










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.109 no.5-6 Pretoria ene. 2013
RESEARCH ARTICLE
Pathogenicity associated genes in Fusarium oxysporum f. sp. cubense race 4
René SutherlandI; Altus ViljoenII; Alexander A. MyburgIII; Noëlani van den BergIII
IDepartment of Microbiology and Plant Pathology, Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, South Africa
IIDepartment of Plant Pathology, Stellenbosch University, Stellenbosch, South Africa
IIIDepartment of Genetics, Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, South Africa
ABSTRACT
Fusarium oxysporum f. sp. cúbense (Foc) is a fungus that infects banana roots and causes a destructive plant disease called Fusarium wilt. Foc consists of three pathogenic races (Foc races 1, 2 and 4), classified according to their selective impairment of banana cultivars. Foc race 4 is economically important as it comprises strains that infect Cavendish bananas, which are the most widely planted variety of bananas in the world, in both the tropics (Foc TR4) and subtropics (Foc STR4). The aim of this study was to investigate which genes are potentially involved in fungal pathogenicity by comparing transcript-derived cDNA fragments (TDFs) from Foc STR4 and TR4 to those from non-pathogenic F. oxysporum using cDNA-AFLP analysis. This comparison resulted in the identification of 229 unique gene fragments which include the putative pathogenicity-related TDFs encoding chitinase class V (chsV), GTPase activating protein, Major Facilitator Superfamily (MFS) multidrug transporter and serine/threonine protein kinase (ste12) genes. Quantitative analysis of transcript abundance showed a significant increase in expression of chsV, MFS multidrug transporter and ste12 genes in Foc STR4 and TR4 compared with the non-pathogenic F. oxysporum. These genes play a role in escaping host defence responses and in cell wall degradation. In addition, pathogenicity-related genes from other formae speciales of F. oxysporum, such as the sucrose non-fermenting, cytochrome P450 and F-box protein required for pathogenicity genes, were significantly up-regulated in Foc STR4 and TR4 but not in F. oxysporum isolates non-pathogenic to banana. This study provides the first in vitro comparative analysis of TDFs expressed in pathogenic Foc race 4 isolates and non-pathogenic F. oxysporum isolates from banana.
Keywords: banana; cDNA-AFLP; Fusarium oxysporum; pathogenicity; virulence
Introduction
The vascular wilt fungus Fusarium oxysporum is a soil-borne facultative parasite that causes disease in more than 100 plant species, including important agricultural crops.1 The fungus is a morphospecies that is divided into specialised groups (i.e. formae speciales) according to the hosts they attack, and subdivided into races according to the susceptibility of specific host cultivars.2 Host specificity is believed to have evolved independently in F. oxysporum, and does not necessarily reflect phylogenetic relatedness among pathogenic members of the individual hosts.2 In F. oxysporum, host specificity has been attributed to mutations in avirulence genes and lateral chromosome transfer that overcome defence responses in the host plant.3,4
Fungal pathogenicity genes are responsible for events such as spore attachment and germination, infection and colonisation of the host, and are divided into categories such as formation of infection structures, cell wall degradation, toxin biosynthesis and signalling.5-7 Certain pathogenicity genes also encode proteins that are involved in the suppression or disruption of host defence mechanisms.8,9 In F. oxysporum, genes that encode cell wall degrading enzymes (CWDEs), such as endo-polygalacturonase (pg1), exo-polygalacturonase (pgx4), pectate lyase (pl1), xylanase and a plant defence detoxifying enzyme like tomatinase, have been identified in F. oxysporum f. sp. lycopersici (Fol).10-14 Pathogenicity is also influenced by the expression of CWDEs which are regulated by sucrose non-fermenting (snf) gene in F. oxysporum strain O-685 and the F-box protein required for pathogenicity (frp1) gene in Fol.15-17 Signalling genes expressed during pathogenesis have also been identified in Fol (e.g. Fusarium mitogen-activated protein kinase (fmk1))15 and F. oxysporum f. sp. cucumerinum (e.g. G protein α subunit (fga1J and G protein β subunit (fgb1)).18,19 Several transcription factors that regulate pathogenicity genes during infection have been discovered in F. oxysporum, such as serine/threonine protein kinases (ste12),20 a Zn(II)2Cys6-type transcription regulator (ftw2)21 and F. oxysporum ste12 homolog (fost12).22
Strains of F. oxysporum pathogenic to bananas are known as F. oxysporum f. sp. cubense (Foc). Three races of Foc are recognised based on their ability to cause disease in a set of different banana cultivars, with Foc race 1 affecting Gros Michel, Silk and Pome bananas and Foc race 2 affecting Bluggoe and other cooking bananas.23Foc race 4 affects Cavendish bananas, which make up 80% of the world's banana export, as well as Foc race 1 and 2 susceptible bananas.24Foc race 4 is further subdivided into 'tropical' and 'subtropical' strains. Those belonging to Foc 'tropical' race 4 (TR4) are limited to tropical Asia and northern Australia, while Foc 'subtropical' race 4 (STR4) strains are mostly associated with Cavendish bananas in subtropical countries like South Africa, Australia, Taiwan and the Canary Islands. Foc TR4 is more virulent than Foc STR4, and can infect Cavendish bananas under stressed and non-stressed conditions, whereas Foc STR4 typically only infects bananas after the host has been exposed to stressful environments.23
Despite the economic importance of Foc,24 the mechanisms of pathogenesis to banana are still poorly understood. Additionally, non-pathogenic strains of F. oxysporum are known to infect and colonise the cambium tissue of banana roots, but do not enter the xylem to cause Fusarium wilt. Occasionally, the non-pathogens even protect the banana plant from damage caused by Foc25,26 and nematodes.27 It is not known why non-pathogenic strains of F. oxysporum are unable to cause disease to banana. Therefore, the objective of this study was to identify gene transcripts that are present in Foc TR4 and Foc STR 4 but absent in non-pathogenic F. oxysporum using cDNA-amplified fragment length polymorphism (cDNA-AFLP) analysis. In addition, a reverse transcriptase-quantitative polymerase chain- reaction (RT-qPCR) was employed to study the transcript abundance of eight previously described pathogenicity genes from other formas speciales of F. oxysporum.15-17,21,28-31
Materials and methods
Fungal isolates and culture conditions
A total of 27 F. oxysporum isolates were selected for this study. These isolates included Foc STR4 from South Africa, Australia and the Canary Islands, Foc TR4 from Malaysia, Indonesia and Northern Australia, and non-pathogenic F. oxysporum obtained from Cavendish banana roots in South Africa (Table 1). The non-pathogenic F. oxysporum isolates were shown to be non-pathogenic as no internal disease symptoms developed after inoculating banana roots with a spore suspension (1x105 spores/mL) in a hydroponic system.25,27,32,33 All isolates were maintained in 15% glycerol at -80 °C at the Department of Plant Pathology, Stellenbosch University.
RNA extraction
RNA was extracted from fungal mycelia grown in vitro rather than in planta, as insufficient genes of fungal origin were previously detected in the roots of tissue-cultured banana plants 14 d after inoculation with Foc race 4 (1x105 spore/mL). The F. oxysporum isolates were first grown on half strength potato dextrose agar (PDA) (19.5 g/L PDA and 10 g/L agar) for 5 days at ±25 °C, and then transferred to liquid minimal medium (MM) without a carbon source to enhance the transcript abundance of pathogenicity genes.34OhZd After culturing the isolates in MM on a rotary shaker set at 90 rpm for 5 days at 25 °C, the medium was filtered through sterile cheesecloth. The mycelial mass was scraped and frozen in liquid nitrogen, ground to a fine powder with a basic analytical mill (IKA A111, United Scientific (Pty) Ltd., San Diego, CA, USA), and stored at -80 °C until RNA was extracted.
RNA of each isolate was extracted from mycelia using Qiazol (Qiagen, Valencia, CA, USA), quantified with a NanoDrop ND-1000- spectrophotometer (Nanodrop Technologies Inc., Montchanin, DE, USA) and assessed by formaldehyde agarose gel electrophoresis (1.2%). RNA from three isolates collected from the same country or location were pooled (Table 1), DNaseI-treated (Fermentas, Life Sciences, Hanover, MD, USA) and column purified with an RNeasy mini kit (Qiagen). Messenger RNA (mRNA) was isolated using the Oligotex mRNA mini kit (Qiagen). Double-stranded cDNA was synthesised with the cDNA Synthesis System (Roche Diagnostics, Mannheim, Germany) using oligo dT15 primers and contamination was assessed by performing a PCR with the intron flanking primers EF1 and EF2.35
cDNA-AFLP analysis
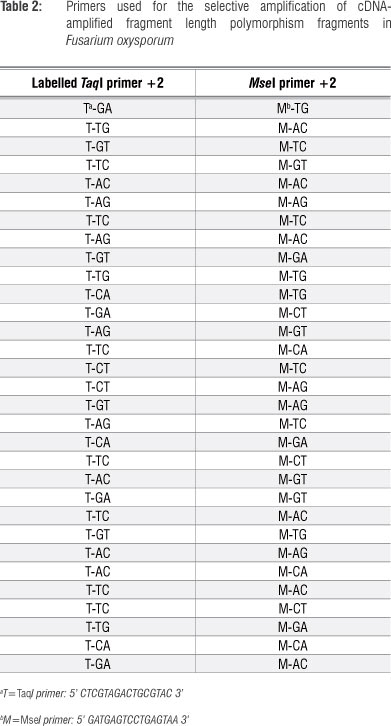
Transcript expression levels of putative pathogenicity genes in F. oxysporum were assessed by cDNA-AFLP analysis. The AFLP® Expression Analysis Kit (LICOR, Lincoln, NE, USA) was employed according to the manufacturer's instructions to determine differential gene expression patterns. Briefly, cDNA was digested with the restriction enzymes TaqI and Msel, followed by ligating adapters using T4 DNA ligase. Pre-selective amplification was performed with TaqI+0/MseI+0 primers, and 31 different TaqI + 2/MseI+2 primer combinations were used during selective amplification (Table 2). Band intensities of differentially expressed fragments on cDNA-AFLP gels were visually assessed and divided into four groups: (1) no transcripts detected (-), (2) low level of transcript abundance (+), (3) moderate level of transcript abundance (++) and (4) high level of transcript abundance (+++). Band intensities correspond to the original expression level.
Isolation of polymorphic fragments and sequence data analysis
After polyacrylamide gels were resolved on the LICOR analyser and scanned with the Odyssey® infrared imaging system (LICOR), unique bands were identified using Quantity One 1-D analysis software (Bio-Rad Laboratories Inc., Hercules, CA, USA). Bands were excised and cloned into a vector with the InsTAclone™ PCR cloning kit (Fermentas) and sequenced in both directions. Vector sequences were manually removed from the raw sequences by Chromas 1.45 (www.technelysium.com.au/chromas.html), while BioEdit Sequence Alignment Editor 7.0.5.3 software36 was used to create a consensus sequence for each individual fragment. The consensus sequences were compared with Fusarium genome sequences of the Broad Institute (http://www.broad.mit.edu/annotation/genome/fusarium_group/Blast.html) and on the National Center for Biotechnology Information (NCBI) database (http://blast.ncbi.nlm.nih.gov/Blast.cgi) for identification. The transcript-derived fragments (TDFs) obtained with cDNA-AFLPs were further characterised using Desktop cDNA Annotation System (dCAS, NIAID, Bethesda, MD, USA).37 Functional groups were defined according to the Munich Information Center for Protein Sequences (MIPS)38 and Gene Ontology (GO)39 databases.
Quantitative analysis of transcript abundance
The transcript abundance of six putative pathogenicity genes identified by cDNA-AFLP in the current study, and eight known pathogenicity genes of F. oxysporum (Table 3), was assessed using a LightCycler 480 instrument (Roche Diagnostics). Five reference genes - elongation factor 1α (TEF), β-tubulin (TUB), isocitrate dehydrogenase (IDH), glucoses-phosphate 1-dehydrogenase (G6DH) and glyceraldehyde 3-phosphate (GAPDH) - were also evaluated (Table 3). Primers for the putative and known pathogenicity and reference genes were designed using Primer3 (Whitehead Institute, MIT, Cambridge, MA, USA) and Netprimer (Premier Biosoft, Palo Alto, CA, USA) (Table 3) and synthesised by Operon Biotechnologies GmbH (Cologne, Germany).
RT-qPCR reactions were performed in 10-uL volumes containing cDNA template (1:10 dilution), 1 of each of the forward and reverse primers and 5 DNA MasterPLUS SYBR Green mix (Roche Diagnostic). The protocol included 10 min at 95 °C followed by 55 cycles of 10 s at 95 °C, 10 s at 57 °C and 10 s at 72 °C. The amplification process was completed by a melting cycle from 55 °C to 95 °C to assess specificity. The fluorescence reading was recorded at 72 °C at the end of the elongation cycles. The PCR products were analysed by electrophoresis on a 2% agarose gel to verify that a single product of the expected size was produced. All reactions were performed in triplicate with three independent biological replicates and a negative control (no template) for all genes. A standard curve was generated by preparing a dilution series (1:10, 1:100 and 1:1000) for each pathogenicity and reference gene. Gene expression stability (M-value) and pairwise variation (V-values) were determined using Genorm.40 Ct values were imported into qbasePLUS (Biogazelle, Ghent, Belgium) for further analysis. The difference in Ct values was determined statistically by one-way analysis of variance, followed by Tukey's post-hoc analysis; p<0.05 was considered statistically significant.
Results
cDNA-AFLP analysis
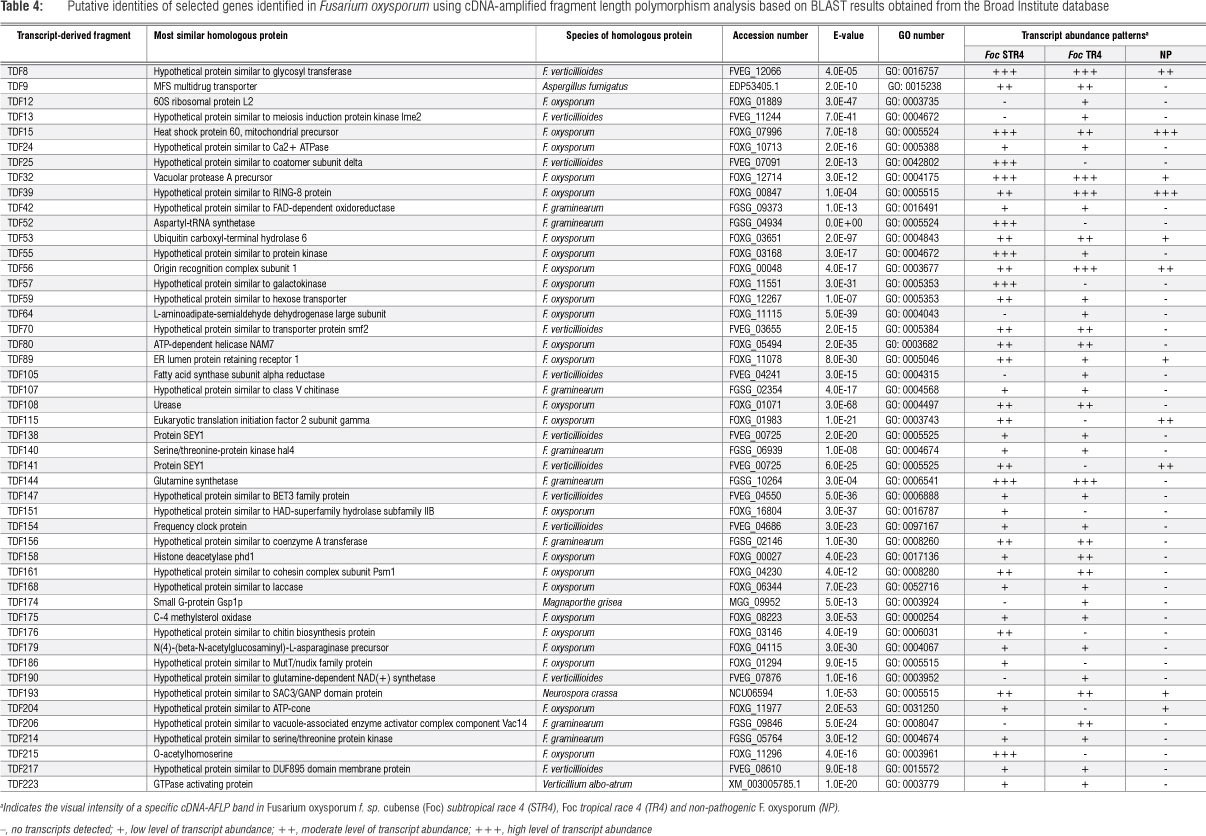
cDNA expression patterns of approximately 3150 transcripts were examined with 31 different TaqI+2/MseI+2 primer combinations. For each primer combination, 63-138 TDFs were visualised and varied from 100 bp to 700 bp with approximately 8% of the TDF showing differential expression. cDNA-AFLP analysis allowed the identification and isolation of 229 differentially expressed TDFs of between 103 bp and 546 bp in size (Table 4). The TDFs were classified into these functional categories: hypothetical proteins from Fusarium (90) and other fungal species (6), energy metabolism (13), transport (13), cell division and growth (11), protein turnover (8), cell signalling (9),- lipid or fatty acid metabolism (5), transcription and translation factors (6), and those with no significant homology (68) (Figure 1). BLAST analysis with an rRNA operon showed that only one TDF (0.4%) had homology to rRNA. Several TDFs represent genes with numerous functions, including pathogenicity. These TDFs included the putative chitinase class V (chsV) (TDF107), GTPase activating protein (rhoI) (TDF223), Major Facilitator Superfamily (MFS) multidrug transporter (TDF9), laccase (lcc) (TDF168), Ca2+ ATPase (TDF24) and serine/ threonine protein kinase (ste12) (TDF214) genes (Table 4). The TDFs corresponding to chsV, rhoI, lcc, Ca2+ ATPase, and ste12 showed low intensity levels in Foc STR4 and Foc TR4 compared with non-pathogenic F. oxysporum where transcripts were not visually detected. The TDF representing the MFS multidrug transporter gene displayed moderate intensity in Foc STR4 and Foc TR4 compared with undetectable levels in non-pathogenic F. oxysporum (Figure 2a).
Several different transcript abundance patterns were detected during cDNA-AFLP gel analysis (Table 4). In the first pattern, high transcript abundance was detected in Foc STR4 with no transcripts detected in Foc TR4 or non-pathogenic F. oxysporum. Examples showing high abundance include TDFs corresponding to the aspartyl-tRNA synthetase (TDF52), galactokinase (TDF57) and O-acetylhomoserine (TDF215). The second transcript abundance pattern showed an increase in transcripts in Foc TR4 with no transcripts detected in Foc STR4 or non-pathogenic F. oxysporum. TDFs that exhibited this pattern were 60S ribosomal protein L2 (TDF12), meiosis induction protein (TDF13), L-aminoadipate semialdehyde dehydrogenase large subunit (TDF64), fatty acid synthase subunit alpha reductase (TDF105), small G-protein Gsp1p (TDF174) and glutamine-dependent NAD+ synthetase (TDF190). In the third pattern, transcripts were detected in Foc STR4 and Foc TR4 with no detection in non-pathogenic F. oxysporum. Examples of these transcripts include-chsV (TDF107), Ca2+ ATPase (TDF24), FAD-dependent oxidoreductase (TDF42), ste12 (TDF214), GTPase activating protein (TDF223) and lcc (TDF168). Other transcript abundance patterns included transcript presence in Foc STR4 and the non-pathogenic strains, with no transcripts detected in Foc TR4. These transcripts included eukaryotic translation initiation factor 2 subunit gamma (TDF115) and ATP-cone (TDF204). Another pattern displayed low levels of transcript abundance in Foc STR4 with high transcript abundance in Foc TR4 and non-pathogenic F. oxysporum.
Quantitative verification of cDNA-AFLP
Five reference genes (Table 3) were evaluated for stable expression. The average pairwise variation (V-value) calculated for IDH, G6DH and GAPDH was 0.113, with TEF and TUB showing less stable expression levels (V=0.225). As a result, the reference genes IDH, G6DH and GAPDH were used to normalise the data as suggested by Vandesompele et al.40
The relative transcript abundance of six genes measured by cDNA-AFLP analysis - encoding a MFS multidrug transporter (TDF9), a L-aminoadipate-semialdehyde dehydrogenase large subunit (TDF64), an aspartyl-tRNA synthetase (TDF52), a chsV (TDF107), a ste12 (TDF214) and rhoI (TDF223) - was compared with results obtained by qRT-PCR (Figure 2). Both cDNA-AFLP and qRT-PCR analyses showed an increased abundance of the MFS multidrug transporter gene in Foc STR4 and Foc TR4 when compared with non-pathogenic F. oxysporum (Figure 2a). When the abundance levels of L-aminoadipate-semialdehyde dehydrogenase large subunit were compared, the cDNA-AFLP analysis demonstrated that the transcript was present in Foc TR4 but absent in the transcript in Foc STR4 and the non-pathogenic F. oxysporum. The qRT-PCR data showed similar levels of transcript abundance in Foc TR4, Foc STR4 and the non-pathogenic F. oxysporum (Figure 2b). cDNA-AFLP analyses showed an increased abundance of transcripts of aspartyl-tRNA synthetase in Foc STR4 compared with Foc TR4 and the non-pathogenic F. oxysporum, while qRT-PCR revealed similar transcript levels among the different isolates (Figure 2c). An increase in transcript abundance of chsV was found in Foc STR4 and Foc TR4 when compared with the non-pathogenic F. oxysporum using both cDNA-AFLP analysis and qRT-PCR (Figure 2d). Transcript abundance profiles determined for ste12 by cDNA-AFLP and qRT-PCR were similar, and showed an increase in Foc STR4 and Foc TR4 compared with the non-pathogenic F. oxysporum (Figure 2e). In the case of rhoI, cDNA-AFLP analysis showed an increase in the number of transcripts in Foc race 4 compared with the non-pathogenic F. oxysporum (Figure 2f). However, qRT-PCR showed a higher number of transcripts in Foc STR4 than in Foc TR4 and the non-pathogenic F. oxysporum. Thus, the transcript abundance patterns measured by qRT-PCR were similar to those measured for the corresponding TDFs analysed using cDNA-AFLP.
Transcript abundance of known pathogenicity genes using qRT-PCR
Foc STR4 and Foc TR4 expressed the pathogenicity genes snf (Figure 3a), frp1 (Figure 3b) and cyp55 (Figure 3c) at significantly higher levels than non-pathogenic F. oxysporum. The transcript abundance of snf was 2.6-fold higher in Foc STR4 than in the non-pathogenic F. oxysporum. The transcript abundance levels of frp1 were lower in non-pathogenic F. oxysporum isolates than in pathogenic Foc STR4 and Foc TR4 isolates, by 3.6-fold and 2.5-fold, respectively. Snf and frp1 are involved in the degradation of plant cell walls.17,41Cyp55 had a 1.6-fold higher expression in Foc TR4 compared with Foc STR4, but this difference was not statistically significant. Cyp55 is a nitric oxide reductase involved in the nitrogen response pathway, which is fundamental for pathogenicity.
Fmk1 is responsible for maintaining fungal cell wall architecture and signalling. Fmk1 was expressed significantly more in Foc STR4 than in either Foc TR4 or non-pathogenic F. oxysporum (Figure 3d). Fmk1 expression was 2.9-fold higher in Foc STR4 than in the non-pathogenic-F. oxysporum (Figure 3d). In addition, transcript abundance of fmk1 was a significant 2.1-fold higher in Foc STR4 than in Foc TR4. Expression of the chloride channel (clc) gene, which controls laccase activity, was also significantly higher in Foc STR4 than in the non-pathogenic F. oxysporum (Figure 3e). In contrast, fow2, the fungal gene involved in regulating pathogenicity-related transcription, was expressed significantly more in Foc TR4 than in the non-pathogenic F. oxysporum, but not significantly more than in Foc STR4 (Figure 3f). There were no significant differences observed in the transcript abundance profiles of the arginine biosynthesis gene (arg1) (Figure 3g) or mitochondrial protein gene (fow1) (Figure 3h).
Discussion and conclusion
The transcriptomes of Foc STR4, Foc TR4 and non-pathogenic F. oxysporum isolates on MM (without carbon source) were visually detected with cDNA-AFLP More than 3000 TDFs were detected, of which 8% showed differential expression patterns. A total of 3% of these TDFs were putatively involved in pathogenicity. Several fungal gene transcripts that have previously been associated with pathogenicity in other fungal organisms have been identified for the first time in the banana pathogen Foc. These genes include chsV, rhoI, MFS multidrug transporter and stel2. In addition, the genes snf, frp1 and cyp55, which result in diseases of crops other than banana, were more abundantly expressed in Foc STR4 and Foc TR4 than in non-pathogenic F. oxysporum.
The genes chsV and rhoI have previously been associated with pathogenicity in Fol on tomato.42,43ChsV restricts toxic substances- produced by the plant for defence against pathogens,42 whereas rho1 plays a role in preventing the host plant from recognising the pathogen.43 Both genes, therefore, protect the pathogen against the host's defence response. Because chsV and rho1 showed higher transcript abundances in Foc STR4 and Foc TR4 than in the non-pathogen, we hypothesise that Foc expresses these genes when infecting the xylem vessels of Cavendish bananas to avoid the plant's defence responses.
The transcript abundance of the MFS multidrug transporter was fivefold higher in pathogenic Foc than in the non-pathogen. This family of transporters regulate the movement of sugars, Krebs-cycle metabolites, phosphorylated glycolytic intermediates, amino acids, peptides, osmolites, iron-siderophores, nucleosides and organic and inorganic anions and cations.44 In addition, MFS transporters have been linked to fungal pathogenicity by avoiding toxic compounds produced by the pathogen, or by protection against plant defence compounds.45 MFS transpor ter gene in the ascomycete Verticillium dahlia - a vascular pathogen - is essential for pathogenicity on lettuce plants.46 With a significantly higher transcript abundance of the MFS multidrug transporter, Foc STR4 and Foc TR4 may possibly protect themselves from toxic substances produced by the plant during defence.
The transcription factor ste12 is important during fungal infection of plant roots where it regulates genes involved in the MAPK cascade.20,47 In a study by Garcia-Sanchez et al.22, a ste12-like gene, fost12, showed an increased expression after 12-24 h of infection of bean plants by F.-oxysporum f. sp. phaseoli. A significant increase in transcript abundance of ste12 in Foc STR4 and Foc TR4 can activate the MAPK signalling pathways, thereby increasing CWDE during the infection process. A second transcription factor, fow2,21 a Zn(II)2Cys6-type transcription regulator involved in pathogenicity in F. oxysporum f. sp. melonis, was significantly higher in Foc TR4 than in the non-pathogen, but there was no significant difference between Foc STR4 and the non-pathogen. Because Foc TR4 is a more virulent pathogen than Foc STR4, fow2 may assist in the more rapid invasion of root tissue or may be differentially regulated in Foc STR4 and Foc TR4.
Two well-studied pathogenicity genes previously isolated from F. oxysporum that regulate the abundance of CWDEs are snf and frp1.16,17,41 Both snf and frp1 were significantly higher in Foc STR4 and Foc TR4 than in the non-pathogen, which suggests that these genes are important for the Fusarium wilt pathogen to enter the host xylem tissue. As an endophyte, the non-pathogenic F. oxysporum isolates are usually restricted to the root cortex, and do not enter the xylem vessels.48 In contrast, Foc STR4 and Foc TR4 both degrade the xylem cell walls to enter the vascular tissue.
Pathogenicity and cell wall degradation are affected by the enhanced expression of MAP kinases in several fungi, for example Fol,15 Fusarium graminearum,49 Magnaporthe grisea50 and Ustilago maydis51. In Fol, fmk1 also aids in root attachment, penetration, invasive growth and increased CWDE activity.15 The significant increase in fmk1 in Foc STR4 and non-significant increase in Foc TR4 compared with non-pathogenic F. oxysporum may explain pathogenesis in the banana Fusarium wilt pathogen, that is by accelerating invasive growth as in other Fusarium species.15,52 Pathogenic Foc isolates are able to colonise both the cortex and the xylem tissue, resulting in severe discoloration of the corm and blocking of the vascular bundles. In contrast, the non-pathogenic strains are restricted to the root cortex, which results in no symptoms developing. The reason that fmk1 did not show a significant increase in transcript abundance in Foc TR4 is not certain, but one possible explanation could be that fmk1 transcripts amplified during pathogenicity- at earlier time points were not sampled in this study. Genes expressed during the early time points are either translated into proteins or the RNA is degraded as the half-life of RNA is short and therefore the RNA cannot be detected at later time points.
Cyp55 was more abundant in Foc race 4 than in non-pathogenic F. oxysporum. This gene plays a role in the ability to regulate the nitrogen response pathway, which is essential for pathogenicity.53Cyp55, a cytochrome P450 gene involved in the reduction of nitric oxide in F. oxysporum, was first characterised by Kizawa et al.54 The cyp55 gene of F. oxysporum f. sp. vasinfectum has been previously reported to be highly expressed in cotton plants following root inoculation.55
Laccases serve as virulence factors in fungal pathogens by playing a role in pigmentation, appressorium formation and protection against toxic phytoalexins.56 qRT-PCR analysis in this study revealed a significant increase in clc transcripts in Foc STR4 compared with the non-pathogen. In Fol, mutations of lcc1, lcc3 and lcc5 had no effect on pathogenicity in tomato plants.57 As six lcc genes have been identified in F. oxysporum, Cañero and Roncero28 suggested that a mutation in one of them may not necessarily prevent laccase activity, as the other isozymes fulfil their role.57 However, clc mutants showed a decrease in laccase activity with a reduction in virulence to tomato seedlings.28 Increased clc expression and the role of laccases and chloride transport in the banana Fusarium wilt pathogen may be important pathogenicity determinants.
The cDNA-AFLP technique was useful in differentiating the transcript abundance of genes present in Foc race 4 and non-pathogenic F. oxysporum. However, DNA sequence differences could result in the absence or presence of a TDF not necessarily implicating differential expression. To minimise these single nucleotide polymorphisms, nine isolates from different geographical regions were combined for each of the Foc STR4, Foc TR4 and non-pathogenic F. oxysporum fungal samples. Most of the gene expression patterns measured by cDNA-AFLPs were confirmed by qRT-PCR analyses. However, next-generation-
DNA and RNA sequencing could provide significantly better results for identifying pathogenicity genes in Foc, both in STR4 and TR4, especially once the full genome sequence of the Fusarium wilt fungus becomes available. Comparison of the Foc genome with that of other forma speciales of F. oxysporum will elucidate the ability of Foc to infect banana roots. Virulence factors can be studied when the genomes of Foc TR4, a more virulent pathogen, are compared with Foc STR4. Furthermore, the function of putative pathogenicity genes during infection should be investigated by gene knockout studies and RNAi silencing. Knockout mutants would help to identify additional genes required for pathogenicity in Foc race 4.
An in-depth understanding of pathogenicity in Foc is required if novel approaches to disease management are to be developed. We have identified several transcripts in Foc race 4 that are more abundant in the pathogenic strains compared with the non-pathogens. Many of these TDFs have been shown to play a role in host infection and colonisation by other Fusarium spp. These TDFs encode for CWDEs and proteins involved in avoiding toxic substances produced during plant defence. To establish function, knockout mutants of genes underlying these transcripts need to be generated, and the role of genes such as chsV, rhoI, MFS multidrug transporter, ste12, snf, frp1, cyp55 and fmk1 needs further investigation. With the rapid advancement in molecular techniques in recent years, new strategies for increasing plant resistance against specific Fusarium wilt pathogens can be generated by exploiting the molecular and cellular bases of pathogenicity.
Acknowledgements
This research was funded by The Human Resources for Industry Programme (THRIP), an initiative of the Department of Trade and Industry (DTI); the National Research Foundation Thuthuka programme; and the Banana Growers Association of South Africa.
Authors' contributions
R.S. performed the experiments; A.V., A.A.M., N.v.d.B. and R.S. were involved in the experimental design and data analysis; and all authors contributed in writing the manuscript.
References
1. Michielse CB, Rep M. Pathogen profile update: Fusarium oxysporum. Mol Plant Pathol. 2009;10(3):311-324. http://dx.doi.org/10.1111/j.1364-3703.2009.00538.x
2. Gordon TR, Martyn RD. The evolutionary biology of Fusarium oxysporum. Annu Rev Phytopathol. 1997;35(1):111-128. http://dx.doi.org/10.1146/annurev.phyto.35.1.111
3. Ma L-J, Van der Does HC, Borkovich KA, Coleman JJ, Daboussi M-J, Di Pietro A, et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature. 2010;464(7287):367-373. http://dx.doi.org/10.1038/nature08850
4. Rep M, Kistler HC. The genomic organization of plant pathogenicity in Fusarium species. Curr Opin Plant Biol. 2010;13(4):420-426. http://dx.doi.org/10.1016/j.pbi.2010.04.004
5. Idnurm A, Howlett BJ. Pathogenicity genes of phytopathogenic fungi. Mol Plant Pathol. 2001;2(4):241-255. http://dx.doi.org/10.1046/j.1464-6722.2001.00070.x
6. Mõbius N, Hertweck C. Fungal phytotoxins as mediators of virulence. Curr Opin Plant Biol. 2009;12(4):390-398. http://dx.doi.org/10.1016/j.pbi.2009.06.004
7. Werner S, Sugui JA, Steinberg G, Deising HB. A chitin synthase with a myosin-like motor domain is essential for hyphal growth, appressorium differentiation, and pathogenicity of the maize anthracnose fungus Colletotrichum graminicola. Mol Plant Microbe In. 2007;20(12):1555-1567. http://dx.doi.org/10.1094/MPMI-20-12-1555
8. Chi M-H, Park S-Y, Kim S, Lee Y-H. A novel pathogenicity gene is required in the rice blast fungus to suppress the basal defenses of the host. PLoSPathog. 2009;5(4):e1000401. http://dx.doi.org/10.1371/journal.ppat.1000401
9. De Wit PJGM, Mehrabi R, Van Den Burg HA, Stergiopoulos I. Fungal effector proteins: Past, present and future. Mol Plant Pathol. 2009;10(6):735-747. http://dx.doi.org/10.1111/j.1364-3703.2009.00591.x
10. Di Pietro A, Roncero MIG. Cloning, expression, and role in pathogenicity of pg1 encoding the major extracellular endopolygalacturonase of the vascular wilt pathogen Fusarium oxysporum. Mol Plant Microbe In. 1998;11(2):91-98. http://dx.doi.org/10.1094/MPMI.1998.11.2.91 [ Links ]
11. García-Maceira FI, Di Pietro A, Roncero MIG. Cloning and disruption of pgx4 encoding an in planta expressed exopolygalacturonase from Fusarium oxysporum. Mol Plant Microbe In. 2000;13(4):359-365. http://dx.doi.org/10.1094/MPMI.2000.13.4.359
12. Huertas-González MD, Ruiz-Roldán MC, García Maceira FI, Roncero MIG, Di Pietro A. Cloning and characterization of pl1 encoding an in planta-secreted pectate lyase of Fusarium oxysporum. Curr Genet. 1999;35(1):36-40. http:// dx.doi.org/10.1007/s002940050430
13. Roldán-Arjona T, Pérez-Espinosa A, Ruiz-Rubio M. Tomatinase from Fusarium oxysporum f. sp. lycopersici defines a new class of saponinases. Mol Plant Microbe In. 1999;12(10):852-861. http://dx.doi.org/10.1094/MPMI.1999.12.10.852
14. Ruiz-Roldán MC, Di Pietro A, Huertas-González MD, Roncero MIG. Two xylanase genes of the vascular wilt pathogen Fusarium oxysporum are differentially expressed during infection of tomato plants. Mol Gen Genet. 1999;261(3):530-536. http://dx.doi.org/10.1007/s004380050997
15. Di Pietro A, García-Maceira FI, Méglecz E, Roncero MIG. A MAP kinase of the vascular wilt fungus Fusarium oxysporum is essential for root penetration and pathogenesis. Mol Microbiol. 2001;39(5):1140-1152. http://dx.doi.org/10.1111/j.1365-2958.2001.02307.x
16. Duyvesteijn RGE, Van Wijk R, Boer Y, Rep M, Cornelissen BJC, Haring MA. Frp1 is a Fusarium oxysporum F-box protein required for pathogenicity on tomato. Mol Microbiol. 2005;57(4):1051-1063. http://dx.doi.org/10.1111/j.1365-2958.2005.04751.x
17. Ospina-Giraldo MD, Mullins E, Kang S. Loss of function of the Fusarium oxysporum SNF1 gene reduces virulence on cabbage and Arabidopsis. Curr Genet.. 2003;44:49-57. http://dx.doi.org/10.1007/s00294-003-0419-y [ Links ]
18. Jain S, Akiyama K, Kan T, Ohguchi T, Takata R. The G protein β subunit FGB1 regulates development and pathogenicity in Fusarium oxysporum. Curr Genet.. 2003;43(2):79-86.
19. Jain S, Akiyama K, Mae K, Ohguchi T, Takata R. Targeted disruption of a G protein a subunit gene results in reduced pathogenicity in Fusarium oxysporum. Curr Genet.. 2002;41(6):407-413. http://dx.doi.org/10.1007/s00294-002-0322-y
20. Rispail N, Di Pietro A. Fusarium oxysporum ste12 controls invasive growth and virulence downstream of the Fmk1 MAPK cascade. Mol Plant Microbe In. 2009;22(7):830-839. http://dx.doi.org/10.1094/MPMI-22-7-0830
21. Imazaki I, Kurahashi M, Iida Y Tsuge T. Fow2, a Zn(II)2Cys6-type transcription regulator, controls plant infection of the vascular wilt fungus Fusarium oxysporum. Mol Microbiol. 2007;63(3):737-753. http://dx.doi.org/10.1111/j.1365-2958.2006.05554.x
22. Garcia-Sanchez AM, Martin-Rodrigues N, Ramos B, De Vega-Bartol JJ, Perlin MH, Diaz-Minguez JM. Fost12, the Fusarium oxysporum homolog of the transcription factor Ste12, is upregulated during plant infection and required for virulence. Fungal Genet Biol. 2010;47:216-225. http://dx.doi.org/10.1016/j.fgb.2009.11.006
23. Ploetz RC, Pegg KG. Fungal diseases of the root, corm and pseudostem: Fusarium wilt. In: Jones DR, editor. Diseases of banana, abacá and enset. London: CAB International; 2000. p. 143-159. http://dx.doi.org/10.1094/PHP-2005-1221-01-RV [ Links ]
24. Ploetz RC. Panama disease: An old nemesis rears its ugly head. Part 1. The beginnings of the banana export trades. Plant Health Progress. 2005; doi:10.1094/PHP-2005-1221-01-RV. [ Links ]
25. Belgrove A, Steinberg C, Viljoen A. Evaluation of non-pathogenic Fusarium oxysporum and Pseudomonas fluorescens for Panama disease control. Plant Dis. 2011;95(8):951-959. http://dx.doi.org/10.1094/PDIS-06-10-0409
26. Thangavelu R, Jayanthi A. RFLP analysis of rDNA-ITS regions of native non-pathogenic Fusarium oxysporum isolates and their field evaluation for the suppression of Fusarium wilt disease of banana. Australas Plant Path. 2009;38(1):13-21. http://dx.doi.org/10.1071/AP08071
27. Athman SY Dubois T, Viljoen A, Labuschagne N, Coyne D, Ragama P et al. In vitro antagonism of endophytic Fusarium oxysporum isolates against the burrowing nematode Radopholus similis. Nematology. 2006;8(4):627-636. http://dx.doi.org/10.1163/156854106778613976
28. Cañero DC, Roncero MIG. Influence of the chloride channel of Fusarium oxysporum on extracellular laccase activity and virulence on tomato plants. Microbiology. 2008;154:1474-1481. http://dx.doi.org/10.1099/mic.0.2007/015388-0
29. Inoue I, Namiki F, Tsuge T. Plant colonization by the vascular wilt fungus Fusarium oxysporum requires FOW1, a gene encoding a mitochondrial protein. Plant Cell. 2002;14:1869-1883. http://dx.doi.org/10.1105/tpc.002576 [ Links ]
30. Namiki F, Matsunaga M, Okuda M, Inoue I, Nishi K, Fujita Y et al. Mutation of an arginine biosynthesis gene causes reduced pathogenicity in Fusarium oxysporum f. sp. melonis. Mol Plant Microbe In. 2001;14(4):580-584. http://dx.doi.org/10.1094/MPML2001.14A580 [ Links ]
31. Tomura D, Obika K, Fukamizu A, Shoun H. Nitric oxide reductase cytochrome P-450 gene, CYP55, of the fungus Fusarium oxysporum containing a potential binding-site for FNR, the transcription factor involved in the regulation of anaerobic growth of Escherichia coli. J Biochem. 1994;116(1):88-94. [ Links ]
32. Nel B, Steinberg C, Labuschagne N, Viljoen A. Isolation and characterization of nonpathogenic Fusarium oxysporum isolates from the rhizosphere of healthy banana plants. Plant Pathol. 2006;55:207-216. http://dx.doi.org/10.1111/j.1365-3059.2006.01343.x [ Links ]
33. Van den Berg N, Berger DK, Hein I, Birch PRJ, Wingfield MJ, Viljoen A. Tolerance in banana to Fusarium wilt is associated with early up-regulation of cell wall-strengthening genes in the roots. Mol Plant Pathol. 2007;8(3):333- 341. http://dx.doi.org/10.1111/j.1364-3703.2007.00389.x
34. Trail F, Xu J-R, Miguel PS, Halgren RG, Corby Kistler H. Analysis of expressed sequence tags from Gibberella zeae (anamorph Fusarium graminearum). Fungal Genet Biol. 2003;38(2):187-197. http://dx.doi.org/10.1016/S1087-1845(02)00529-7 [ Links ]
35. O'Donnell K, Corby Kistler H, Cigelnik E, Ploetz R. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc Natl Acad Sci USA. 1998;95:2044-2049. http://dx.doi.org/10.1073/pnas.95.5.2044 [ Links ]
36. Hall TA. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acid S. 1999;41:95-98.
37. Guo Y Ribeiro JM, Anderson JM, Bour S. dCAS: A desktop application for cDNA sequence annotation. Bioinformatics. 2009;25(9):1195-1196. http://dx.doi.org/10.1093/bioinformatics/btp129 [ Links ]
38. Ruepp A, Zollner A, Maier D, Albermann K, Hani J, Mokrejs M, et al. The FunCat, a functional annotation scheme for systematic classification of proteins from whole genomes. Nucleic Acids Res. 2004 October 14, 2004;32(18):5539-5545. [ Links ]
39. The Gene Ontology Consortium. Gene ontology: Tool for the unification of biology. Nature Genetics. 2000;25(1):25-29. http://dx.doi.org/10.1038/75556
40. Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A, et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002;3:research0034.
41. Jonkers W, Rodrigues CDA, Rep M. Impaired colonization and infection of tomato roots by the Afrpl mutant of Fusarium oxysporum correlates with reduced CWDE gene expression. Mol Plant Microbe In. 2009;22(5):507-518. http://dx.doi.org/10.1094/MPMI-22-5-0507
42. Madrid MP Di Pietro A, Roncero MIG. Class V chitin synthase determines pathogenesis in the vascular wilt fungus Fusarium oxysporum and mediates resistance to plant defence compounds. Mol Microbiol. 2003;47(1):257- 266. http://dx.doi.org/10.1046/j.1365-2958.2003.03299.x
43. Martínez-Rocha AL, Roncero MIG, López-Ramirez A, Mariné M, Guarro J, Martínez-Cadena G, et al. Rho1 has distinct functions in morphogenesis, cell wall biosynthesis and virulence of Fusarium oxysporum. Cell Microbiol. 2008;10(6):1339-1351. http://dx.doi.org/10.1111/j.1462-5822.2008.01130.x
44. Stergiopoulos I, Zwiers L-H, De Waard MA. Secretion of natural and synthetic toxic compounds from filamentous fungi by membrane transporters of the ATP-binding cassette and major facilitator superfamily. Eur J Plant Pathol. 2002;108(7):719-734. http://dx.doi.org/10.1023/A:1020604716500
45. Del Sorbo G, Schoonbeek H-J, De Waard MA. Fungal transporters involved in efflux of natural toxic compounds and fungicides. Fungal Genet Biol. 2000;30(1):1-15. http://dx.doi.org/10.1006/fgbi.2000.1206
46. Maruthachalam K, Klosterman S, Kang S, Hayes R, Subbarao K. Identification of pathogenicity-related genes in the vascular wilt fungus Verticillium dahliae by Agrobacterium tumefaciens-mediated T-DNA insertional mutagenesis. Mol Biotechnol. 2011;49(3):209-221. http://dx.doi.org/10.1007/s12033-011-9392-8
47. Rispail N, Di Pietro A. The homeodomain transcription factor Ste12: Connecting fungal MAPK signaling to plant pathogenicity. Commun Integr Biol. 2010;3(4):327-332. http://dx.doi.org/10.4161/cib.3.4.11908 [ Links ]
48. MacHardy WE, Beckman CH. Vascular wilt Fusaria: Infection and pathogenesis. In: Nelson PE, Toussoun TA, Cook RJ, editors. Fusarium: Diseases, biology, and taxonomy. London: Pennsylvania State University Press; 1981. p. 365-390. [ Links ]
49. Jenczmionka NJ, Schafer W. The Gpmk1 MAP kinase of Fusarium graminearum regulates the induction of specific secreted enzymes. Curr Genet. 2005;47(1):29-36. http://dx.doi.org/10.1007/s00294-004-0547-z
50. Xu J-R, Hamer JE. MAP kinase and cAMP signaling regulate infection structure formation and pathogenic growth in the rice blast fungus Magnaporthe grisea. Gene Dev. 1996;10:2696-2706. http://dx.doi.org/10.1101/gad.10.21.2696
51. Mayorga ME, Gold SE. A MAP kinase encoded by the ubc3 gene of Ustilago maydis is required for filamentous growth and full virulence. Mol Microbiol. 1999;34(3):485-497. http://dx.doi.org/10.1046/j.1365-2958.1999.01610.x
52. Zhang Y Choi Y-E, Zou X, Xu J-R. The FvMK1 mitogen-activated protein kinase gene regulates conidiation, pathogenesis, and fumonisin production in Fusarium verticillioides. Fungal Genet Biol. 2011;48(2):71-79. http://dx.doi.org/10.1016/j.fgb.2010.09.004
53. Lopez-Berges MS, Rispail N, Prados-Rosales RC, Di Pietro A. A nitrogen response pathway regulates virulence functions in Fusarium oxysporum via the protein kinase TOR and the bZIP protein MeaB. Plant Cell. 2010;22(7):2459- 2475. http://dx.doi.org/10.1105/tpc.110.075937 [ Links ]
54. Kizawa H, Tomura D, Oda M, Fukamizu A, Hoshino T, Gotoh O, et al. Nucleotide sequence of the unique nitrate/nitrite-inducible cytochrome P-450 cDNA from Fusarium oxysporum. J Biol Chem. 1991;266(16):10632-10637.
55. McFadden HG, Wilson IW, Chapple RM, Dowd C. Fusarium wilt (Fusarium oxysporum f. sp. vasinfectum) genes expressed during infection of cotton (Gossypium hirsutum). Mol Plant Pathol. 2006;7(2):87-101. http://dx.doi.org/10.1111/j.1364-3703.2006.00327.x [ Links ]
56. Mayer AM, Staples RC. Laccase: New functions for an old enzyme. Phytochemistry. 2002;60:551-565. http://dx.doi.org/10.1016/S0031-9422(02)00171-1
57. Cañero DC, Roncero MIG. Functional analyses of laccase genes from Fusarium oxysporum. Phytopathology. 2008;98:509-518. http://dx.doi.org/10.1094/PHYTO-98-5-0509
 Correspondence:
Correspondence:
René Sutherland
FABI, University of Pretoria
Private Bag X20, Hatfield 0028, South Africa
EMAIL: rene.sutherland@fabi.up.ac.za
Received: 30 July 2012
Revised: 21 Sep. 2012
Accepted: 05 Nov. 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}