










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.110 no.7-8 Pretoria ago. 2014
RESEARCH ARTICLE
Craniodental continuity and change between Iron Age peoples and their descendants
Kerryn A. Warren; Simon Hall; Rebecca R. Ackermann
Department of Archaeology, University of Cape Town, Cape Town, South Africa
ABSTRACT
The appearance of the Iron Age of southern Africa early in the first millennium AD is associated with the migration of Bantu speakers who were broadly ancestral to present-day Bantu speakers. While there is sufficient genetic, physical anthropological and cultural evidence to support general continuity into contemporary populations, the extent to which events since colonialism have affected morphological variation is poorly understood. We used dental anthropological techniques and three-dimensional craniomandibular metrics to examine biological relationships among Iron Age farmers, a historical 19th-century Ndebele sample and 20th-century Bantu speakers. We show that, although Iron Age and modern morphologies are generally similar, there are differences. Moreover, the historical sample falls between the precolonial and modern samples, suggesting increased genetic exchange from the 19th century onwards. These results suggest that recent historical events altered the genetic make-up of Bantu speakers and that, as a result, extrapolations from modern groups to the past should be done with caution as morphological variability is relative to historical context.
Keywords: dental anthropology; southern Africa; Bantu speakers; admixture; Historic Cave
Introduction
In southern Africa over the last 2000 years, there have been several significant demographic shifts that imply gene flow or exchange at several different geographical scales. Early in the first millennium AD, the Iron Age appears rapidly in this region and is identified archaeologically through a 'package' of agropastoralist elements associated with permanent and semi-permanent settlements.1-4 These elements include a distinctive ceramic style (the Chifumbaze complex) that is widespread in East and southern Africa, and metallurgical skill. The archaeological and linguistic evidence indicates that the abrupt introduction of this 'Iron Age package' is associated with the migration of farmers who spoke Bantu languages and that there is a general continuity through to Bantu speakers in present-day southern Africa. This scenario is also supported by genetic evidence from modern-day descendants, which indicates recent and common origins throughout much of southernmost Africa.5-8
Debate concerning the cultural continuity between the Early Iron Age (EIA) and historical Bantu speakers5,9,10 considers that EIA people did not arrive and develop in isolation; the evidence for intermarriage and cultural exchange with Stone Age hunter-gatherers is clear.5,11 Additionally, the shift in ceramic style between the EIA and Later Iron Age (LIA) early in the second millennium AD12 is associated with further migration from east Africa. Blackburn ceramics (ancestral Nguni speakers) appear in KwaZulu-Natal from around 1100 AD and Moloko ceramics (ancestral Sotho-Tswana speakers) appear north of the Soutpansberg from around 1300 AD.12-15 Furthermore, Sotho-Tswana and Nguni ancestors colonised the southern grasslands, south of the Vaal River16, and the Eastern Cape from the 16th century12. Once again, linguistic, cultural and genetic evidence shows considerable interaction with San hunter-gatherers. Significant shifts among agropastoralists, especially Nguni diasporas from northern KwaZulu-Natal from the 16th century AD, may have been prompted by climatic change and the introduction of maize via the Portuguese.12
Demographic movement increased from the early 19th century as colonial expansion intensified from the southeast African coast and from the Cape. This expansion contributed to the early 19th-century mfecane/difaqane (troubled times) during which there was considerable demographic movement and change.12 Colonial encroachment and intensified trade demands for ivory and labour contributed to agropastoralist political centralisation in the west, the development of large Tswana towns17 and the establishment of the Zulu state in northern KwaZulu-Natal18,19. These events exacerbated a demographic 'swirl' and Nguni speakers, for example, moved into southwestern Zimbabwe as well as northern Malawi and southern Tanzania. Despite the colonial characterisation of this period as savage and chaotic, the movement of people and the negotiation of new political arrangements elsewhere simply continued from previous political processes that underpinned inclusiveness. The Land Act of 1913 and the establishment of apartheid homelands during the 20th century collapsed identity into an immutable package of race, culture and language.
While the historical and archaeological record indicates overall continuity through the EIA and into the present there has been gene flow from both closely related groups and more distantly related ones (e.g. Khoesan, colonists) and this is likely to have influenced the biological composition of these peoples over time.8,20 Although studies of the remains of Iron Age peoples have the potential to contribute to our understanding of biological continuity (or change) over time, to date the contribution of physical anthropology to the question of temporal continuity is limited. Analyses of morphological variability within and between Iron Age human samples, as well as between Iron Age and later samples, are rare (but see Ribot et al.21). Because Iron Age human burials are typically dispersed within a settlement (in a patterned way according to gender and status), it is unlikely that more than a few burials are discovered and excavated per site.22 The exceptions are the burials excavated from the early second millennium AD Mapungubwe and K2 capitals, which have been analysed and shown to be within the range of variation expected in modern Bantu speakers.22-25 These specimens have since been reburied.26
Additionally, a number of studies have looked at skeletal variation in modern Bantu-speaking peoples. A large-scale study on variability between Bantu speakers, using linguistic identity as a proxy for 'tribal' affiliation was conducted by De Villiers27, based on cranial metric and non-metric analyses27. Her study showed that there is low variation between recent historical South African Bantu speakers and that many features were similar between these groups and the Khoesan (suggesting admixture). This finding was supported by a more recent study using geometric morphometrics.28 Dental anthropology on modern Bantu groups conducted by Jacobson29 and Irish30,30 also support conclusions of low inter-population variability2931. These conclusions are supported by the genetics32, although later research suggests that the level of admixture varies between groups, indicating that differences between southern Bantu and more northern populations result from the intensity of interaction and intermarriage with Khoesan28,33.
In this study, we compared modern and archaeological Iron Age human samples, as well as an historical Ndebele sample, in order to assess the degree of morphological continuity among these broad groups. These analyses focus on cranial and dental morphology for two reasons. Firstly, in other studies on both contemporary and archaeological human samples,23,28,33-40 both dental and cranial morphology have been shown to be population-specific indicators of identity. Secondly, teeth are better preserved in the archaeological record than the postcranial skeleton and therefore comprise a larger sample size. Our main objective in this study was to characterise cranial and dental variability within Iron Age and more contemporary Bantu speakers, and identify differences in variation that may have arisen as a result of recent historical processes influencing gene flow. As within-group variability in both the Iron Age and modern Bantu speakers has been shown to be small,27,29,30,38,41 we considered them as cohesive entities. Significant differences between these larger temporal groups is likely to have arisen during the colonial and post-colonial periods, represented here (albeit incompletely) by the historical Ndebele (Historic Cave) sample. More broadly, this research will allow us to evaluate the assumptions that can be made of historical groups using contemporary Bantu-speaking populations.
Materials and Methods
Specimens
The sample is separated into three groups: Iron Age, historical and modern. The Iron Age specimens (n=142) come from a number of sites and are housed in the following institutions: University of Cape Town, the Natal Museum, the University of the Witwatersrand, the University of Pretoria, the National Museum of Cultural History (Ditsong Museums of South Africa) and the University of Botswana (in association with the Botswana National Museum). Sites from where the samples originate range from as far north as Zambia (Ingombe Ilede and Isamu Patu), east into the Kalahari of eastern Botswana (Toutswemogala sites) and along the KwaZulu-Natal (eastern) coast of South Africa. All these sites date to between 1600 and 150 BP and consequently straddle both the EIA and LIA. Because current research suggests that morphological variability (both temporal and geographical) between groups in the Iron Age is low,41 the Iron Age is treated as a single group for comparisons within this study. The historical specimens are Ndebele peoples from Historic Cave, located in the Makapan Valley in South Africa, and date to a historically documented siege by the Trekboers in 1854.18 The sample consists mostly of loose teeth (166 teeth in total). While the Historic Cave sample is clearly not representative of the entire colonial period, it does provide a useful and interesting marker with which to compare the modern and Iron Age samples.
Data for the modern sample were collected from the Raymond A. Dart Collection of human skeletons at the University of the Witwatersrand. The full sample consists of 39 individuals derived from southern Bantu-speaking groups. These specimens are classified as Zulu, Sotho and Xhosa in the catalogues; however, it is important to remember that many of the specimens within the Raymond A. Dart Collection of human skeletons have been classified into these groups on the basis of language, and more specifically surnames,42 so these affiliations may not be entirely accurate. Details for the list of specimens, including descriptions of individual specimens (Iron Age and modern) can be found in Warren41. Sex was not taken into account in the analyses as a large portion of the sample consists of individual teeth and there are currently no reliable metrical methods for determining sex from teeth available for South Africa. Only adult crania and teeth (including adult teeth from subadult individuals) were used for morphological and dental analyses.
Dental anthropology
Both metric and non-metric data were collected to examine dental variability among the Iron Age, historical (19th century) and modern Bantu speakers (cadavers from the Dart Collection). Non-metric dental traits were scored according to the Arizona State University Dental Anthropological System.43 These procedures were calibrated with external researchers (Ms Wendy Black who, herself, calibrated with Dr Joel Irish) to increase accuracy to within 93%. Sample sizes varied for each trait within each group, based on presence of tooth and the visibility of the trait. The modern group ranged from 12 to 39 specimens, the historical sample from 1 to 25 specimens, and the Iron Age sample from 47 to 108 specimens. Two dental calliper measurements - buccolingual and mesiodistal lengths - were also taken on each available tooth. Statistical analyses were performed using t-tests (in Microsoft Excel) conducted between each group for all comparable traits. Chi-squared tests were performed in Microsoft Excel on the non-metric traits in order to compare trait frequencies between groups. Testing multiple hypotheses is problematic by nature. If no corrections are made for multiple comparisons, significant findings may be observed by chance - i.e. it is too easy to make a Type I error. By contrast, if corrections for multiple comparisons are made, power to detect real differences is lost - i.e. it is too easy to make a Type II error. Therefore we report both uncorrected and Bonferroni corrected p-values here, for both the chi-squared and the metric comparisons. The mean measure of divergence (MMD; Freeman Turkey transformation) was also calculated using script for R created by A. Soltysiak. MMD was calculated only between the Iron Age and modern samples; MMDs were not calculated using the Historic Cave sample, given the very small sample sizes for some traits. A principal components analysis (PCA) was performed in Statistica (version 11) in order to further illustrate variability between the modern and Iron Age samples. The t-tests were conducted on the regression scores in Excel. For the PCA analysis, Historic Cave specimens were not included because of small sample sizes.
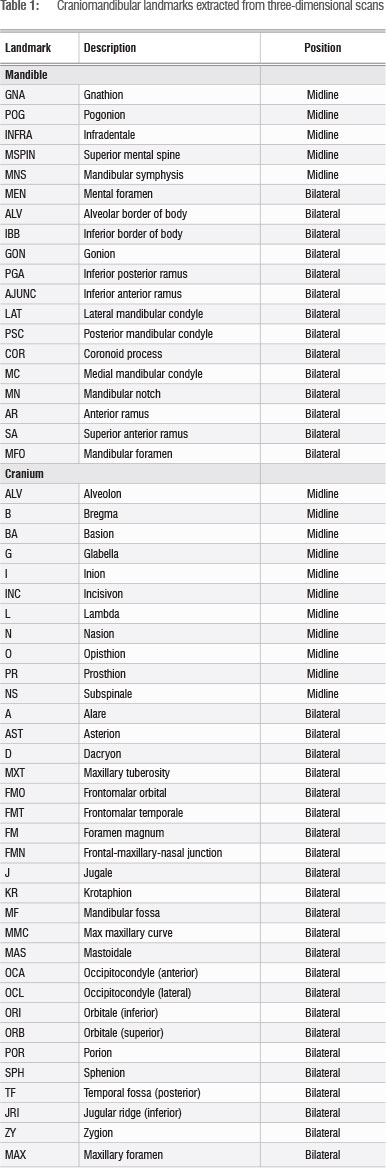
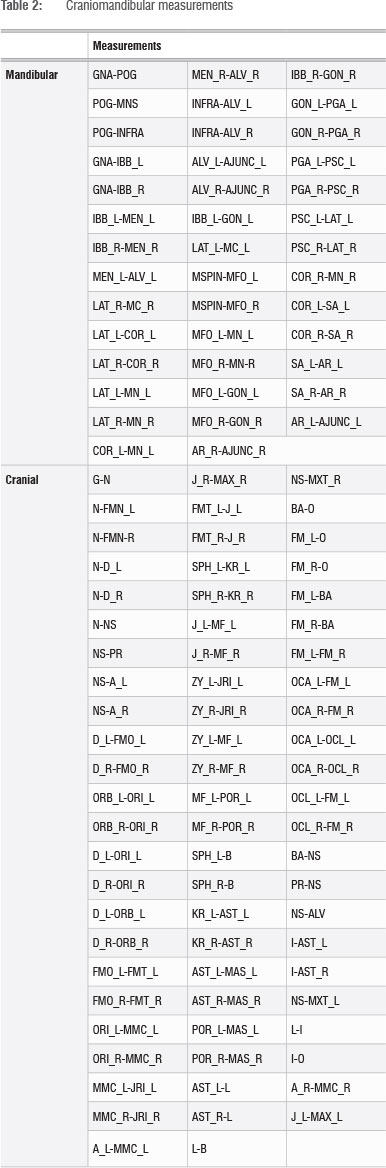
Craniomandibular metrics
A NextEngine 3D Laser Scanner and Scanstudio HD software (version 1.1) were used to create three-dimensional scans of the available cranial and mandibular material. Three-dimensional coordinates for 34 cranial landmarks and 19 mandibular landmarks were then extracted using MeshLab v1.3.1 (Table 1), and a series of Euclidean distances were derived from these landmarks (Table 2). These distances were chosen to capture overall morphology while minimising redundancy. T-tests were performed using Microsoft Excel in order to compare the modern and Iron Age sample; Historic Cave specimens were not included as they were represented by teeth only. Again, we report both uncorrected and Bonferroni corrected p-values for these comparisons. A PCA was performed in Statistica (version 11) to visually display variation, and t-tests were conducted on the regression scores in Excel. Because of missing data, this PCA was by necessity based on only a subset of landmarks.
Results
Dental non-metric results
Table 3 shows the frequencies and sample sizes of the non-metric dental traits for each sample (modern, historical and Iron Age). Table 4 lists the chi-squared p-values calculated for each comparison. Between the Iron Age and the modern samples, 6 out of the 25 traits (24%) are significantly different at p<0.05. These traits are winging (UI1), UM1 cusp 5, Carabelli's trait, protostylid (LM1), torsomolar angle and LM1 root number. Between the Iron Age and historical samples, only three traits show significant differences (12%) at p<0.05. These traits are winging (UI1), canine mesial ridge and LM2 cusp number. Winging is therefore the only trait that is significantly different between Iron Age samples and both the historical and modern samples. However, winging could only be observed on two Historic Cave specimens, the significance of which will be addressed in the discussion. Between the modern and historical samples, two traits (8%) are significantly different at p<0.05: LM1 deflecting wrinkle and canine mesial ridge. Of these, canine mesial ridge was also significantly different for the comparison between the Iron Age and historical material. For the trait deflecting wrinkle the historical sample was once again very small (n=3). Because the chance of calculating at least one significant p-value (0.05) between any two samples is high (72%), we further adjusted the p-value to the Bonferroni correction, at p=0.002. Four traits remain significantly different between the Iron Age and modern samples (Carabelli UM1, protostylid LM1, torsomolar angle and LM1 root number), while canine mesial ridge is still significantly different between the Historic Cave sample and both the Iron Age and modern samples. Although correcting the values confirms that the samples are different, it is nonetheless important to look at each trait (e.g. at p=0.05) to identify the manner in which these samples differ. The MMD (i.e. phenetic distance) between the modern and Iron Age samples is low at 0.088, again suggesting only a small amount of difference between these groups.
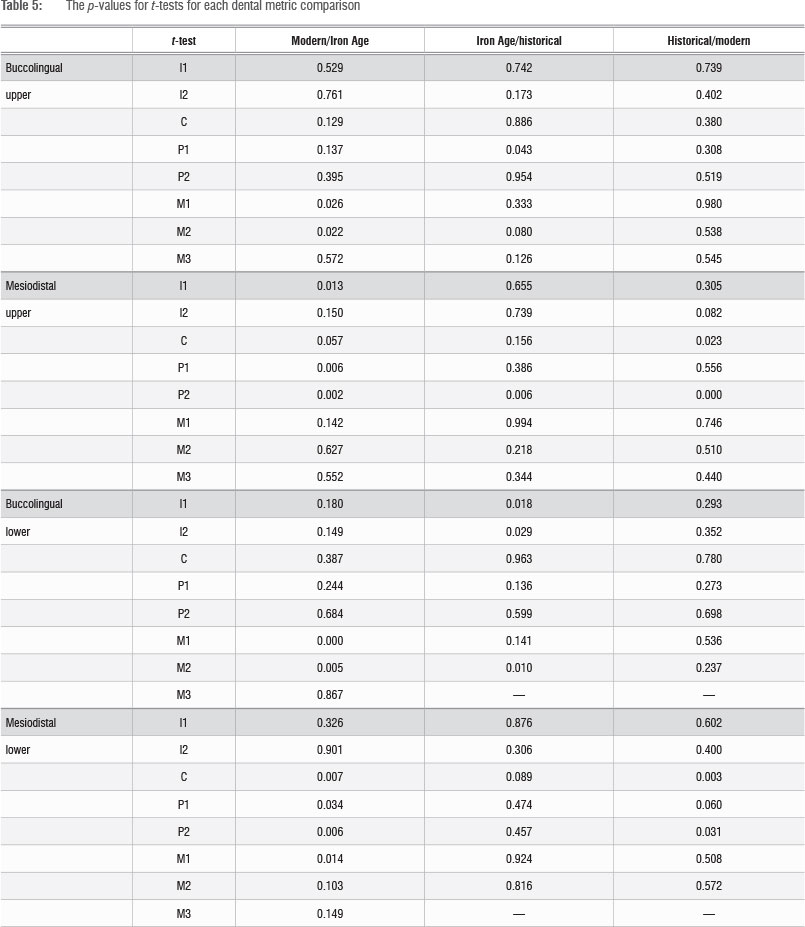
Dental metric results
The 32 dental metric comparisons are shown in Table 5. Between the Iron Age and modern samples, 11 measurements (34%) show significant differences at p<0.05. For buccolingual measurements, the first and second molars (upper and lower) are all significantly different. For mesiodistal length, the premolars (upper P1 and P2, lower P1 and P2), the lower canines, the lower first molar and upper first incisor are significantly different. Between the Iron Age and historical samples, 5 of the 30 measurements (17%) were significantly different at p<0.05. These measurements are buccolingual lengths of the upper first premolar, lower first and second incisors, lower M2, and the mesiodistal length of the upper P2. Of these, mesiodistal length of the upper second premolar and buccolingual length of the lower second molars are also significantly different for the Iron Age versus modern comparison. Between the historical and modern comparisons, four measurements (13%) are significantly different at p<0.05: mesiodistal measures of the upper canine and second premolar and the lower canine and lower second premolar. Of these, mesiodistal measurement of the lower canine and lower second premolar are also significant for the Iron Age versus modern comparison. Mesiodistal length of the upper second premolar is significant for all three comparisons.
Although the use of p=0.05 for significance is useful when comparing individual traits, the chances of calculating one significant p-value when the samples are actually similar is as high as 80%. In order to detect any real difference between the samples themselves (and not just the traits), the p-value was corrected to 0.0016. When this correction was done, only a single measurement differed significantly between the modern and Iron Age samples (buccolingual LM1), and between the Historic Cave and modern samples (mesiodistal UP2). There is no significant difference between the Historic Cave and Iron Age samples. Whether this finding is a result of a small sample size, close biological relationships or a combination of both, is unknown.
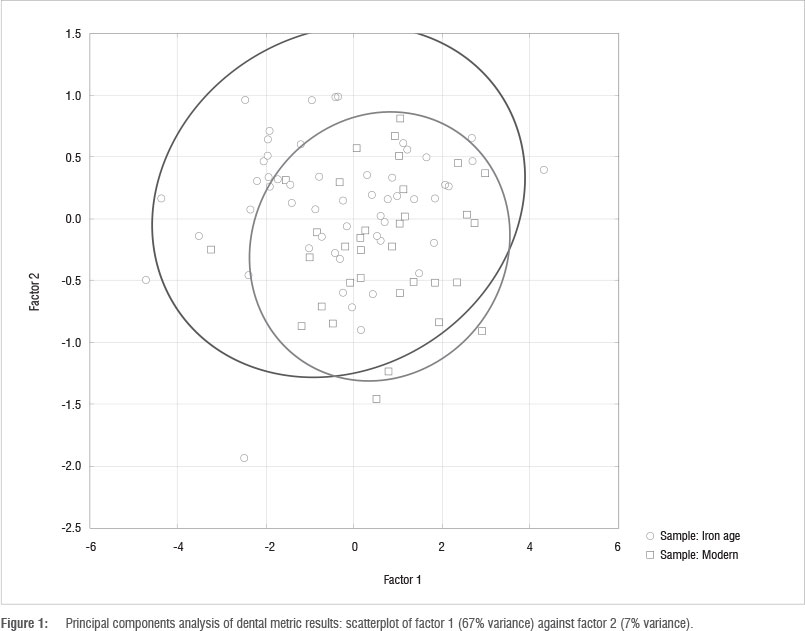
Figure 1 shows the dental metric covariance PCA for factor 1 versus factor 2 for the Iron Age and modern specimens. Coefficient ellipses (95%) surround each group. PC1 shows loadings that are all positive, ranging from only 0.030 (mesiodistal lower first premolar) to 0.179 (buccolingual upper second molar), indicating that PC1 is a size variable. PC1 includes 67% of the variance and PC2 includes 7% of the variance (total 74%). The greatest contributions to the first PC are from the buccolingual upper second molar (0.179 loading proportion) and mesiodistal lower second molar (0.159). The smallest contributions to PC1 come from the buccolingual (0.043) and mesiodistal (0.03) lower first premolars. For PC2, the mesiodistal upper second molar contributes the largest proportion (0.613), with buccolingual and mesiodistal upper first premolars contributing the least (less than 0.01). Figure 1 indicates that although there is much overlap between the modern and Iron Age samples, there are some differences in both size and shape, as indicated by the placement of modern specimens at the lower right of the graph. The t-tests on the regression scores indicate that the two samples are significantly different for PC1, 2 and 6 (size and two shape factors) at p<0.05. At the Bonferroni corrected value of p<0.005, differences for PC2 are still significant, indicating shape differences between the Iron Age and modern samples.
Craniomandibular metric results
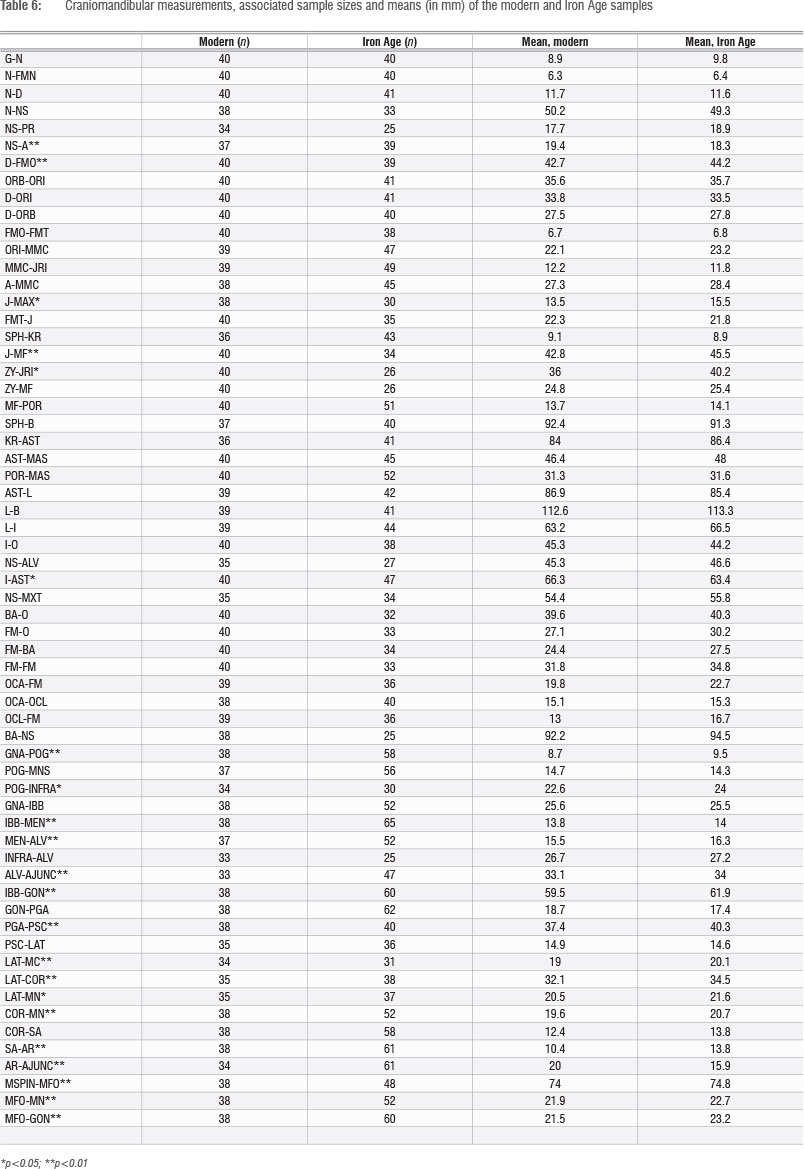
Table 6 shows the results of t-tests between the Iron Age and modern samples for a series of craniomandibular measurements (there were no complete adult cranial or mandibular historical specimens from Historic Cave, only isolated teeth). The table shows the measurements used, sample sizes for each measurement for each category and the mean measurement (in millimetres) for each sample. Sample sizes vary between measurements, depending on preservation and visibility of the landmarks. For the modern sample, each measurement was taken on between 33 and 40 specimens (Table 6). For the Iron Age sample, each measurement was taken on between 25 (NS-PR and INFRA-ALV) and 61 (SA-AR and AR-AJUNC) specimens. Out of the 62 measurements, 22 (35%) are significant at p<0.05 and 17 (27%) are significant at p<0.01. Although only 22 measurements are mandibular, 16 of these measurements (73%) show significant differences (p<0.05).
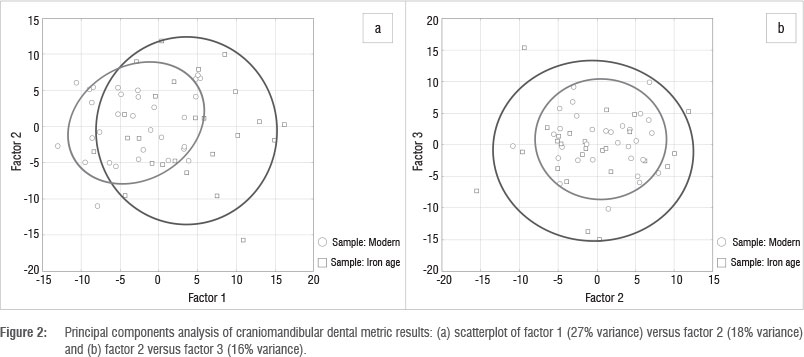
Figure 2 shows the results of the craniomandibular metric PCA. To maximise sample size, only 13 measurements, mostly mandibular, were used in this analysis. The first three PCs account for 27%, 18% and 16% of the variance, respectively (total 61%). The other PCs each contribute less than 10% of the variance. For the first component, mandibular ramus (SA-AR; 0.361) and mandibular foramen to gonion (0.197) contribute the most. Gnathion to pogonion, pogonion to the inferior posterior ramus, coronoid process to mandibular notch and mandibular foramen to poronion measurements contribute the least to PC1 (less than 0.005). For PC2, the inferior border of the body to the gonion contributes the most (0.366) and pogonion to mandibular symphysis, coronoid process to mandibular notch and mandibular foramen to poronion contribute the least (less than 0.005). PC3 has large contributions from gonion to inferior posterior ramus (0.254) and mandibular foramen to gonion (0.296). Figure 2a shows PC1 against PC2 for modern and Iron Age specimens. Although there is much overlap, modern specimens are more concentrated on the left of the graph and Iron Age specimens on the right (i.e. along PC1). Figure 2b (PC2 against PC3) illustrates complete overlap between the Iron Age and modern samples for either PC2 or PC3, although the Iron Age sample does appear to be more variable than the modern one (this pattern is also seen in Figure 2a). The t-tests conducted on the factor scores derived from the craniomandibular measurements of the specimens reveal that PC1 (size) and PC4 show significant difference between the Iron Age and modern samples at p <0.05. PC1 remains significantly different when the Bonferroni p-value is adjusted to 0.004, showing that differences between these groups are statistically meaningful.
Discussion
The results indicate that there is a large amount of phenotypic (and presumably underlying genetic) similarity among the Iron Age, historical and modern samples. This finding is consistent with archaeological research as well as previous genetic and linguistic work that support continuity between precolonial and modern Bantu-speaking peoples and/or cohesion among modern Bantu-speaking peoples.7,20,27-29,32,45 Continuities through the ceramic sequence suggest that ancestral forms of Shona were spoken in the first millennium AD12, and, more specifically, the archaeology shows direct cultural continuity between second millennium AD agropastoralists and historical Sotho-Tswana15 and Nguni speakers13. Despite these overall continuities, significant regional demographic shifts resulted in linguistic and cultural entanglements, and the creation of new identities (e.g. a Venda identity emerged in the 16th century AD from intermarriage between Shona and Sotho speakers46).
It is not surprising that the data in this study indicate homogeneity in the measured traits between the Iron Age and historical samples. The violent nature of the mfecane early in the 19th century led to the dispersal of people, the fragmentation of polities and the reformulation of people into new political units. The small group of Ndebele, estimated to be about 300, who left KwaZulu-Natal under the leadership of Mzilikazi, increased to about 20 000 in the late 1820s - in the early years of the Ndebele state in the present-day Rustenburg area.47 This example underpins that while the initial numbers were low, Mzilikazi 'accumulated' people as his settlement focus shifted further to the west and included a significant number of Sotho-Tswana speakers. While the scale of these demographic shifts in the early 19th century may have been larger, the archaeology of both first and second millennium agropastoralists shows that regional demographic movement was essential to the political process. The examples of the origins of Venda and the earlier precolonial Nguni diasporas underpin demographic and cultural fluidity.12
Despite this overall pattern of similarity among the Iron Age, historical and modern samples, there are some important differences among these groups. Where dental traits do differ (both discrete and metric), the Iron Age sample tends to be more similar to the historical sample than it is to the modern one, suggesting recent, continuous gene flow from other populations in historical and modern times. This increasing gene flow could stem from a number of historical causes. For example, interaction between hunter-gatherers and farmers is observed frequently in the archaeological record,5,12,45,48 and it is possible that admixture occurred throughout this period and may have increased because of more recent historical pressures. This scenario is supported by ongoing research on the dentition of Khoesan people that indicates that there is more similarity between a recent Khoesan sample and modern Bantu speakers than between the Khoesan sample and the Iron Age samples studied here.41 The relationship between the Historic Cave sample and the other samples might reflect increasing population pressures and interactions through time, resulting in them being morphologically intermediate between the Iron Age sample and modern Bantu speakers. This Historic Cave sample comes from the siege of October 1854 when the Kekana (a Ndebele group) took refuge in this large cavern complex in the Makapan valley as a response to a Boer commando seeking revenge for the murder of Trekboers in the previous month.18,49 There was substantial mortality during this siege. It is important to note that while the results do indeed indicate an intermediate position for the Historic Cave material, larger samples (and more sites) are needed in order to better evaluate this interpretation. Further research on the dentition of other colonial period peoples is necessary to advance our understanding of variation and morphological affinities at this time.
In more recent times, our expectation is that admixture increases, which again may explain the continued divergence of the modern samples from the archaeological ones. The metric and non-metric dental comparisons show a greater similarity between the Iron Age and Historic Cave samples than between the modern and Historic Cave samples, supporting increased levels of gene flow into these groups in recent times. The intensification of trade (particularly in ivory and slaves) in the second half of the 18th century and through the 19th century took place in a context of expanding European mercantile interests and both 19th-century historical contexts mentioned above were inextricably linked to this context.18,47 Despite the negative interactions there was also admixture between Bantu speakers and Europeans, which is clearly evident in the modern sample and has also been observed in Mozambican populations.20 This admixture is typically sex biased (European Y-chromosome haplotypes), which is not surprising given that Europeans, specifically the Portuguese, colonised the coast and parts of the interior of Mozambique from the 17th century.50
In addition to offering insight into the morphological effects of admixture as detailed above, this study is also important because it is the first to apply standardised dental anthropology techniques to investigate historical relationships between present-day and archaeological southern African populations. Many dental anthropological studies have focused on modern variation across broad geographical regions, with sub-Saharan Africa representing a distinct dental complex.30,35,36,38,51 Both non-metric and metric dental studies have also focused on variation among non-sub-Saharan-African archaeological samples, indicating continuity, gene flow and morphological change in archaeological samples over time.34,37,39,52-54 Although these studies have successfully addressed questions about variability and genetic identity using archaeological and contemporary samples, little work using these methods has been done in southern Africa. Early research by Shaw described the dentition of modern-day Bantu speakers,55,56 but was unstandardised. Additionally, Jacobson's study which shows there is low variability among modern Bantu speakers, is difficult to compare with more recent standardised dental research.29 Kieser et al.57 evaluated changes in tooth size between living and recent-historical Bantu speakers drawn from the Dart Collection,57 indicating larger mesiodistal and buccolingual lengths within the 19th century. This study, however, is the first to extend dental comparisons into archaeological samples.
Finally, this study has important implications for understanding variation within the modern sample. The research presented here - not unexpectedly - shows a degree of historical admixture in the Raymond A. Dart Collection of human skeletons. But, importantly, it also suggests that it might be possible to use archaeological samples to better understand the biological affinities of these modern specimens using skeletal indicators. This is relevant because there is not necessarily a relationship between the biological history and 'tribal' identities of specimens in the Dart Collection because they are categorised based on surnames.42 Consequently, the biological distinction between the modern and Iron Age samples is a further reminder that modern samples do not provide a comparative baseline for the past.
Conclusion
Three important conclusions can be made from this research. Firstly, there is general similarity between Iron Age farmers, modern Bantu speakers and an historically very specific Ndebele group. This similarity supports a general genetic continuity between precolonial and historical agropastoralists and modern people in southern Africa.
Secondly, despite these similarities, there are more differences between the Iron Age and modern samples than expected, given the seeming homogeneity of both Iron Age and modern Bantu speakers indicated by previous research. This observation demonstrates gene flow between Iron Age descendants and other groups in historical and modern times, and cautions against using modern Bantu speakers as baselines for understanding variation in the past. Finally, the Historic Cave sample appears to be morphologically intermediate between the Iron Age and modern samples, once again supporting the conclusion of increasing gene flow into these groups through time, although small sample sizes reduce the value of this conclusion.
Acknowledgements
This research would not have been possible without the help of the curators and persons from the institutions visited: the University of Cape Town, the Dart Collection in the University of the Witwatersrand, the University of Botswana, the University of Pretoria and the Cultural History Museum (Ditsong Museums). We thank Alan Morris, Lauren Schroeder, Tessa Campbell and Wendy Black for advice. Research was supported by the National Research Foundation of South Africa (R.R.A.) and the Harry Crossley Foundation (K.A.W.).
Authors' contributions
K.A.W. collected the data, analysed samples, performed calculations and wrote the manuscript. R.R.A. conceptualised the project, provided technical and methodological input and helped write the paper. S.H. provided theoretical input and helped write the paper.
References
1. Huffman TN. The Early Iron Age and the spread of the Bantu. S Afr Archaeol Bull. 1970;25(97):3-21. http://dx.doi.org/10.2307/3888762 [ Links ]
2. Huffman TN. Handbook to the Iron Age: the archaeology of pre-colonial farming societies in southern Africa. Pietermaritzburg: University of Natal Press; 2007. [ Links ]
3. Mitchell P The archaeology of southern Africa. Cambridge: Cambridge University Press; 2002. [ Links ]
4. Mitchell P, Whitelaw G. The archaeology of southernmost Africa from c. 2000 BP to the early 1800s: A review of recent research. J Afr Hist. 2005;46(2):1-33. http://dx.doi.org/10.1017/S0021853705000770 [ Links ]
5. Parkington J, Hall S. The appearance of food production in southern Africa 1,000 to 2,000 years ago. In: Hamilton C, Mbenga B, Ross R, editors. The Cambridge history of South Africa. vol. 1. New York: Cambridge University Press; 2010. p. 63-111. [ Links ]
6. Blench R. Archaeology, language, and the African past. Walnut Creek: AltaMira Press; 2006. [ Links ]
7. Greenberg JH. Historical inferences from linguistic research in sub-Saharan Africa. Boston: Boston University Press; 1964. [ Links ]
8. Poloni ES, Semino O, Passarino G, Santachiara-Benerecetti AS, Dupanloup I, Langaney A, et al. Human genetic affinities for Y-chromosome P49a, f/< i> Taq I haplotypes show strong correspondence with linguistics. Am J Hum Genet. 1997;61(5):1015-1035. http://dx.doi.org/10.1086/301602 [ Links ]
9. Huffman TN. Broederstroom and the central cattle pattern. S Afr J Sci 1995;89(5):220-226. [ Links ]
10. Hall M. The role of cattle in southern African agropastoral societies: More than bones alone can tell. Goodwin Series. 1986:83-87. [ Links ]
11. Denbow J. Material culture and the dialectics of identity in the Kalahari: AD 700-1700. Beyond chiefdoms. In: McIntosh SK. Pathways to complexity in Africa. Cambridge: Cambridge University Press; 1999. p. 110-123. [ Links ]
12. Hall S. Farming communities of the second millennium: Internal frontiers, identity, continuity and change. In: Hamilton C, Mbenga BK, Ross R, editors. The Cambridge history of South Africa. vol. 1. Cambridge: Cambridge University Press; 2010. p. 112-167. [ Links ]
13. Huffman TN. The archaeology of the Nguni past. Southern Afr Hum. 2004;16:79-111. [ Links ]
14. Huffman TN. Ceramics, settlements and late Iron Age migrations. Afr Archaeol Rev. 1989;7(1):155-182. http://dx.doi.org/10.1007/BF01116842 [ Links ]
15. Huffman TN. Regionality in the Iron Age: The case of the Sotho-Tswana. Southern Afr Hum. 2002;14:1-22. [ Links ]
16. Maggs T. Iron Age communities of the southern Highveld. Pietermaritzburg: Natal Museum; 1976. [ Links ]
17. Boeyens JC. The Late Iron Age sequence in the Marico and early Tswana history. S Afr Archaeol Bull. 2003:63-78. [ Links ]
18. Esterhuysen A, Sanders V, Smith J. Human skeletal and mummified remains from the AD1854 siege of Mugombane, Limpopo South Africa. J Archaeol Sci. 2009;36(4):1038-1049. http://dx.doi.org/10.1016/j.jas.2008.12.006 [ Links ]
19. Hamilton C. The Mfecane aftermath. Johannesburg: University of the Witwatersrand Press; 1995. [ Links ]
20. Pereira L, Gusmao L, Alves C, Amorim A, Prata MJ. Bantu and European Y-lineages in sub-Saharan Africa. Ann Hum Genet. 2002;66(5-6):369-378. http://dx.doi.org/10.1046/j.1469-1809.2002.00130.x [ Links ]
21. Ribot I, Morris AG, Sealy J, Maggs T. Population history and economic change in the last 2000 years in KwaZulu-Natal, RSA. Southern Afr Hum. 2010;22:89-112. [ Links ]
22. Steyn M. A reassessment of the human skeletons from K2 and Mapungubwe (South Africa). S Afr Archaeol Bull. 1997:14-20. http://dx.doi.org/10.1080/0043824032000111425 [ Links ]
23. LAbbe EN, Ribot I, Steyn M. A craniometric study of the 20th century Venda. S Afr Archaeol Bull. 2006:19-25. [ Links ]
24. Steyn M. A comparison between pre-and post-colonial health in the northern parts of South Africa, a preliminary study. World Archaeol. 2003;35(2):276-288. [ Links ]
25. Rightmire G. Iron age skulls from Southern Africa re-assessed by multiple discriminant analysis. Am J Phys Anthropol. 1970;33(2):147-167. http://dx.doi.org/10.1002/ajpa.1330330203 [ Links ]
26. Nienaber W, Keough N, Steyn M, Meiring J. Reburial of the Mapungubwe human remains: An overview of process and procedure. S Afr Archaeol Bull. 2008;63(188):164-169. [ Links ]
27. DeVilliers H. The skull of the South African negro. Johannesburg: Witwatersrand University Press; 1968. [ Links ]
28. Franklin D, Cardini A, Oxnard CE. A geometric morphometric approach to the quantification of population variation in sub-Saharan African crania. Am J Hum Biol. 2010;22(1):23-35. http://dx.doi.org/10.1002/ajhb.20908 [ Links ]
29. Jacobson A. The dentition of the South African negro. Anniston, AL: Higginbotham; 1982. [ Links ]
30. Irish JD. Characteristic high- and low-frequency dental traits in sub-Saharan African populations. Am J Phys Anthropol. 1997;102(4):455-467. http://dx.doi.org/10.1002/(SICI)1096-8644(199704)102:4<455::AID-AJPA3>3.0.CO;2-R [ Links ]
31. Irish JD. Ancestral dental traits in recent sub-Saharan Africans and the origins of modern humans. J Hum Evol. 1998;34(1):81-98. http://dx.doi.org/10.1006/jhev.1997.0191 [ Links ]
32. Tishkoff SA, Reed FA, Friedlaender FR, Ehret C, Ranciaro A, Froment A, et al. The genetic structure and history of Africans and African Americans. Science. 2009;324(5930):1035-1044. http://dx.doi.org/10.1126/science.1172257 [ Links ]
33. Franklin D, Freedman L, Milne N, Oxnard C. Geometric morphometric study of population variation in indigenous southern African crania. Am J Hum Biol. 2007;19(1):20-33. http://dx.doi.org/10.1002/ajhb.20569 [ Links ]
34. Coppa A, Cucina A, Mancinelli D, Vargiu R, Calcagno JM. Dental anthropology of Central-Southern, Iron Age Italy: The evidence of metric versus nonmetric traits. Am J Phys Anthropol. 1998;107(4):371-386. http://dx.doi.org/10.1002/(SICI)1096-8644(199812)107:4<371::AID-AJPA1>3.0.CO;2-9 [ Links ]
35. Hanihara T, Ishida H. Metric dental variation of major human populations. Am J Phys Anthropol. 2005;128(2):287-298. http://dx.doi.org/10.1002/ajpa.20792 [ Links ]
36. Hanihara T. Morphological variation of major human populations based on nonmetric dental traits. Am J Phys Anthropol. 2008;136(2):169-182. http://dx.doi.org/10.1002/ajpa.20792 [ Links ]
37. Irish JD. Who were the ancient Egyptians? Dental affinities among Neolithic through postdynastic peoples. Am J Phys Anthropol. 2006;129(4):529-543. http://dx.doi.org/10.1002/ajpa.20261 [ Links ]
38. Irish JD. 12 Afridonty: The "Sub-Saharan African Dental Complex" revisited. In: Scott GR, Irish JD, editors. Anthropological perspectives on tooth morphology: Genetics, evolution, variation. Vol 66. Cambridge: Cambridge University Press; 2013. p. 278. http://dx.doi.org/10.1017/CBO9780511984464.012 [ Links ]
39. Jackes M, Silva AM, Irish J. Dental morphology: A valuable contribution to our understanding of prehistory. J Iber Archaeol. 2001;3:97-119. [ Links ]
40. LAbbé EN, Van Rooyen C, Nawrocki S, Becker PJ. An evaluation of non- metric cranial traits used to estimate ancestry in a South African sample. Forensic Sci Int. 2011;209(1):195. [ Links ]
41. Warren KA. Population variation within the Iron Age of southern Africa: An assessment using dental anthropological and cranio-mandibular metric techniques [dissertation]. Cape Town: University of Cape Town; 2013. [ Links ]
42. Dayal MR, Kegley AD, Strkalj G, Bidmos MA, Kuykendall KL. The history and composition of the Raymond A. Dart Collection of human skeletons at the University of the Witwatersrand, Johannesburg, South Africa. Am J Phys Anthropol. 2009;140(2):324-335. http://dx.doi.org/10.1002/ajpa.21072 [ Links ]
43. Turner CI. Scoring procedures for key morphological traits of the permanent dentition: the Arizona State University dental anthropology system. In: Kelley MA, Larson CS, editors. Advances in dental anthropology. New York: Wiley-Liss; 1991. p. 13-31. [ Links ]
44. Sottysiak A. Technical Note: An R script for Smith's Mean Measure of Divergence. Bioarchaeol Near East. 2011;5:41-44. [ Links ]
45. Mitchell P. Genetics and southern African prehistory: An archaeological view. J Anthropol Sci. 2010;88:73-92. [ Links ]
46. Loubser JHN. The ethnoarchaeology of Venda-speakers in southern Africa. Bloemfontein: Nasionale Museum; 1991. [ Links ]
47. Rasmussen RK. Migrant kingdom: Mzilikazi's Ndebele in South Africa. London: Rex Collings; 1978. [ Links ]
48. Hall S, Smith B. Empowering places: Rock shelters and ritual control in farmer-forager interactions in the Northern Province. Goodwin Series. 2000:30-46. [ Links ]
49. Esterhuysen A. Divining the siege of Mugombane. J Anthropol Archaeol. 2008;27(4):461-474. http://dx.doi.org/10.1016/j.jaa.2008.08.001 [ Links ]
50. Newitt MDD. A history of Mozambique. Bloomington, IN: Indiana University Press; 1995. [ Links ]
51. Scott GR, Turner CG. The anthropology of modern human teeth: Dental morphology and its variation in recent human populations. Cambridge: Cambridge University Press; 1997. [ Links ]
52. Matsumura H, Hudson MJ. Dental perspectives on the population history of Southeast Asia. Am J Phys Anthropol. 2005;127(2):182-209. http://dx.doi.org/10.1002/ajpa.20067 [ Links ]
53. Greenberg JH, Turner CG, Zegura SL, Campbell L, Fox JA, Laughlin W, et al. The settlement of the Americas: A comparison of the linguistic, dental, and genetic evidence [and comments and reply]. Curr Anthropol. 1986:477-497. http://dx.doi.org/10.1086/203472 [ Links ]
54. Christensen AF. Odontometric microevolution in the Valley of Oaxaca, Mexico. J Hum Evol. 1998;34(4):333-360. http://dx.doi.org/10.1006/jhev.1997.0194 [ Links ]
55. Shaw JM. Taurodont teeth in South African races. J Anat. 1928;62(Pt 4):476. [ Links ]
56. Shaw JCM. The teeth, the bony palate and the mandible in Bantu races of South Africa. London: John Bale, Sons & Danielsson; 1931. [ Links ]
57. Kieser JA, Cameron N, Groeneveld H. Evidence for a secular trend in the Negro dentition. Ann Hum Biol. 1987;14(6):517-532. http://dx.doi.org/10.1080/03014468700009361 [ Links ]
 Correspondence:
Correspondence:
Kerryn Warren
Department of Archaeology
University of Cape Town
Private Bag, Rondebosch 7701
South Africa
wrrker001@myuct.ac.za
Received: 02 Sep. 2013
Revised: 24 Jan. 2014
Accepted: 04 Feb. 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}