










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.109 n.5-6 Pretoria Jan. 2013
RESEARCH LETTER
Beyond just species: Is Africa the most taxonomically diverse bird continent?
Chris N. LotzI; John A. CaddickII; Monika FornerIII; Michael I. CherryIV, V
IPO Box 215, Newlands 7725, South Africa
II30 White Oak Drive, Beckenham BR3 6QE, UK
IIIPO Box 219, Stompneusbaai 7382, South Africa
IVDepartment of Botany & Zoology, Stellenbosch University, Stellenbosch, South Africa
VDST Centre of Excellence at the Fitzpatrick Institute of African Ornithology, University of Cape Town, Cape Town, South Africa
ABSTRACT
We analysed avian diversity in 8 similar-sized regions of Africa, and in an additional 16 regions spread across the world; half of these 24 regions were tropical and the other half were temperate. For each region, counts of species, genus, family and order were recorded rather than only a species count. We assert that this approach gives more accurate insights into diversity patterns, as we show that in relatively species-rich parts of the world species are on average taxonomically more similar to each other than in species-poor areas. Northwestern South America is the world's most species-rich region for birds, but we show that sub-Saharan Africa has greater diversity at higher taxonomic levels and is thus arguably the richest corner of the world for birds: the Mozambique-Zimbabwe region displays the highest diversity at the order level (with 30 orders), with all other sub-Saharan regions having between 27 and 29 orders each. Northern India is also extremely diverse (surprisingly so for a marginally temperate region) at all taxonomic levels below that of order. We hope that our study might generate further analyses of avian diversity beyond the species level.
Keywords: systematics; avian diversity; species; genus; conservation
Introduction
Using species as a measure of diversity, without counting higher taxonomic levels such as genera and families, can be misleading. Species can be seen as outer branches of a phylogenetic tree, and instead of just counting these, a more accurate measure of diversity can be obtained by taking systematics into account. Over the past 20 years, indices of diversity have been derived which rely on the information content of phylogenetic classification, providing a measure of taxonomic distinctness,1 in particular 'phylogenetic diversity'.2,3 While these indices are useful provided that the requisite phylogenies are available, in their absence the most practical way of obtaining a broader view of diversity may be simply to count not only species, but genera, families and orders.
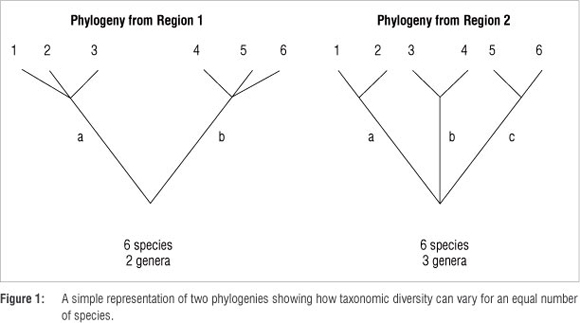
In the case of birds, a wealth of distributional data has been collected by amateurs, some of which has been formalised in atlas projects. These data can be used particularly in optimising choices of which areas to prioritise for conservation,4 but we believe it is important to look beyond species to other taxonomic levels. In bird studies, authors rarely consider other taxonomic levels, despite the fact that the non-avian literature highlights the need to do so.4,5 Similarly when considering endemic bird areas, endemic genera, families and orders should also be (but usually are not) included in assessments. Considering Figure 1, the simplified phylogenies from two hypothetical regions both generate species counts of six. Region 2, however, obviously has a more diverse birdlife, because its six species are distributed across three genera compared to two genera for Region 1. This distribution effectively means that the species of Region 2 are on average genetically more different from each other than those of Region 1, giving a greater 'genetic diversity' in Region 2 despite identical species counts between the two regions.
Although it has been established for some time that the tropics have greater diversity of species than do temperate zones, it was established only recently6 that this pattern, in birds and mammals at least, derives from a faster turnover of species (greater rates of speciation and extinction) at high latitudes. A recent phylogeny for the entire world's extant bird species indicates a strong increase in this rate in the past 50 million years, in particular within songbirds, waterfowl, gulls and woodpeckers.7 Geographically, these recent diversifications have been concentrated in the temperate zones of Asia, North America and southern South America.7
Here we have undertaken a preliminary analysis of patterns of bird diversity in Africa by counting all taxonomic levels from species upwards from eight similar-sized areas of the continent. Using the same method, we also compared avian diversity in 16 of the most species-diverse areas of similar size on five other continents.
Methods
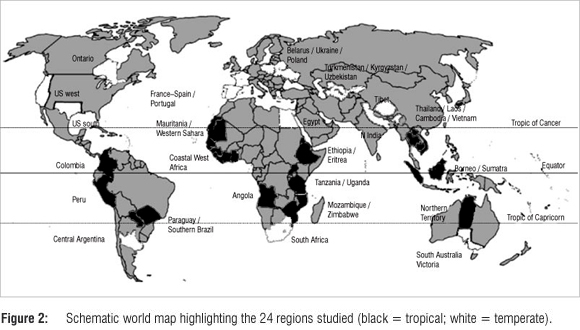
We downloaded bird lists from Avibase8 for regions of similar size, using the taxonomy of the International Ornithological Congress (IOC 2011)9 throughout to control for differences in taxonomy. We counted the number of species, genera, families and orders for each region, removing accidental (vagrant) and introduced (feral or alien) species. By combining lists for smaller countries (e.g. Tanzania and Uganda) we were able to obtain contiguous areas of approximately the same size as medium-sized countries such as South Africa or Colombia, and about the same surface areas as parts of larger countries such as the USA, India or Australia (Figure 2). All of these areas have a coastline and therefore contain the widespread coastal bird families. The smallest region in our analysis is Egypt, which is 74% of the size of the largest region (Northern Territory, Australia). Table 1 shows the area of each region relative to that of Northern Territory (Australia), along with all our other data, sorted by decreasing family diversity.
We used simple regression analyses (Microsoft Excel) to test for relationships between genus count, family count, order count, species:genus ratio, species:family ratio and species:order ratio, all as functions of species count. We did the same for higher taxonomic levels as a function of genus count. Analyses were undertaken separately for tropical and temperate regions.
Results
Colombia and Peru displayed the highest species diversity, with 1816 and 1752 species, respectively (Table 1). Figure 3a shows that there is- a strong relationship between genus and species counts in tropical (r2 = 0.894, F = 84.346, p < 0.001) and temperate (r2 = 0.847, F = 55.238, p < 0.001) regions.
Interestingly, family diversity is not related to species diversity in tropical regions (r2 = 0.002, F = 0.016, p = 0.902) but is in temperate regions (r2 = 0.719, F = 25.643, p < 0.001) (Figure 3b). Family diversity is remarkably low in both Colombia and Peru, with only 86 and 87 families, respectively. Family diversity is exceptionally high in Northern India (104 families) and sub-Saharan Africa (103 families in three regions and 97-101 in all the other sub-Saharan African regions; Table 1). Again, family count plotted as a function of genus count is not significant in tropical regions (r2 = 0.030, F = 0.311, p = 0.589), but is in temperate regions (r2 = 0.562, F = 12.821, p = 0.005) (Figure 3c).
There is no relationship between species and order counts in tropical (r2 = 0.053, F = 0.555, p = 0.473) or temperate (r2 = 0.267, F = 3.647, p = 0.085) regions. Nor is there a relationship between family and order counts in tropical regions (r2 = 0.289, F = 4.061, p = 0.072), but there is in temperate regions (r2 = 0.527, F = 11.162, p = 0.007). It is noteworthy that the top five order counts pertain to the sub-Saharan African regions: 29 each for Coastal West Africa, South Africa, Ethiopia-Eritrea and Angola, and 30 for Mozambique-Zimbabwe, the highest on the planet. No other continent contains regions with more than 28 orders (Table 1).
The more species a region has, the more speciose its families are, in both tropical (r2 = 0.955, F = 214.389, p < 0.001) and temperate (r2 = 0.844, F = 53.999, p < 0.001) regions (Figure 3d). Similarly, the more species a region has, the more speciose its genera are in both tropical (r2 = 0.500, F = 9.988, p = 0.010) and temperate (r2 = 0.491, F = 9.659, p = 0.011) regions. Increasing family and genus sizes as species count increases imply that species become more genetically similar to each other as species diversity increases.
Removing the exclusively pelagic order - Procellariiformes (comprising four pelagic families) - changes none of the trends or statistical significances reported above.
Discussion
To the best of our knowledge, this study is the first to compare higher avian taxonomic diversity across different regions of the world, while controlling for surface area. In terms of higher taxonomy, sub-Saharan Africa is arguably the richest part of the planet for birds, with all six regions sampled having order counts of 28-30, and family counts over 100, with three of the tropical regions (Tanzania-Uganda, Coastal West Africa and Mozambique-Zimbabwe) having 103 families each (Table 1). Northern India has the highest family count of 104, but has an order count of only 26. Bird species in Colombia and Peru are distributed in a surprisingly small number of families (86 and 87, respectively), even though the numbers of species were the highest - 1816 and 1752, respectively. The average bird family sizes in Colombia and Peru are exceptional - 21 and 20 species, respectively - compared with 13 in Uganda-Tanzania (Africa's most species-diverse region with 1324 species) and less than 12 in all other areas we considered (Table 1). Colombia and Peru have some enormous families, such as tanagers and antbirds, containing a large number of genetically similar species.9 We speculate that the large family diversity of marginally temperate Northern India results from the merging of Gondwanaland and Asian taxa when the continents collided, coupled with the current diversity of habitats in this region - from plains at sea level to the world's highest mountain range. Northern India has also been identified as having experienced rapid speciation over the past 50 million years.7
The high and deep-rooted (family and order level) avian diversity in sub-Saharan Africa can be attributed to the Gondwanan origin of a large clade of mainly terrestrial birds which includes the songbirds, Coraciforms, Piciforms, owls, diurnal raptors, trogons, mousebirds, cuckoo-rollers, seriemas, parrots and New World vultures. Three reciprocally monophyletic groups within this clade diversified in Australia, South America and Africa during the Late Cretaceous to early Palaeogene. But although each continent may have originally included other groups, the songbirds and parrots remain the only survivors of the Australian radiation and the falcons and seriemas of the South American radiation. All remaining taxa within the clade are included in the taxonomically diverse group of survivors of the African radiation. In Australia, South America and Africa, different groups have radiated separately in response to local conditions, but the African continent contains by far the largest surviving group of survivors of the Gondwanaland avifauna.10 This history, together with the great diversity of habitats in sub-Saharan Africa, has led to this region having the world's highest levels of avian diversity.
When prioritising which areas to conserve, it also seems imperative that higher taxonomic levels need to be considered, but these are often ignored in work on birds. Once the species of an area are known, it is a trivial exercise to obtain counts of higher taxonomic levels from the species list; two previous studies have indicated the potential importance of this approach. Phylogenetic diversity2 and genus diversity in birds were measured5 using atlas data11 in three provinces of South Africa: Northern Province, Gauteng and Mpumalanga. The minimum number of sites which represent all genera or the maximum phylogenetic diversity of birds was then optimised, and these two measures were equally effective in selecting complementary sets of potential sites for conservation. Changes in breeding birds in New York State over a 20-year period were studied4 using atlas data12,13; changes in both genus and family richness between 1985 and 2005 correlated strongly with changes in species richness and displayed similar spatial patterns, with genus richness being the more reliable predictor of the two. In terms of determining priorities for conservation, considering taxonomic representation above the species level is probably even more crucial in Africa, where resources for this purpose are usually more limited than in the developed world. For example, consider two Important Bird Areas (IBAs) each with eight Endemic Bird Area (EBA) species. The eight species in Malawi's Nyika National Park IBA (within the Tanzania-Malawi Mountains EBA)- are distributed in seven genera, all of which are very widespread African genera. The eight species in Madagascar's Mikea Forest (threatened yet currently unprotected), should be an even higher conservation priority, as they are in eight genera, one of which is found in no other IBA, and three of which are confined to the South Malagasy spiny forests EBA.14 Patterns of endemism, and endemic bird areas in particular, urgently need analysis beyond the species level, as by considering higher taxonomic levels the patterns of diversity (and endemism) revealed can change substantially. We recommend counting at all taxonomic levels in existing IBAs and EBAs as a first step in assessing how priorities might be affected.
Acknowledgements
M.I.C. thanks the National Research Foundation for financial support, and the late Phil Hockey for hosting his sabbatical at the Fitzpatrick Institute, where this paper was written. Comments from three anonymous reviewers significantly improved the manuscript.
Authors' contributions
C.N.L. conceived the idea and wrote the first draft; J.A.C. and M.F. sourced and analysed data; and M.I.C. revised and wrote subsequent versions of the manuscript.
References
1. Vane-Wright RI, Humphries CJ, Williams PH. What to protect? Systematics and the agony of choice. Biol Cons. 1991;55:235-254. http://dx.doi.org/10.1016/0006-3207(91)90030-D
2. Faith DP. Conservation evaluation and phylogenetic diversity. Biol Cons. 1992;61:1-10. http://dx.doi.org/10.1016/0006-3207(92)91201-3
3. Barker GM. Phylogenetic diversity: A quantitative framework for measurement of priority and achievement in biodiversity conservation. Biol J Linn Soc. 2002;76:165-194. http://dx.doi.org/10.1111/j.1095-8312.2002.tb02081.x
4. Kallimanis AS, Mazaris AD, Tsakanikas D, Dimopoulosa P Pantis, JD, Sgardelis SP Efficient biodiversity monitoring: Which taxonomic level to study? Ecol Ind. 2012;15:100-104. http://dx.doi.org/10.1016/j.ecolind.2011.09.024
5. Rodrigues, ASL, Gaston KJ. Maximising phylogenetic diversity in the selection of network of conservation areas. Biol Cons. 2002;105:103-111. http://dx.doi.org/10.1016/S0006-3207(01)00208-7
6. Jetz W, Thomas GH, Joy JB, Hartmann K, Mooers AO. The global diversity of birds in space and time. Nature. 2012;491:444-448. http://dx.doi.org/10.1038/nature11631
7. Weir JT, Schluter D. The latitudinal gradient in recent speciation and extinction rates of birds and mammals. Science. 2007;315:1574-1576. http://dx.doi.org/10.1126/science.1135590
8. Lepage D. Avibase [database on the Internet]. c1992 [cited 2013 Jan 15]. Available from: http://avibase.bsc-eoc.org/avibase.jsp?pg=home&lang=EN [ Links ]
9. Gill F, Donsker D, editors. IOC world bird names v 2.10 [homepage on the Internet]. c2012 [cited 2013 Jan 15]. Available from: http://www.worldbirdnames.org/ [ Links ]
10. Ericson PGP. Evolution of terrestrial birds in three continents: Biogeography and parallel radiations. J Biogeogr. 2012;39:813-824. http://dx.doi.org/10.1111/j.1365-2699.2011.02650.x
11. Harrison JA, Allan DG, Underhill LG, Herremans M, Tree AJ, Parker V, et al. The atlas of southern African birds. Johannesburg: BirdLife South Africa; 1997. [ Links ]
12. Andrle RF, Carroll JR. The atlas of breeding birds in New York State. Ithaca, NY: Cornell University Press; 1988. [ Links ]
13. McGowan K, Corwin K. The second atlas of breeding birds in New York State. Ithaca, NY: Cornell University Press; 2008. [ Links ]
14. Fishpool LDC, Evans MI, editors. Important Bird Areas in Africa and associated islands: Priority sites for conservation. Newbury: Pisces Publications & Cambridge, UK: BirdLife International; 2001. [ Links ]
 Correspondence:
Correspondence:
Michael Cherry
Department of Botany & Zoology, Stellenbosch University
Private Bag X1, Matieland 7602, South Africa
Email: mic@sun.ac.za
Received: 05 Sep. 2012
Revised: 15 Jan. 2013
Accepted: 18 Jan. 2013


{kind=link}