










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.109 no.11-12 Pretoria Jan. 2013
REVIEW ARTICLES
Bridging disciplines to better elucidate the evolution of early Homo sapiens in southern Africa
Marlize LombardI; Carina SchlebuschII; Himla SoodyallIII
IDepartment of Anthropology and Development Studies, University of Johannesburg, Johannesburg, South Africa
IIDepartment of Evolutionary Biology, Evolutionary Biology Centre, Uppsala University, Uppsala, Sweden
IIIDivision of Human Genetics, School of Pathology, Faculty of Health Sciences, University of the Witwatersrand and National Health Laboratory Service, Johannesburg, South Africa
ABSTRACT
Elucidating the history of Homo sapiens has been a passion shared by many researchers spanning several decades. There are now overwhelming lines of evidence from genetic, archaeological, palaeoanthropological and, to some extent, palaeoenvironmental research, that place Africa as the region of origin of our species. The different fields of study use diverse types of data, and methods are subject to variances introduced by mutation rates, time estimates and/or sampling biases. All of these approaches have their respective shortcomings and error ranges and are accompanied by intense debate. Yet, it is timeous to review the most recent and salient highlights that the different approaches are contributing towards explaining our deep history and ancestry. It is, after all, one history, and consequently, there ought to be several convergent patterns between data sets. Our focus is to present an updated regional synthesis from each discipline for a specific window in time within the southern African context, namely between ~ 160 ka and 85 ka, and to speculate about possible connections between data sets for this period. Even though our focus is specific in time and space, it is not intended to consider southern Africa in isolation from the rest of Africa or to suggest a singular 'origins' locale for modern Homo sapiens. We hope that this integrated approach will stimulate discussions to include broader time periods within Africa and between continents.
Keywords: Homo sapiens; genetic studies; archaeology; palaeoanthropology; southern Africa
Genetic insights into the past
The genetic approach, when reconstructing the past, focuses on the patterns of genetic variation found in living people, and in some instances uses ancient DNA found in human remains such as skeletal material and teeth. Computational methods are then applied to examine how the changes or mutations in DNA can be traced to a common point in the past. Since the 1980s, haploid genomes - the maternally transmitted mitochondrial DNA (mtDNA) and the paternally transmitted Y-chromosome DNA - are widely used in population genetic studies to examine the genetic affinities of groups around the globe. Several independent studies have now shown that the ancestors of all living humans diverged from a common mtDNA ancestor before 100 ka, and that the most divergent mtDNA haplogroup, L0d, is strongly associated with the Khoe-San as well as some Coloured groups from southern Africa.1-4 Haplogroup L0d is found at high frequencies in all Khoe-San and selected Coloured groups, ranging in frequency from 45% in the Cape Coloured group3 to 100% in the Karretjie People1. Using mtDNA control region sequence variation, haplogroup L0d was estimated to have a coalescence time of 87±18 ka and to have diverged from the other L0 groups about 96±20 ka.4 These age estimates are shallower in time when compared with estimates based on whole mtDNA genome sequencing data. In the latter case, the coalescent time for L0d was ~100 ± 10 ka, the divergence time of L0d from other L0 haplogroups was ~152 ± 12.5 ka, and the time to the most recent common ancestor for all divergent mtDNA lineages was estimated to be ~200.8 ± 12.7 ka.5
Until recently, the oldest Y-chromosome haplogroup, haplogroup A, was found at high frequencies in Khoe-San groups in southern Africa, and at lower but appreciable frequencies in non-Niger-Kordofanian-speaking African populations in north, east, west and central Africa. However, the recent discovery of a new Y-chromosome haplogroup - haplogroup A006, with a coalescence time of ~209 ka (338 ka with newer mutation rate estimates7) - was found at low frequencies in western Africa, but not among southern African populations.6 Here we also draw attention to the fact that dating in genetic studies is subject to various biases and errors, and that estimates should not be viewed as finite. For example, new sequencing efforts indicate that the mutation rate in humans today is about half of what has been used up until now in evolutionary genetic studies.7,8 Genetic dates might, therefore, be older than reported thus far. It has also been suggested that the molecular clock may not be steady for primate lineages and might have slowed down over time. Until consensus is reached, the most parsimonious approach is to use formerly established rates, even though some or all might be adjusted in the future.
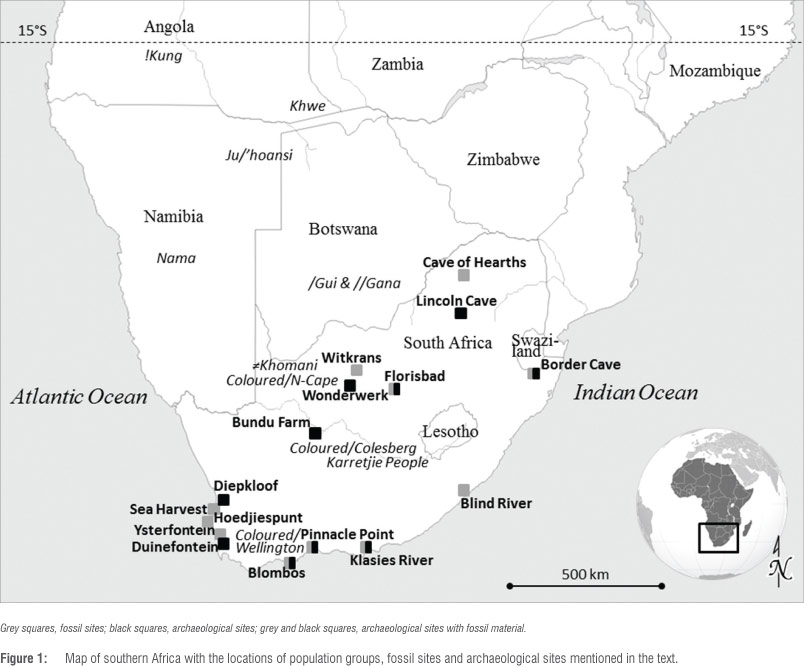
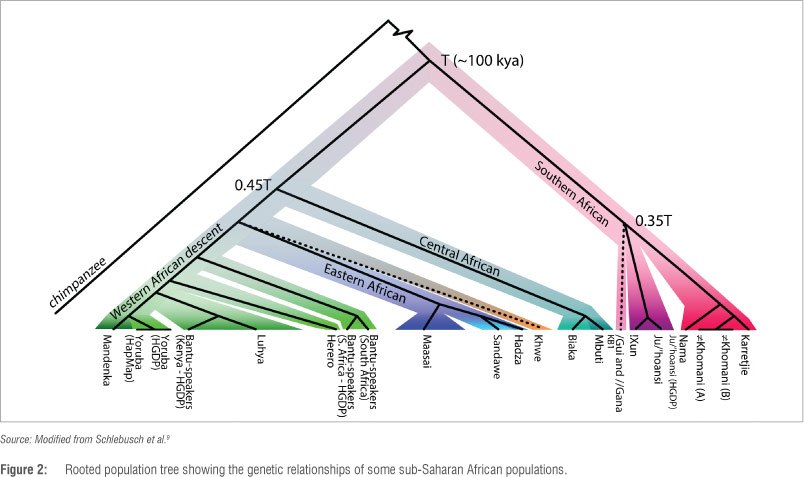
Recent technological advances have facilitated the examination of whole human genomes in a quicker and more cost-effective manner, permitting a robust and simultaneous analysis of many nucleotides. Such genome-wide studies, using single nucleotide polymorphisms (SNPs), examined the similarities and differences among African populations, and highlight the unique genetic make-up of Khoe-San groups.9,10 One of these studies examined ~2.3 million SNPs from a group of 220 individuals that comprised 11 southern African populations: Ju/'hoansi, !Xun, /Gui and //Gana, Karretjie People, ≠Khomani, Nama, Khwe, Coloured from Colesberg, Coloured from Wellington, Herero and south-eastern Bantu-speaking (Figures 1 and 2).9
These data were compared with data from other Africans. Although the merged data were based on less information (~270 000 SNPs instead of ~2.3 million SNPs), the results were consistent with the deepest branch in the human tree harbouring six Khoe-San groups (Khwe excluded) dating to ≥100 ka, almost twice the age of other African populations (Figure 2).9 This finding supports previous estimates for the division at ~160-110 ka.11,12 Moreover, there was evidence of genetic structure within the Khoe-San that showed a separation between northern (Ju speakers: !Xun and Ju/'hoansi) and southern Khoe-San groups (Tuu and Khoe speakers: Karretjie, ≠Khomani and Nama) dating to ~35 ka. The Khwe clustered on a separate, unique branch, which could be because of admixture with non-Khoe-San African groups (Figures 1 and 2).
Another study, which examined human variation in conjunction with the spread of Helicobacter pylori strains (Gram-negative, micro-aerophilic bacteria found in the stomach), estimated that the H. pylori strain that is uniquely associated with Khoe-San populations, diverged from all other human associated H. pylori strains at ~116-88 ka.13 By examining the co-evolution of H. pylori and humans, an independent scrutiny of deep human ancestry is provided.
Context within the MIS chronology
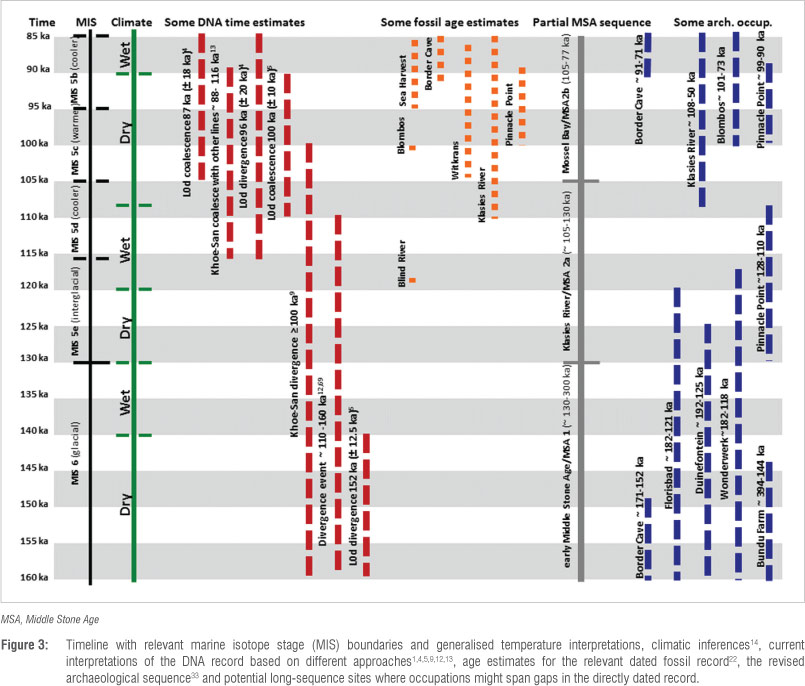
Using the current time estimates of ~160-88 ka from the genetic data as a proxy for the history of Homo sapiens in southern Africa, we explore possible connections with the palaeoenvironments and marine isotope stages (MISs) in which these ancestors would have lived. Most of the earliest population and lineage divergences in our species, based on existing genetic dates, involve human populations currently living in southern Africa. These divergence dates overlap with the younger part of MIS 6 (~195-130 ka) and MIS 5b-e (~130-85 ka) (Figure 3). Even though we use the MIS record as a broad, global time frame and palaeoenvironmental backdrop, we acknowledge that MIS boundaries might not account for many of Africa's climate changes,14 or all variation in human behaviour.15
MIS 6 is generally seen as one of the longest, coldest glacial periods.16 Its effect on the ecosystems of southern Africa remains poorly understood.17 Comparative work, based on a range of data, shows that generalised climatic patterns derived from northern hemisphere glacial/ interglacial cycles are imprecise when applied to the African record.14 Nonetheless, in sub-Saharan Africa, glacial periods such as MIS 6 were often, but not exclusively, accompanied by aridity with expanding deserts and arid regions.18,19 Such regions could have hampered inter-and/or intra-regional movement of humans and other animals.17 Data for MIS 6 in South Africa indicate two dry phases, interspersed by a wet phase at ~140-130 ka (Figure 3).14
MIS 6 was followed by MIS 5e, a sub-stage during which global conditions warmed and sea levels probably rose rapidly.20 During such interglacial phases wetter climates and extended woodlands are expected, yet these remain general trends that might not apply equally to all regions or environmental niches.18,19 As a whole, MIS 5 comprises a sequence of sub-stages alternating between warmer and cooler conditions (Figure 3).21 MIS 5e was an early warm interval, and was followed by irregular, but generally cooler, conditions culminating in the glacial conditions of MIS 4 (~71/74-60 ka). Inferred climatic conditions point to a dry spell spanning the MIS 6-5 boundary and again at ~108-90 ka (Figure 3).14
Similar to current patterns, past climates varied with latitude, so that periods of relatively increased aridity or humidity were asynchronous across the northern, eastern, tropical and southern portions of Africa.14 Changes most likely occurred under different regimes of climate forcing, creating alternating opportunities for migration into adjacent areas. For continental Africa, current data suggest: (1) little relation between large-scale demographic and climate change in southern Africa between ~130 ka and 30 ka, (2) strong support of hominin occupation of the Sahara during discrete humid intervals from ~135 ka to 115 ka and from ~105 to 75 ka and (3) that hominin populations in equatorial and eastern Africa may have been buffered from the extremes of climate change.14 The data are also consistent with human migrations out of Africa through varying exit points during ~140-80 ka.14
The palaeoanthropological record and genetic data
Fossil remains, dated close to the estimate of genetic divergence times of current southern Africans from other humans, could possibly be linked to the common ancestral H. sapiens population. For example, by ~160-80 ka some of the earliest modern H. sapiens fossil material is reported from South African contexts (Figures 1 and 3) 22:
• A phalanx and tooth from Sea Harvest mostly likely date to MIS 5b (~95-85 ka).
• Human teeth from Witkrans probably date to ~103-86 ka.
• Human teeth at Blombos occur in deposits with age estimates of ~100 ka.
• Human cranial fragments, jaws, teeth and post-cranial bones from Klasies River were found in archaeological contexts with age estimates of ~110 ka, ~100 ka and ~100-80 ka.
• A partial femur from Blind River dates to MIS 5e (~118 ka).
• A partial cranium and mandible from Border Cave with age estimates of either >150 ka or ~91-71 ka, could be relevant to our time slice. Because the material was found in secondary context, however, the age estimates are still insecure. A partial infant skeleton found in a grave at the site is thought to date to ~90-66 ka, but its context too has been questioned.
With the exception of Klasies River that produced more than 30 specimens, the fossil sample that can be traced to this period with a measure of surety is small, and the majority of the specimens are isolated teeth. Based on morphological characteristics, the population is mainly accepted as H. sapiens, but some specimens exhibit archaic morphology.23 It seems that during this period South Africa was inhabited by populations who showed a wide range of anatomical variation, mostly morphologically similar to present-day H. sapiens. However, the process of gracilisation (reduction of prominence of facial features) was not yet completed,22 and did not yet reflect more recent Khoe-San morphology.24
Gracilisation is expected to be accompanied by several morphological changes. We suggest that certain aspects of gracilisation could be the result of gene interactions under the influence of selective forces. Some evolutionary processes that might have influenced the shaping of genomic variation in humans at >100 ka have been identified in genetic studies.9,25,26 Of the five genomic regions that were found to have signatures for strong selective pressure in humans before ~100 ka,9 three contained genes that were associated with skeletal development, and may have contributed to the process of gracilisation. Positive signals for selection were obtained in a region immediately upstream of the ROR2 gene that is involved in the regulation of bone and cartilage development and SPTLC1, a gene involved in hereditary sensory neuropathy. Other signals for selection were obtained for the region containing the SULF2 gene that regulates cartilage development, and a region containing RUNX2, which is involved in skeletal morphogenesis.9
Tentatively, variations in RUNX2 have also been linked to phenotypic differences between H. sapiens and archaic humans, such as frontal bossing, clavicle morphology, a bell-shaped ribcage and regulating the closure of the fontanel26-28 - the latter being critical for brain expansion. Perhaps also relevant, is that a fixed difference between the DNA of archaic and modern humans occurs in EVC2, which when mutated in modern humans, causes Ellis-van Creveld syndrome.26 Among other symptoms, this syndrome includes taurodontism, an enlargement of the dental pulp cavity and fusion of the roots. Taurodontism is a common trait in the teeth of H. neanderthalensis and other archaic humans. Although a Denisovan molar also has an enlarged pulp cavity, it lacks fused roots.26 These observations suggest that mutations in EVC2, perhaps in conjunction with mutations in other genes, might have caused a change in the dental morphology of early H. sapiens. Because gracile H. sapiens morphology appeared fairly abruptly, selection on a few morphology genes such as those mentioned above, could have been involved in the 'anatomical' emergence of our species.9,26 Morphological changes are, however, complex, and further work needs to test these suggestions.
Homo fossils in the South African record pre-dating 200 ka, and considered to perhaps represent morphological trends leading up to current anatomical characteristics, include for example (Figure 1)22:
• A partial cranium from Florisbad dated to 259±35 ka that is ascribed to Homo helmei or late archaic H. sapiens.
• A mandible and radius from Cave of Hearths ascribed to archaic H. sapiens and/or early H. sapiens that have an age estimate of ~500-200 ka.
• A tibia and teeth (most of which show modern morphology, but are larger than the contemporary African average) from Hoedjiespunt, dated to ~300-200 ka, that are ascribed to H. heidelbergensis.
The relevant South African Homo fossil record before ~200 ka is sparse, and presently we are not aware of material that can be placed securely at ~200-120 ka, except perhaps the material from Border Cave that has a questionable age estimate of ~171-152 ka. This gap could simply reflect current sampling, preservation and/or dating deficits. On the other hand, possible morphologically intermediary fossil material is present in assemblages pre- and post-dating ~200-120 ka. The gap in the fossil record could be seen as evidence that early H. sapiens evolved elsewhere in Africa and migrated to southern Africa, but this possibility should be considered in the light of the archaeological record, some aspects of which are highlighted below. Fossil material from other African regions dating to this period is also sparse, but include remains from Omo Kibish, Ethiopia, at ~195 ka,29 Herto, Ethiopia, at 160-154 ka,30,31 and Singa, Sudan, at >133 ka.32 These remains have all been interpreted as either archaic (potentially ancestral to modern humans) or anatomically modern H. sapiens, and they provide direct evidence of human presence in east Africa at the time.
The archaeological record, human behaviour and the genetic record
In contrast to the fossil record, the South African archaeological record supports human occupation at several sites during ~200120 ka (Figure 1). Even though the dating of some contexts remains controversial, examples of sites include (Figures 1 and 3)22,33:
• Pinnacle Point 13B - MIS 5 assemblages have age estimates of ~128-110 ka, and MIS 6 assemblages of ~166-160 ka.
• Bundu Farm - early Middle Stone Age assemblages have best age estimates of ~394-144 ka.
• Duinefontein - has occupations dated to ~292-125 ka.
• Florisbad - ages for the early Middle Stone Age range from ~281 ka to 121 ka.
• Wonderwerk Cave - best age estimates for early Middle Stone Age assemblages are ~182-118 ka..
• Ysterfontein - ages of ~132-128 ka were obtained, but their implications are still being discussed.
• Border Cave - even though the context of the fossil material being associated with our chosen time slice can be questioned, the site has archaeology with age estimates of ~171-152 ka, ~91-71 ka and ~90-60 ka.
This group of assemblages represents a parsimonious reflection of human presence at the time. Many more sites with similar Middle Stone Age archaeology are known from across the region, but dating is weak or absent.
Some genetic time estimates of the first divergences within H. sapiens, derived from present-day populations in southern Africa, overlap with the archaeological record of a section of the Middle Stone Age in sub-Saharan Africa (Figure 3). Wurz34 provides a synthesis of developments in stone tool technologies in southern Africa for the period under discussion. According to her, prepared core technology with blades and points, typical of the Middle Stone Age, starts before MIS 7 (~244-195 ka) and continues into MIS 6 with inter- and intra-site variability, but without discernible technological breaks. By MIS 5, Middle Stone Age assemblages become widespread and a variety of technological strategies, perhaps representing technological trends that developed from earlier technological approaches, are evident. Early Middle Stone Age assemblages are typically >130 ka and characterised by prepared core technologies in what can be considered components of generalised toolkits.34,35 Thus, apart from indicating human presence on the landscape, technological approaches also do not seem to point to discernible breaks, yet this interpretation could be an artefact of still coarse research resolution.35
Prepared core technologies, also referred to as Mode 3, provide a level of flexibility in predetermining tool shapes that is not evident in preceding knapping strategies.34 Such flexibility is expressed in the variability observed in and between stone tool assemblages.36 But it is not only knapping strategies that provide insight into human technological and associated cognitive capacity at the time. It has been suggested that hafted technologies, such as stone-tipped spears, often associated with Mode 3 artefacts, signal development towards advanced technological, behavioural and cognitive flexibility, opening the way towards a considerable expansion of, and variability in, problem solution ability.37,38 Ambrose39 argues that composite tools, such as hafted hunting weapons, place greater demands on integrating working memory with prospective memory, and ultimately constructive memory.40 These faculties are associated with the anterior frontal lobe of the human brain and Broca's area that also facilitate processes involved with grammatical language and manual hierarchical assembly.39,41 Early H. sapiens were, however, not the only hominins to produce composite tools, and thus probably shared the associated cognitive and behavioural traits with H. neanderthalensis, H. heidelbergensis and other archaic humans.38
Other interesting trends that appear in the archaeological record, concurrently with genetic evidence for the divergence of the Khoe-San ancestral population and fossil evidence of early H. sapiens, include so-called symbolically mediated behaviours. For example, the use of red ochre has, arguably, been interpreted as circumstantial evidence for the painting of abstract designs on the bodies of ritual performers.42 Utilised red ochre nodules occur in South Africa associated with archaeological material at Pinnacle Point 13B by ~164 ka.43 From ~100 ka, such red pigments become a common feature at most Middle Stone Age sites. The recent find of an ochre-processing workshop at Blombos Cave,44 in which a liquefied ochre-rich mixture was produced and stored in two abalone (Haliotis midae) shells by ~100 ka, confirms that pigment use was firmly established at the time. Perhaps more secure evidence of symbolically mediated behaviour includes engraved ochre nodules, some with complex geometric patterns, also from contexts dating to ~100 ka at Blombos, Pinnacle Point 13B and Klasies River.45,46 But, even when evidence of symbolic thinking (i.e. the capacity to attribute specific meaning to conventional signs) is apparent, it is a trait shared with a growing number of hominins.47 Yet, it can hardly be argued that during the period under discussion we see early signals of a uniquely human way of doing and thinking in the archaeological record of southern Africa. Whereas our focus is the southern African records, we draw attention to the fact that the discussion of the above archaeology also roughly reflects that of sub-Saharan Africa as a whole,48 so that any behavioural and cognitive inferences apply to the greater sub-continental region as well as regions in north Africa.49-53
Some of these behavioural and cognitive trends can, conceivably, also be explored through genetic research. By using what is known from medical science and comparing the DNA in contemporary humans with that of archaic humans such as Neanderthals and the Denisovan specimen, researchers are finding tentative links with certain genetic variants involved in cognitive function that might be exclusive to H. sapiens.25,26 Furthermore, when compared with Neanderthals, several genes involved in cognitive development showed strong evidence for selection in early H. sapiens.25 These genes include DYRK1A that is medically associated with cognitive impairment, NRG3 medically associated with schizophrenia, and two genes, CADPS2 and AUTS2, medically associated with autism.25 Other genes linked with development and neuronal function have also been shown to be strongly selected for in early humans.9SDCCAG8 has been medically associated with microcephaly, LRAT with Alzheimer's disease and SULF2 with distorted brain development.54 And ROR2, that was previously mentioned in relation to its role in skeletal development, is upregulated by FOXP2,55 which is the only gene currently known to be involved with speech and language disorders that follows Mendelian inheritance.26,56,57
The above-mentioned medical associations with genes are mere pointers to some of the gene/genomic region's functions. Most of these functional associations and their evolutionary advantages are still poorly understood. With further research, particularly when linked with gene expression studies, we should learn more about the developmental processes that distinguish our species from our early ancestors.
What have we learnt from the multidisciplinary approach?
It has been suggested58 that an appropriate theoretical framework for understanding our origins can be achieved by integrating evolutionary geography, based on spatial distributions of populations and contingent, adaptive and demographic aspects of micro-evolutionary change. Following on previous calls for the integration of different data sets,58,59 our cross-disciplinary approach attempts to update and contribute to the broader theoretical framework from a southern African perspective for the period between ~160 ka and 88 ka. Bottleneck scenarios are often used to explain the relative lack of genetic diversity in H. sapiens compared to other species.58,60 Two broad hypotheses have been formulated for a population bottleneck during MIS 6.58 According to the first, humans in sub-Saharan Africa separated into a number of regional groups, some of whom survived, depending on resources and/or their respective evolutionary responses. The second hypothesis suggests that a single (founder) population survived as a result of the interaction between population size and the carrying capacity of the environment. The first scenario would imply that the palaeoanthropological diversity in Africans between ~200 ka and ~100 ka represents true ancestral diversity. On the other hand, if the second hypothesis is accurate, archaeological and Homo fossil diversity in Africa at the time does not represent ancestral diversity, but a single surviving H. sapiens ancestral group.58
Using genetic data, various possible scenarios have been explored regarding the origins of early H. sapiens or anatomically modern humans. One possible scenario is a multiregional origin of H. sapiens, where anatomically modern humans originated in several places in Africa within the same time frame as a result of similar selective pressures. Alternatively, anatomically modern H. sapiens could have originated in one place on the continent and then replaced archaic populations (groups that do not show the distinctive modern human morphology). The third scenario is an intermediate scenario in which anatomically modern H. sapiens originated in one place on the continent and then distributed and produced offspring with 'archaic' groups, resulting in the spread of 'modern' genetic material. Whereas mtDNA and Y-chromosome studies initially alluded to a single origin model, recent studies based on autosomal genetic variation in African populations do not conclusively point to one region of origin within the continent.9 Genetic support, therefore, currently leans toward either a multiregional scenario or a situation in which admixture took place between early H. sapiens and human populations with archaic features. Various recent studies detected indirect evidence of archaic admixture in the genomes of current Africans, albeit at low frequencies.61-63
Human fossil remains could potentially be used to assess bottleneck and/or population isolation scenarios, and it is suggested that expecting craniometric similarities between H. sapiens populations from different regions and genetic relationships is probably misplaced.64 What is more, the palaeoanthropological data also indicate that early H. sapiens populations in South Africa did not yet have the gracile features associated with more recent Khoe-San groups or their immediate ancestors. Rather, such features might have developed more recently as a result of isolation and local adaptation to the environment of the region.24,64,65 The seeming absence of well-contextualised human fossil specimens during a key stage in South Africa could indicate that anatomically modern H. sapiens 'originated' elsewhere on the continent, as is often hypothesised for east Africa.66 The archaeological sequence serves to expose a possible sampling and/or preservation bias in the fossil record, but is itself still plagued with shortcomings in dating resolution and sampling. However, similar to the genetic record, the current archaeological data could be seen to support a local, but not necessarily unique, origin and/or an archaic-modern admixture scenario.
The multidisciplinary approach incorporating information from diverse disciplines afforded us the opportunity to review some aspects of early human history in southern Africa. For example, the 'Cape Floral Region -South Coast Model' for the origins of early H. sapiens during the Middle Stone Age, is linked to a genetic bottleneck during which populations crashed and survived only in favourable refugia, possibly resulting in a single ancestral group.17,67 Yet, South African inland sites such as Bundu Farm, Wonderwerk Cave, Florisbad and Border Cave were also inhabited during MIS 6 (Figures 1 and 3). None of these sites are located near the Cape Floral Region, and all are in zones with very different climatic conditions. Based on the topography of South Africa, past climatic conditions, even though different from current ones, would also have been different between the Cape Floral Region and the inland sites mentioned above. From a genetic viewpoint, the current interpretation that all human mtDNA and Y-chromosome lineages coalesced ~200-300 ka, just before MIS 6, might suggest such a population bottleneck,5,6,68 and numerical simulations on autosomal data to reproduce current genetic diversity initially supported these findings.69 Blum and Jakobsson70, however, suggest that although a bottleneck model at MIS 6 might be viable, a model of multiple archaic African populations surviving and giving rise to early H. sapiens is also plausible. Re-sequencing of the African data furthermore shows that a demographic model without an ancestral bottleneck during MIS 6 is the most likely scenario.71 The work of Li and Durbin72 even indicates an increase in human population sizes during this time, or, alternatively, a scenario in which multiple populations became separated and then mixed again. Additionally, new studies on mtDNA and Y-chromosomes, together with newly proposed mutation rates, keep pushing back the coalescence times of the non-recombining lineages of these uniparental markers, suggesting perhaps deeper ancestries of early humans in Africa.5-7
Based on current understanding of the different data sets and their interpretations, we predict that if a bottleneck scenario with a single surviving population is considered, it probably occurred before MIS 6, and that subsequent H. sapiens populations survived in several African landscapes. This outcome is similar to the best interpretation ventured 15 years ago.58 However, it pushes back the time frame and might explain what we see in the collective records that seem to increasingly indicate a number of regionalised early H. sapiens populations. These groups might have been impacted upon by more recent or secondary bottlenecks, such as the Mt Toba volcanic winter at ~75 ka,60 and/or extended periods of separation before new expansions and admixtures recurred. Palaeoenvironmental data, however, remain coarse-grained. The notion of a volcanic winter in east Africa at ~75 ka and an accompanying population bottleneck is being challenged based on sediments recovered from Lake Malawi.73 The integrative theoretical framework suggested by Lahr and Foley58 includes three key aspects: (1) the geographical relationships between parent and daughter populations, (2) the longevity of populations and their subsequent roles in sub-structuring and diversification and (3) the relationships between environmental change and behavioural innovations in relation to range expansion. This framework requires a multidisciplinary approach. However, currently we do not have the fine-grained resolution in any of the research areas and/ or their associated disciplines to produce the conclusive outline for the origins of our species. Yet, by pooling knowledge we can continue to refine our interpretations of past data sets.
Regardless of ongoing 'origins' debates and shortcomings in ages and data sets, several traits, currently considered to reflect the modern characteristics of our species (anatomically and behaviourally), had their southern African roots in MIS 6-5. And there seems to be acceleration in behavioural and cognitive trends towards complexity and higher levels of variability in the archaeological record from at least ~160 ka, subsequent to the time identified in several genetic studies associated with the establishment of a split in the common population lineage of our species. The current genetic record remains inconclusive as to where in Africa early H. sapiens first emerged, and indeed if there was only one 'origin', but it does not negate a local (southern African) scenario.
This short synthesis shows that much progress has been made regarding how we understand and interpret the presence of early H. sapiens in southern Africa, and that we can come closer to a robust history by interweaving data obtained from multiple approaches (Figure 3). Relationships between the data sets remain tentative, but we suggest they clearly provide impetus for future cross-disciplinary exploration. By collating information from the different disciplines, it seems increasingly possible that the perceived gap between early H. sapiens biology and archaeological interpretations of our behavioural and cognitive evolution will continue to shrink.
Acknowledgements
We thank three anonymous reviewers, friends and colleagues who improved this paper by taking the time to comment on earlier drafts. The research of M.L. is supported by an African Origins grant from the National Research Foundation of South Africa; C.S. was supported by the Wenner-Gren Foundation (Sweden); and H.S. wishes to acknowledge the South African Medical Research Council and the National Health Laboratory Service for financial support. Opinions and mistakes are our own and not those of our funding agencies.
Authors' contributions
M.L. initiated the collaborative study and provided archaeological, palaeoanthropological, environmental and some theoretical content. C.S. and H.S. provided all the genetic data and discussion on aspects of genetic research as well as some theoretical content.
References
1. Schlebusch CM, De Jongh M, Soodyall H. Different contributions of ancient mitochondrial and Y-chromosomal lineages in 'Karretjie people' of the Great Karoo in South Africa. J Hum Genet. 2011;56:623-630. http://dx.doi.org/10.1038/jhg.2011.71 [ Links ]
2. Soodyall H, Schlebusch CM. The genetic landscape of sub-Saharan African populations. In: Kumar D, editor. Genomics and health in the developing world. Oxford: Oxford University Press; 2011. p. 369-381. [ Links ]
3. De Wit E, Delport W, Rugamika CE, Meintjes A, Moller M, Van Helden PD, et al. Genome-wide analysis of the structure of the South African Coloured population in the Western Cape. Hum Genet. 2010;128:145-153. http://dx.doi.org/10.1007/s00439-010-0836-1 [ Links ]
4. Schlebusch CM, Lombard M, Soodyall H. MtDNA control region variation in populations from southern Africa. BMC Evol Biol. 2013;13:56. http://dx.doi.org/10.1186/1471-2148-13-56 [ Links ]
5. Behar DM, Villems R, Soodyall H, Blue-Smith J, Pereira L, Metspalu E, et al. The dawn of human matrilineal diversity. Am J Hum Genet. 2008;82:1130-1140. http://dx.doi.org/10.1016/j.ajhg.2008.04.002 [ Links ]
6. Mendez FL, Krahn T, Schrack B, Krahn A-M, Veeramah KR, Woerner AE, et al. An African American paternal lineage adds an extremely ancient root to the human Y chromosome phylogenetic tree. Am J Hum Genet. 2013;92:1-6. [ Links ]
7. Kong A, Frigge ML, Masson G, Besenbacher S, Sulem P Magnusson G, et al. Rate of de novo mutations and the importance of father's age to disease risk. Nature. 2012;488:471-475. http://dx.doi.org/10.1038/nature11396 [ Links ]
8. Scally A, Durbin R. Revising the human mutation rate: Implications for understanding human evolution. Nat Rev Genet. 2012;13(10):745-753. http://dx.doi.org/10.1038/nrg3295 [ Links ]
9. Schlebusch CM, Skoglund P Sjodin P Gattepaille LM, Hernandez D, Jay F, et al. Genomic variation in seven Khoe-San groups reveals adaptation and complex African history. Science. 2012;338:374-379. http://dx.doi.org/10.1126/science.1227721 [ Links ]
10. Pickrell JK, Patterson N, Barbieri C, Berthold F, Gerlach L, Güldemann T, et al. The genetic prehistory of southern Africa. Nat Commun. 2012;3:1143. http://dx.doi.org/10.1038/ncomms2140 [ Links ]
11. Gronau I, Hubisz MJ, Gulko B, Danko CG, Siepel A. Bayesian inference of ancient human demography from individual genome sequences. Nat Genet. 2011;43:1031-1034. http://dx.doi.org/10.1038/ng.937 [ Links ]
12. Veeramah KR, Wegmann D, Woerner A, Mendez FL, Watkins JC, Destro-Bisol G, et al. An early divergence of KhoeSan ancestors from those of other modern humans is supported by an ABC-based analysis of autosomal resequencing data. Mol Biol Evol. 2012;29:617-630. http://dx.doi.org/10.1093/molbev/msr212 [ Links ]
13. Moodley Y Linz B, Bond RP Nieuwoudt M, Soodyall H, Schlebusch CM, et al. Age of the association between Helicobacter pylori and man. PLoS Pathog. 2012;8:e1002693. http://dx.doi.org/10.1371/journal.ppat.1002693 [ Links ]
14. Whiting Blome M, Cohen AS, Tryon CA, Brooks AS, Russell J. The environmental context for the origins of modern human diversity: A synthesis of regional variability in African climate 150,000-30,000 years ago. J Hum Evol. 2012; 62:563-592. http://dx.doi.org/10.1016/j.jhevol.2012.01.011 [ Links ]
15. Jacobs Z, Roberts RG, Galbraith RF, Deacon HJ, Grun R, Mackay A, et al. Ages for the Middle Stone Age of southern Africa: Implications for human behavior and dispersal. Science. 2008;322:733-735. http://dx.doi.org/10.1126/science.1162219 [ Links ]
16. Petit JR, Jouzel J, Raynaud D, Barkov NI, Barnola JM, Basile I, et al. Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature. 1999;399:429-436. http://dx.doi.org/10.1038/20859 [ Links ]
17. Marean C. Pinnacle Point Cave 13B (Western Cape Province, South Africa) in context: The Cape Floral Kingdom, shellfish, and modern human origins. J Hum Evol. 2010;59:425-443. http://dx.doi.org/10.1016/j.jhevol.2010.07.011 [ Links ]
18. Hetherington R, Wiebe E, Weaver AJ, Carto SL, Eby M, MacLeod R. Climate, African and Beringian subaerial continental shelves, and migration of early peoples. Quat Int. 2008;183:83-101. http://dx.doi.org/10.1016/j.quaint.2007.06.033 [ Links ]
19. Carto SL, Weaver AJ, Hetherington R, Lam Y, Wiebe EC. Out of Africa and into an ice age: On the role of global climate change in the late Pleistocene migration of early modern humans out of Africa. J Hum Evol. 2009;56:139-151. http://dx.doi.org/10.1016/j.jhevol.2008.09.004 [ Links ]
20. Hearty PJ, Hollin JT, Neumann AC, O'Leary MJ, McCulloch M. Global sealevel fluctuations during the Last Interglaciation (MIS 5e). Quat Sci Rev. 2007;26:2090-2112. http://dx.doi.org/10.1016/j.quascirev.2007.06.019 [ Links ]
21. Schackleton NJ, Sánches-Goni MF, Pailler D, Lancelot Y Marine isotope substage 5e and the Eamian interglacial. Glob Planet Change. 2007;36:151-155. http://dx.doi.org/10.1016/S0921-8181(02)00181-9 [ Links ]
22. Dusseldorp GL, Lombard M, Wurz S. Pleistocene Homo and the updated Stone Age sequence of South Africa. S Afr J Sci. 2013;109(5/6):46-52. http://dx.doi.org/10.1590/sajs.2013/20120042 [ Links ]
23. Churchill SE, Pearson OM, Grine FE, Trinkaus E, Holliday TW. Morphological affinities of the proximal ulna from Klasies River main site: Archaic or modern? J Hum Evol. 1996;31:213-237. http://dx.doi.org/10.1006/jhev.1996.0058 [ Links ]
24. Morris AG. Isolation and the origin of the Khoisan: Late Pleistocene and early Holocene human evolution at the southern end of Africa. J Hum Evol. 2002;17:231-240. http://dx.doi.org/10.1007/BF02436374 [ Links ]
25. Green RE, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, et al. A draft sequence of the Neandertal genome. Science. 2010;328:710-722. http://dx.doi.org/10.1126/science.1188021 [ Links ]
26. Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, et al. A high-coverage genome sequence from an archaic Denisovan individual. Science. 2012;338:222-226. http://dx.doi.org/10.1126/science.1224344 [ Links ]
27. Mundlos S, Otto F, Mundlos C, Mulliken J, Aylsworth A, Albright S, et al. Mutations involving the transcription factor CBFA1 cause cleidocranial dysplasia. Cell. 1997;89:773-779. http://dx.doi.org/10.1016/S0092-8674(00)80260-3 [ Links ]
28. Falk D, Zollikofer CPE, Morimoto N, de Leon MSP Metopic suture of Taung (Australopithecus africanus) and its implications for hominin brain evolution. Proc Natl Acad Sci USA. 2012;109:8467-8470. http://dx.doi.org/10.1073/pnas.1119752109 [ Links ]
29. Pearson OM, Royer DF, Grine FE, Fleagle JG. A description of the Omo 1 postcranial skeleton, including newly discovered fossils. J Hum Evol. 2008;55:421-437. http://dx.doi.org/10.1016/j.jhevol.2008.05.018 [ Links ]
30. Clark JD, Beyenne J, WoldeGabriel G, Hart WK, Renne PR, Gilbert H, et al. Stratigraphic, chronological and behavioural contexts of Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature. 2003;423:747-752. http://dx.doi.org/10.1038/nature01670 [ Links ]
31. White TD, Asfaw B, DeGusta D, Gilbert H, Richards GD, Suwa G, et al. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature. 2003;453:742-747. http://dx.doi.org/10.1038/nature01669 [ Links ]
32. Spoor F, Stringer C, Zonneveld F. Rare temporal bone pathology of the Singa calvaria from Sudan. Am J Phys Anthropol. 1998;107:41-50. http://dx.doi.org/10.1002/(SICI)1096-8644(199809)107:1<41::AID-AJPA4>3.0.CO;2-G [ Links ]
33. Lombard M, Wadley L, Deacon J, Wurz S, Parsons I, Mohapi M, et al. South African and Lesotho Stone Age sequence updated (I). S Afr Archaeol Bull. 2012;67:123-144. [ Links ]
34. Wurz S. Technological change in the MSA of South Africa, from MIS 7 to MIS 3. Curr Anthropol. 2013;54(suppl 8):S1-S15. [ Links ]
35. Wurz S. Variability in the Middle Stone Age lithic sequence, 115,000-60,000 years ago at Klasies River, South Africa. J Archaeol Sci. 2002;29:1001-1015. http://dx.doi.org/10.1006/jasc.2001.0799 [ Links ]
36. Shea JJ. Homo sapiens is as Homo sapiens was: Behavioural variability versus 'behavioural modernity' in Paleolithic archaeology. Curr Anthropol. 2011;52:1-35. http://dx.doi.org/10.1086/658067 [ Links ]
37. Haidle MN. Working-memory capacity and the evolution of modern cognitive potential: Implications from animal and early human tool use. Curr Anthropol. 2010;51:S149-S166. [ Links ]
38. Lombard M, Haidle MN. Thinking a bow-and-arrow set: Cognitive implications of Middle Stone Age bow and stone-tipped arrow technology. Camb Archaeol J. 2012;22:237-264. http://dx.doi.org/10.1017/S095977431200025X [ Links ]
39. Ambrose S. Coevolution of composite-tool technology, constructive memory, and language. Curr Anthropol. 2010;51:S135-S147. [ Links ]
40. Coolidge FC, Wynn T. Working memory, its executive functions, and the emergence of modern thinking. Camb Archaeol J. 2005;15:5-26. http://dx.doi.org/10.1017/S0959774305000016 [ Links ]
41. Barham L. A technological fix for 'Dunbar's dilemma'? Proc Br Acad. 2010;158:367-389. [ Links ]
42. Watts I. Red ochre, body-painting, and language: Interpreting the Blombos ochre. In: Botha R, Knight C, editors. The cradle of language. vol. 2. Oxford: Oxford University Press; 2009. p. 62-92. [ Links ]
43. Marean CW, Bar-Matthews M, Bernatchez J, Fisher E, Goldberg P Herries AI, et al. Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature. 2007;449(7164):905-908. http://dx.doi.org/10.1038/nature06204 [ Links ]
44. Henshilwood CS, d'Errico F, Van Niekerk KL, Coquinot Y Jacobs Z, Lauritzen SE, et al. A 100,000-year-old ochre-processing workshop at Blombos Cave, South Africa. Science. 2011;334:219-222. http://dx.doi.org/10.1126/science.1211535 [ Links ]
45. Watts I. The pigments from Pinnacle Point Cave 13B, Western Cape, South Africa. J Hum Evol. 2010;59:392-411. http://dx.doi.org/10.1016/j.jhevol.2010.07.006 [ Links ]
46. Henshilwood CS, d'Errico F, Watts I. Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa. J Hum Evol. 2009;57:27-47. [ Links ]
47. d'Errico F, Stringer CB. Evolution, revolution or saltation scenario for the emergence of modern cultures? Phil Trans R Soc B. 2011;366:1060-1069. http://dx.doi.org/10.1098/rstb.2010.0340 [ Links ]
48. Lombard M. Thinking through the Middle Stone Age of sub-Saharan Africa. Quat Int. 2012;270:140-155. http://dx.doi.org/10.1016/j.quaint.2012.02.033 [ Links ]
49. Vanhaeren M, d'Errico F, Stringer S, James SL, Todd JA, Mienis HK. Middle Paleolithic shell beads in Israel and Algeria. Science. 2006;312:1785-1788. http://dx.doi.org/10.1126/science.1128139 [ Links ]
50. Bouzouggar A, Barton N, Vanhaeren M, d'Errico F, Collcutt S, Higham T, et al. 82.000-year-old shell beads from North Africa and implications for the origins of modern human behaviour. Proc Natl Acad Sci USA. 2007;104:9964-9969. http://dx.doi.org/10.1073/pnas.0703877104 [ Links ]
51. Stringer C, Barton N. Putting North Africa on the map of modern human origins. Evol Anthropol. 2008;17:5-7. [ Links ]
52. Richter D, Moser J, Nami M, Eiwanger J, Mikdad A. New chronometric data from Ifri n'Ammar (Morocco) and the chronostratigraphy of the Middle Palaeolithic in the Western Maghreb. J Hum Evol. 2010;59:672-679. http://dx.doi.org/10.1016/j.jhevol.2010.07.024 [ Links ]
53. Jacobs Z, Roberts RG, Nespoulet R, Abdeljalil El Hajraoui M, Debénath A. Single-grain OSL chronologies for Middle Palaeolithic deposits at El Mnasra and El Harhoura 2, Morocco: Implications for Late Pleistocene human-environment interactions along the Atlantic coast of northwest Africa. J Hum Evol. 2012;62:377-394. http://dx.doi.org/10.1016/j.jhevol.2011.12.001 [ Links ]
54. Kalus I, Salmen B, Viebahn C, Von Figura K, Schmitz D, D'Hooge R, et al. Differential involvement of the extracellular 6-O-endosulfatases Sulf1 and Sulf2 in brain development and neuronal and behavioural plasticity. J Cell Mol Med. 2009;13:4505-4521. http://dx.doi.org/10.1111/j.1582-4934.2008.00558.x [ Links ]
55. Konopka G, Bomar JM, Winden K, Coppola G, Jonsson ZO, Gao F, et al. Human-specific transcriptional regulation of CNS development genes by FOXP2. Nature. 2009;462:213. http://dx.doi.org/10.1038/nature08549 [ Links ]
56. Feuk L, Kalervo A, Lipsanen-Nyman M, Skaug J, Nakabayashi K, Finucane B, et al. Absence of a paternally inherited FOXP2 gene in developmental verbal dyspraxia. Am J Hum Genet. 2006;79:965-972. http://dx.doi.org/10.1086/508902 [ Links ]
57. Enard W, Przeworski M, Fisher SE, Lai CSL, Wiebe V, Kitano T, et al. Molecular evolution of FOXP2, a gene involved in speech and language. Nature. 2002;418:869-872. http://dx.doi.org/10.1038/nature01025 [ Links ]
58. Lahr MM, Foley RA. Towards a theory of modern human origins: Geography, demography, and diversity in recent human evolution. Yearb Phys Anthropol. 1998;41:137-176. [ Links ]
59. Morris AG. Prehistory in blood and bone: An essay on the reconstruction of the past from genetics and morphology. Trans R Soc S Afr. 2005;60:111-114. http://dx.doi.org/10.1080/00359190509520487 [ Links ]
60. Ambrose SH. Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans. J Hum Evol. 1998;34:623-651. http://dx.doi.org/10.1006/jhev.1998.0219 [ Links ]
61. Wall JD, Lohmueller KE, Plagnol V. Detecting ancient admixture and estimating demographic parameters in multiple human populations. Mol Biol Evol. 2009;26:1823-1827. http://dx.doi.org/10.1093/molbev/msp096 [ Links ]
62. Hammer MF, Woerner AE, Mendez FL, Watkins JC, Wall JD. Genetic evidence for archaic admixture in Africa. Proc Natl Acad Sci USA. 2011;108:15123-15128. http://dx.doi.org/10.1073/pnas.1109300108 [ Links ]
63. Lachance J, Vernot B, Elbers CC, Ferwerda B, Froment A, Bodo JM, et al. Evolutionary history and adaptation from high-coverage whole-genome sequences of diverse African hunter-gatherers. Cell. 2012;150:457-469. http://dx.doi.org/10.1016/j.cell.2012.07.009 [ Links ]
64. Grine FE, Bailey RM, Harvati K, Nathan RP, Morris AG, Henderson GM, et al. Late Pleistocene human skull from Hofmeyr, South Africa, and modern human origins. Science. 2007;315:226-229. http://dx.doi.org/10.1126/science.1136294 [ Links ]
65. Pfeiffer S. Conditions for evolution of small adult body size in southern Africa. Curr Anthropol. 2012;53(S6):S383-S394. http://dx.doi.org/10.1086/667521 [ Links ]
66. Stringer C, Andrews P. Genetic and fossil evidence for the origin of modern humans. Science. 1988;239:1263-1268. http://dx.doi.org/10.1126/science.3125610 [ Links ]
67. Compton JS. Pleistocene sea-level fluctuations and human evolution on the southern coastal plain of South Africa. Quat Sci Rev. 2011;30: 506-527. http://dx.doi.org/10.1016/j.quascirev.2010.12.012 [ Links ]
68. Gonder MK, Mortensen HM, Reed FA, de Sousa A, Tishkoff SA. Whole-mtDNA genome sequence analysis of ancient African lineages. Mol Biol Evol. 2007;24:757-768. http://dx.doi.org/10.1093/molbev/msl209 [ Links ]
69. Fagundes NJR, Ray N, Beaumont M, Neuenschwander S, Salzano FM, Bonatto SL, et al. Statistical evaluation of alternative models of human evolution. Proc Natl Acad Sci USA. 2007;104:17614-17619. http://dx.doi.org/10.1073/pnas.0708280104 [ Links ]
70. Blum MGB, Jakobsson M. Deep divergences of human gene trees and models of human origins. Mol Biol Evol. 2011;28:889-898. http://dx.doi.org/10.1093/molbev/msq265 [ Links ]
71. Sjödin P Sjöstrand AE, Jakobsson M, Blum MGB. Resequencing data provide no evidence for a human bottleneck in Africa during the penultimate glacial period. Mol Biol Evol. 2012;29:1851-1860. http://dx.doi.org/10.1093/molbev/mss061 [ Links ]
72. Li H, Durbin R. Inference of human population history from individual whole-genome sequences. Nature. 2011;475:493-497. http://dx.doi.org/10.1038/nature10231 [ Links ]
73. Lane CS, Chorn BT, Johnson TC. Ash from the Toba supereruption in Lake Malawi shows no volcanic winter in East Africa at 75 ka. Proc Natl Acad Sci USA. 2013;110:8025-8028. http://dx.doi.org/10.1073/pnas.1301474110 [ Links ]
 Correspondence:
Correspondence:
Marlize Lombard
Department of Anthropology and Development Studies
University of Johannesburg, Auckland Park Campus
Johannesburg 2006, South Africa
Email: mlombard@uj.ac.za
Received: 13 Mar. 2013
Revised: 06 Aug. 2013
Accepted: 14 Aug. 2013


{kind=link}
{kind=link}
{kind=link}