, Ruff CB. Human body mass estimation: A comparison of "morphometric" and "mechanical" methods. Am J Phys Anthropol. 2004;125(4):331-342. http://dx.doi.org/10.1002/ajpa.20032 [ Links ]
33. Kurki HK, Ginter JK, Stock JT, Pfeiffer S. Body size estimation of smallbodied humans: Applicability of current methods. Am J Phys Anthropol. 2010;141(2):169-180. [ Links ]
34. Sealy J, Pfeiffer S. Diet, body size, and landscape use among holocene people in the southern Cape, South Africa. Curr Anthropol. 2000;41(4):642-655. http://dx.doi.org/10.1086/317392 [ Links ]
35. Wadley L. Changes in the social relations of precolonial hunter-gatherers after agropastoralist contact: An example from the Magaliesberg, South Africa. J Anthropol Archaeol. 1996;15:205-217. http://dx.doi.org/10.1006/jaar.1996.0008 [ Links ]
 Correspondence:
Correspondence:
Michelle Cameron
Department of Archaeology and Anthropology
University of Cambridge
Pembroke Street, CB2 3QG, United Kingdom
EMAIL: mec62@cam.ac.uk
Received: 05 Dec. 2013
Revised: 02 Mar. 2014
Accepted: 04 Mar. 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}