










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.109 no.5-6 Pretoria Jan. 2013
RESEARCH ARTICLE
Hypercarnivory, durophagy or generalised carnivory in the Mio-Pliocene hyaenids of South Africa?
Adam Hartstone-RoseI; Deano D. StynderII
IDepartment of Cell Biology & Anatomy, University of South Carolina School of Medicine, Columbia, SC, USA
IIDepartment of Archaeology, University of Cape Town, Cape Town, South Africa
ABSTRACT
Carnivorans, the members of the order Carnívora, exhibit wide dietary diversity - from overwhelmingly herbivorous species (like the giant and red pandas) to species that specialise in the consumption of flesh (like the hypercarnivorous felids). Throughout the evolution of this order, many craniodental forms have emerged and gone extinct - notably the sabretooth felids that existed until the late Pleistocene. However, one carnivoran lineage, remarkable for its extreme masticatory adaptations, persists - the bone-cracking hyaenids. Three of the four extant members of this family (Crocuta crocuta, Hyaena hyaena and Parahyaena brunnea) are among the most durophagous mammals to have ever lived. The fourth extant hyaenid - the aardwolf (Proteles cristatus) - also exhibits impressive, although wholly different, masticatory adaptations as one of the most derived mammalian insectivores. How and when did the level of durophagy evident in extant bone-cracking hyenas evolve, and how do Mio-Pliocene hyenas compare to the extant members of the order in terms of their own dietary specialisations? An examination of the premolars of the Mio-Pliocene hyaenids from Langebaanweg, South Africa suggests that modern levels of durophagy appeared relatively recently. Results from an analysis of dental radii-of-curvature and premolar intercuspid notches suggest that these hyenas were neither bone crackers nor flesh specialists, but were dietary generalists.
Keywords: hyena; Carnivora; Langebaanweg; Miocene; osteophagy
Introduction
The order Carnivora contains taxa that consume nearly the entire range of mammalian foods, from hypercarnivorous flesh specialists (e.g. the members of the cat family) to species whose diets consist nearly entirely of bamboo (e.g. the giant and red pandas). Many of the smaller members of the order (e.g. many of the mongooses) subsist substantially on insects. Other carnivorans (e.g. kinkajous and binturongs) are substantially frugivorous. Most species in the order exist in some generalised omnivorous space (exemplified by species like raccoons and most bears).
The four extant species in the family Hyaenidae are among the most dentally derived carnivorans. At 8-12 kg,1 the smallest species in the group, the 'aardwolf' (Proteles cristatus), is craniodentally adapted for a diet consisting almost entirely of termites.2,3 Although its fossil record is poor,1,4,5 the aardwolf lineage probably diverged from the civet-like early hyaenids.6,7 While the aardwolf is more craniodentally derived than almost any other recent mammal, the craniodental adaptations of the other three extant hyaenids are equally impressive. These three monospecific genera (Crocuta, Hyaena and Parahyaena) are among the most durophagous mammals alive today. Indeed, only the extinct borophagine canids, borhyaenids and creodonts were comparable in their bone-cracking adaptations.4,8,9 With their ability to consume nearly all carcass elements, the modern hyenas are excellent scavengers, and some are also highly efficient hunters.10 These bone-cracking species have skulls, jaws and teeth that are powerfully built.4,11,12 Their masticatory muscles are large and their premolars - where most of the bone-cracking occurs13 - are broad and stoutly built to transmit large forces.12
Hyenas diverged from the stem feliform in the Oligocene and transitioned through six ecomorph groups from civet-like insectivores/omnivores through generalised jackal-like meat and small bone eaters to the fully developed modern bone crushers.4 This middle phase is well represented by the four hyaenid species that have been described from the South African fossil site of Langebaanweg (LBW) E Quarry14-17 (Figure 1): Chasmaporthetes australis, Hyaenictitherium namaquensis, Hyaenictis hendeyi and Ikelohyaena abronia.4,18
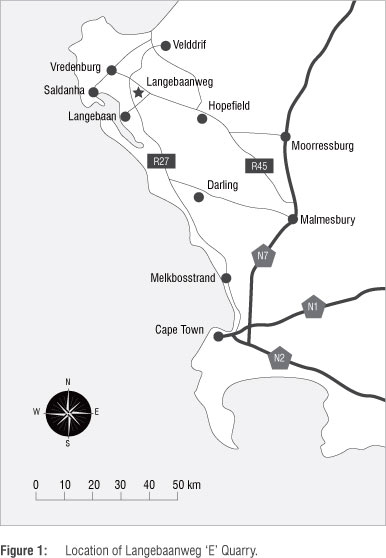
Langebaanweg E Quarry is one of the most significant Early Pliocene fossil-bearing sites in Africa. Originally discovered during phosphate mining operations in the early half of the last century, LBW has produced a great diversity of terrestrial and marine animal taxa.14,15,19-33 Located at what some consider to have been the then mouth of the Berg River,19,20,34-36 LBW's geological history likely involved repeated marine transgression and river derived sedimentation events35,37 which explains its mix of terrestrial and marine fossils. Fossils are primarily derived from two members of the local Varswater Formation, namely the Langeberg Quartzose Sand Member (LQSM) and the Muishondfontein Pelletal Phosphate Member (MPPM).20 The LQSM is construed as a floodplain or estuarine deposit of local origin, while the MPPM, which overlays it in places, is a river channel deposit largely derived from upstream.20 Both levels are presumed to have been laid down at more-or-less the same time around 5.2 Ma.
The fossilised remains of hyenas occur in both fossil-bearing members of the Varswater formation, with two species occurring in the LQSM and MPPM (C. australis and I. abronia) and one each in the LQSM (H. hendeyi) and MPPM (H. namaquensis).30 This distribution pattern, while intriguing, is probably not representative of species distribution in the LBW palaeoenvironment. Terrestrial carnivores are generally not as tied to specific types of environments as, for example, herbivores are and the absence of H. hendeyi in the MPPM and H. namaquensis in the LQSM is most likely attributable to taphonomic or sampling biases.38,39 It is thus not unreasonable to conclude that all four species were probably sympatric in the region of the estuary and most probably further inland too.
The LBW hyaenids, unlike their modern bone-cracking counterparts, were poorly adapted to bone cracking.1-18,40,41Ikelohyaena abronia, which has traditionally been regarded as the most durophagous of the LBW species,4,22,38 possessed derived features in skull stress distribution and levels of strain energy similar to those of Crocuta crocuta, but importantly lacked the bite force of the extant species.42 This fossil species belonged to a clade of early or transitional bone-cracking hyenas that also included other early genera such as Palinhyaena, Belbus, Hyaenid sp. E and Leecyaena.4 Chasmaporthetes australis and H. hendeyi on the other hand, fell within a clade of hypercarnivorous hyenas that also included the extinct genus Lycyaena. The extinct Lycyaena-Chasmaporthetes-Hyaenictis clade, which emerged as habitats opened up during the terminal Miocene, was unique in that its members exhibited post-cranial adaptations indicative of advanced cursoriality.4,30Hyaenictitherium namaquensis was a late-occurring member of the ictitherines, a clade of canid-like hyenas that were prominent during the middle Miocene and began dying out at the end of the Miocene.
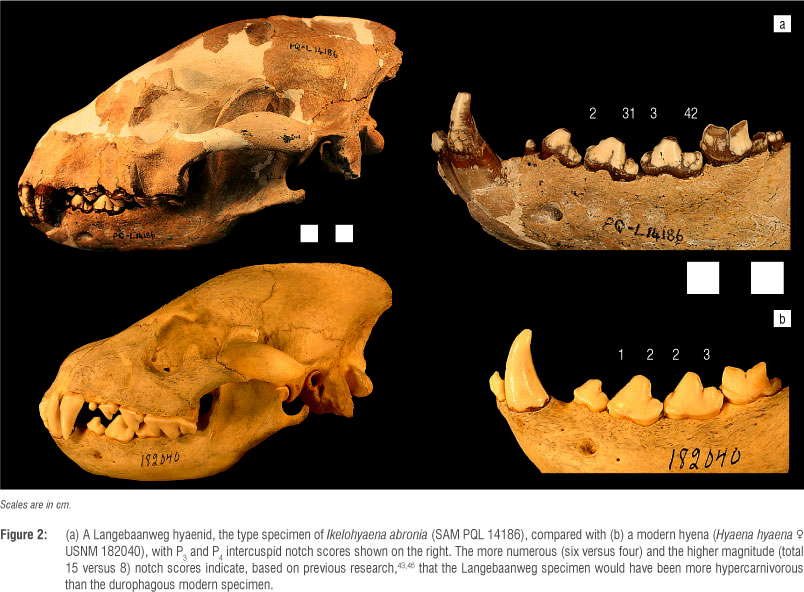
The goals of this study were (1) to compare the LBW hyaenids that are complete enough to include in analyses with the modern durophagous hyenas (Figure 2) and (2) to place them into the broader dietary context using a comparative sample of modern carnivorans and new dietary evaluation methods that focus on the premolar radii-of-curvature (ROC) and intercuspid notch scores (ICN). These methods have proven useful for arraying carnivorans along the carcass-processing continuum from durophagy (i.e. species that specialise in obdurate foods - in the case of hyenas, bone) to hypercarnivory (i.e. species that specialise in vertebrate flesh - a mechanically soft food). In combination, these approaches have yielded interesting conclusions about several guilds of extinct carnivorans43-45 and their application to the LBW hyaenids should help establish the position of these specimens along the hypercarnivory-durophagy gradient.
Materials and methods
Eleven hyaenid lower dental elements (sets of teeth) from LBW (Table 1) were selected based on minimal wear and maximal completeness. Minimal wear is necessary for the ROC analyses, and completeness (i.e. the presence of intact premolars and carnassial) is necessary for ICN analyses. Two LBW felids - Dinofelis cf. diastemata and Felinae indet. - were also included for comparison (Table 1).
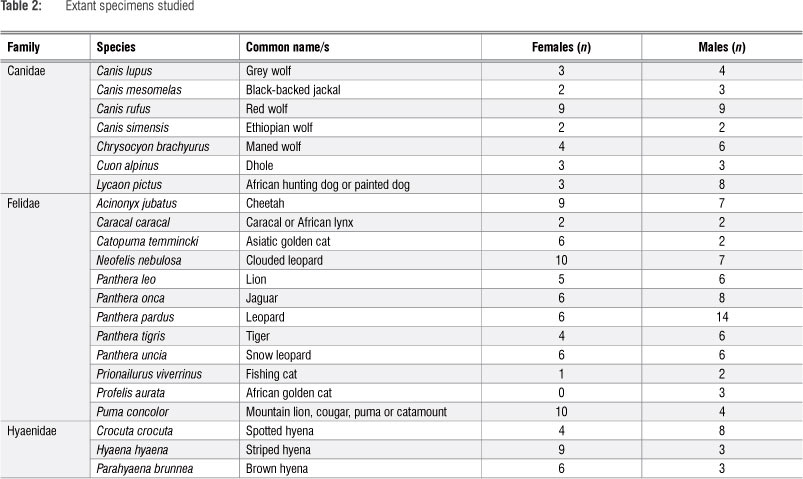
These fossil specimens were compared with a sample of modern specimens (Table 2) from the American Museum of Natural History and the National Museum of Natural History (Smithsonian). This subset of a sample has been described previously.43,46,47 It contains 7 canid species (n=61), 12 felid species (n=132) and all 3 durophagous hyaenid species (n=33). Previous studies using this sample included other modern (i.e. from other families) and fossil taxa;43,45-47 however, these species were omitted from the current study for visual simplicity of the results. Their inclusion (not shown) does not alter the conclusions of this study.
Because of the small sample size used throughout this study, all statistical significance was evaluated using a non-parametric Wilcoxon test with p<0.05 taken to be significant.
The first papers to describe the techniques employed in this paper43,46,47 used modified carnivoran dietary categories developed by Van Valkenburgh48,49 and employed by many subsequent researchers. In the modified approach,43,46,47 carnivoran species are divided into one of four categories. The first category is 'hypercarnivores' (some researchers call this the 'meat' category): species whose diet consists of more than 70% vertebrate flesh. 'Durophages', the second category, are species that masticate bone or other hard dietary elements (e.g. mollusk shell or bamboo). The other two dietary categories - 'meat/non-vertebrate' and 'non-vertebrate/meat' - incorporate fruits and/or insects as important dietary elements. For the purposes of this study, because we are examining taxa that likely included very few non-vertebrate elements in their diets, only the continuum spanning the hypercarnivory and durophagy diets were considered. For further discussion of the other carnivoran dietary niche space see Hartstone-Rose43.
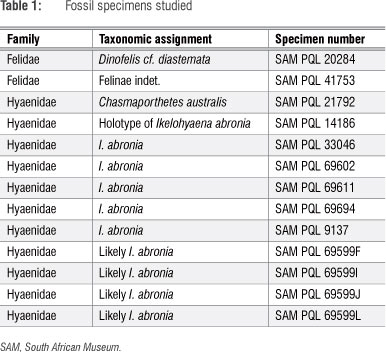
Cusp ROC were measured following methods previously described43,47 in an approach similar to that used by earlier researchers.50 In this approach, lower post-canine dental specimens are moulded using regular-body President Jet (Coltene Whaledent, Cuyahoga Falls, OH, USA), a quick-setting product used primarily in the dental industry, and then cast using Smooth-Cast (Smooth-On, Easton, PA, USA), a fast-setting, low-viscosity, two-part resin. The central cusp of each premolar and the two mesial cusps of the carnassial casts (Figure 3) are then sectioned in the coronal plane using a thin kerf scroll saw (Craftsman, Hoffman Estates, IL, USA). Each coronal section is then scanned (Figure 4) on a flatbed scanner (Lexmark, Lexington, KY, USA) and, using ImageJ (NIH, Bethesda, MD, USA), fitted with the smallest circle that describes the curvature of the tip. Unfortunately, teeth are not perfect simple geometric shapes and fitting a circle to a cusp point is clearly an oversimplification. Other techniques have been employed to fit more complex shapes (e.g. a parabola) to tooth cross sections, but these too are only slightly more accurate in describing the shape. Indeed, each additional term that could be added to the equation would make the line fit the curve of the tooth more accurately. However, the fitting of circles to premolar cross sections has been shown to be reproducible and indicative of diet43,46,47 and, because it is easier and faster than digitising the whole outline of the tooth and fitting lines with algorithms, more data can be incorporated into the sample more efficiently. Thus, as this method is sufficient for dietary analysis, and it is quick and easy, it is preferred. Per convention, the inverse of the radius of the fitted circle (i.e. 1/radius) is the ROC. Thus, sharper teeth have higher ROC values than blunter teeth.

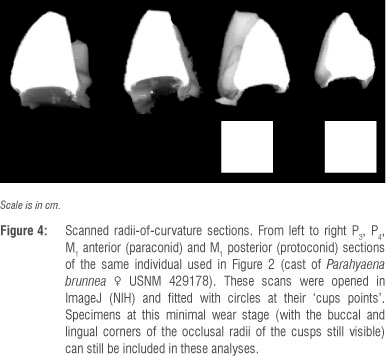
As discussed previously,43,47 only relatively unworn teeth are acceptable for this approach. Although moderate wear leaves enough of the occlusal point intact to conscribe a circle within, specimens that demonstrate wear that exceeds this level are excluded from the study (Figure 5). Not surprisingly, modern hyenas that demonstrate minimal enough wear to remain below this threshold are nearly exclusively cranial sub-adults.43 In other words, although all of the adult teeth are in occlusion (dental adult), the cranial sutures are still unfused and the crania have not reached the maximum adult dimensions. Hyenas that do exhibit cranial adulthood invariably have premolars that are worn beyond their occlusal point (beyond the level at which they can be used in this approach, see Figuren 5). Unfortunately, many of the relatively complete LBW dentaries are also worn beyond this threshold, substantially reducing the available sample for this study.

The measurements were not corrected for the size of the tooth or individual. As discussed previously,43 removing tooth size (e.g. by dividing the ROC by the buccolingual width of the tooth) would be a simple procedure (those data are available) but it actually confounds, rather than clarifies, the signal because the ROC does not vary substantially with body size (very low r-squared values)43 but rather by food mechanical properties. In other words, species with higher amounts of bone in their diets have blunter premolars than species with higher amounts of flesh in their diets independent of body size.43 So, because carnivores across the body size spectrum consume foods of similar mechanical properties, and those mechanical properties are size independent (i.e. a small piece of muscle tissue takes the same ROC to slice as a large piece of muscle tissue), predator body size (and therefore tooth size) is not a confounding factor of ROC. Although tooth size is highly correlated with body size,51 ROC is not43.
The ICNs were also measured following previously described techniques.43,46 In this approach, the notches between the cusps of the premolars are scored on a scale of 1-5 (Figure 2). The sharpness of the crests that lead into the notch is considered perpendicular to the slope of the crest itself. A score of 1 represents a barely visible notch (i.e. the notch distinguishing a barely present cusp). A score of 2 represents a notch between two dull or rounded crests. A score of 3 represents a notch between one dull crest and one sharp crest, <45°. (While this angle seems relatively arbitrary there is very little overlap in the range of crest premolar sharpness: carnivore premolar crests are either fairly rounded or obviously sharp. An intermediate morphology does not make biomechanical sense - a sharp crest needs to be sharp enough to slice or there is no functional reason to be sharp at all.) A score of 4 represents a notch between two sharp crests. A score of 5 represents a 'carnassiform notch' (defined below and in Figure 6).
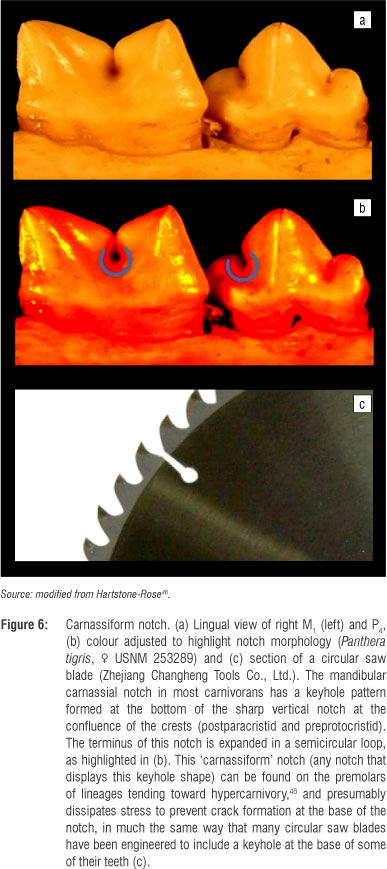
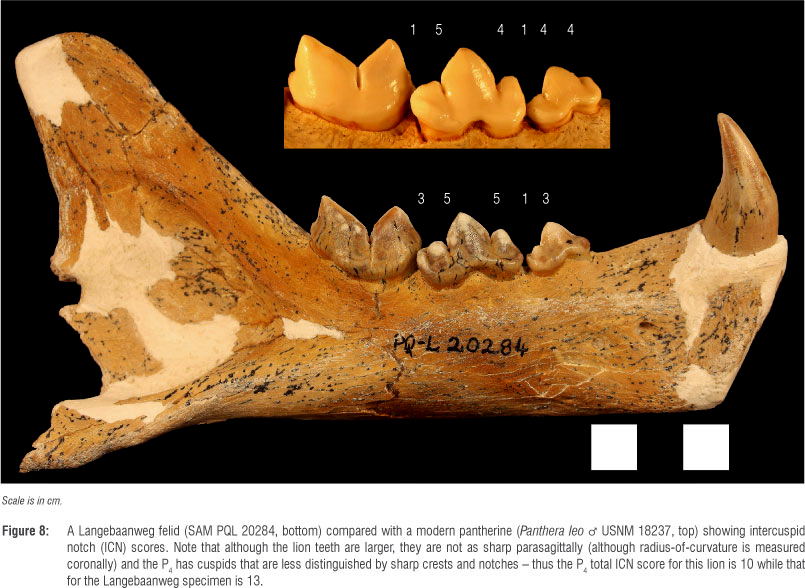
The carnassials (P4/M1) - the defining teeth of carnivorans - contain sharp blades, the lower paraconid and protoconid (although these crests are probably more correctly referred to as 'postparacristid' and 'preprotocristid'), that meet to form a diamond-shaped space that diminishes in size as the upper and lower teeth approach each other, culminating in occlusion (Figure 7). This configuration (much like the diamond-shaped space formed by the blades of wire cutters) concentrates soft foods (e.g. meat) toward the centre rather than squashing them out toward the sides as would occur if the blades did not have these opposing 'v' shapes.52 At its most extreme, this carnassial blade configuration is sharp and mesiodistally long, and the blades meet at their notch with a vertical slit. This slit ends in a rounded keyhole shape that presumably dissipates forces applied at the cusp tips in much the same way that industrial engineers build circular shapes into areas of high stress like the tooth notches in some circular saw blades. This keyhole pattern is found on the carnassials of almost all carnivorans (only the most herbivorous taxa have reduced this feature so much that the carnassial lacks the keyhole pattern and the blade-like configuration in general) and is therefore referred to as 'carnassiform'.43,46 The most hypercarnivorous taxa have these carnassiform notches in their premolars as well - notches that result from the confluence of two sharp crests that terminate in a keyhole, stress dissipating, morphology. The carnassiform notch is most likely found on the 'main notch' (the notch just distal to the paraconid or between the paraconid and the first distal accessory cuspid) on the P4, although more hypercarnivorous species have carnassiform notches in more than one location on their premolar row.43,46
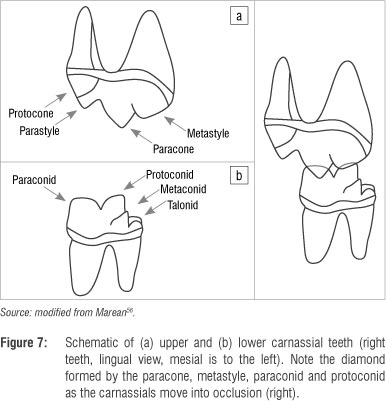
In the most hypercarnivorous carnivorans (e.g. felids), the lower carnassial (M1) consists entirely of these two crested cusps, while more omnivorous carnivorans (e.g. canids) maintain a talonid, grinding, portion of the tooth that occludes with the upper first molar. The three modern bone-eating hyaenids have a highly reduced carnassial talonid, especially C. crocuta which lacks upper molars. Hyaena hyaena and P. brunnea retain the M1 and have more substantial talonids, although their talonids are still highly reduced relative to the cynoid form.
Results
The LBW hyaenid premolars and anterior carnassial cusp (paraconid) are slightly sharper than those of the modern hyaenids, although not statistically so (Table 3). As previously observed, this signal is strongest at the more anterior premolars, with the P3 ROC showing the highest correlation to carcass processing abilities.43,47 The apparently (although not statistically) slightly sharper premolars of the LBW hyaenids would seem to indicate a slight tendency of these hyaenids toward greater hypercarnivory than the durophagous modern hyaenids. The LBW felids fall within the ROC ranges of the modern felids (Table 3) indicating no difference in their carcass processing abilities based on these metrics.
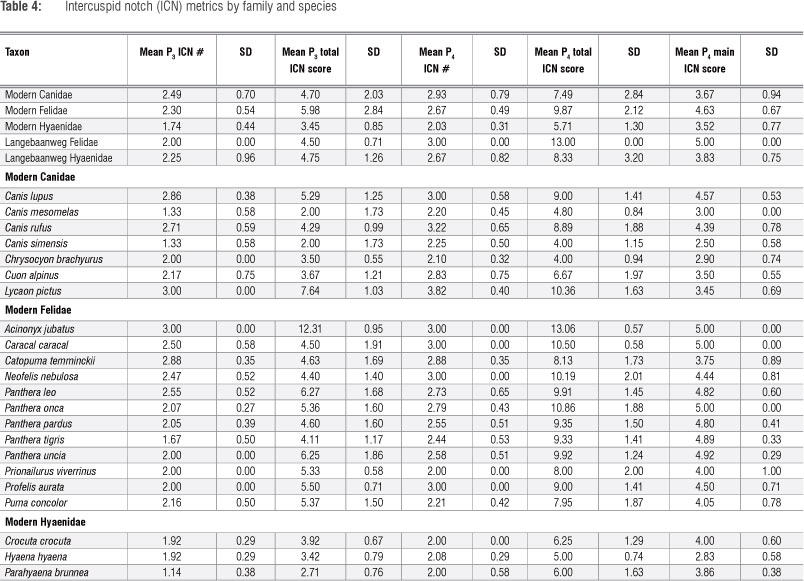
The LBW hyaenids have higher ICN scores than modern hyaenids (Table 4). This finding is statistically significant for the P4 total ICN score. Even though hyenas use their P3 predominantly for bone cracking,13 it is the P4 total ICN score that best separates species according to their carcass processing abilities.43,46 By this measure, the LBW hyaenids were significantly more hypercarnivorous than their durophagous confamilials.
For this same variable (P4 total ICN score), the LBW felids also score higher than modern felids, again suggesting greater hypercarnivory (or even higher reliance on flesh relative to more obdurate dietary elements) than their modern confamilials. However, in other ICN metrics (e.g. analyses of the P3) the LBW felids are close to the mean scores of modern felids. This can be seen when the specimens are compared with a dentary of a modern felid (Figure 8).
Previous research has suggested that the P3 ROC and P4 total ICN scores best separate carnivorans according to their carcass processing abilities with more hypercarnivorous modern taxa having sharper and more cusped premolars than the more durophagous species.43,46 In examining these two metrics together, the LBW hyaenid sample almost exceeds the range of the modern bone-cracking hyaenids and overlaps with some canids and the most durophagous felids (Figure 9). In terms of their ROC, particularly that of the lower third premolar - the tooth that best discriminates taxa according to diet47 - the LBW hyaenid teeth fall toward the sharp end of the modern hyena range and would all be, broadly, within the sharpest half of the modern group. However, as seen metrically, the LBW hyaenids are graphically (and statistically) more clearly differentiated from the modern hyenas in terms of their ICN scores; several of the LBW hyenas fall completely outside of the modern hyena range of total fourth premolar ICN scores, and those that do overlap with the range of modern species, overlap only with the most highly notched individuals (2 of the 12 modern C. crocuta) in the sample (Figure 9). Again, the total P4 ICN score has previously been shown43,46 to best sort species according to diet, with the more hypercarnivorous taxa characterised by notably higher ICN scores (more intercuspid notches) and the more durophagous taxa characterised by lower ICN scores (fewer intercuspid notches).
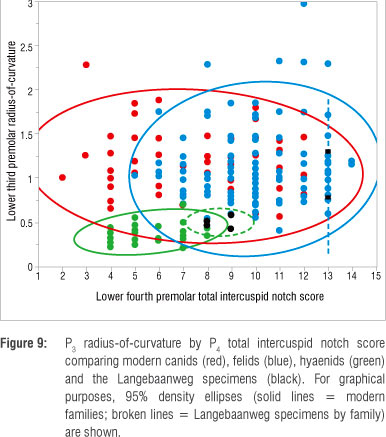
The two LBW felid specimens included in the sample fall within the modern felid range, both in terms of their ROC and ICNs (Figure 9). The total P4 ICN score of the two LBW felids is toward the hypercarnivorous end of the modern felid sample falling within the central range of the modern cheetah (Acinonyx jubatus) sample - more notched (hypercarnivorous) than any of the members of the genus Panthera. Only 1 of the 11 specimens of the most hypercarnivorous canid, the African hunting dog (Lycaon pictus), has an equally high ICN score, and thus the LBW felids were probably more hypercarnivorous than any modern canid species, on average, comparable only to the most hypercarnivorous modern felid, A. jubatus.
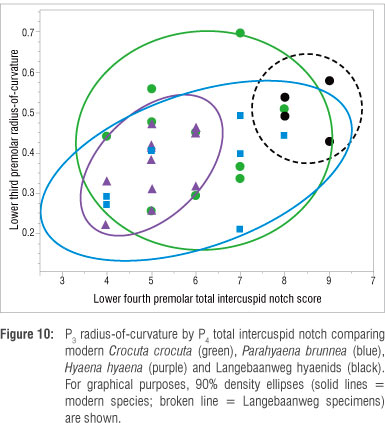
When examining these same metrics (P3 ROC and P4 total ICN) within the hyaenids (Figure 10), it is apparent that the LBW hyaenids appear more hypercarnivorous (toward the upper-right region of the graph) than any of the three modern species of durophagous hyaenids. Although the LBW specimens statistically overlap the ranges of C. crocuta and P. brunnea, they do not overlap that of H. hyaena at all; the fossil specimens all have significantly higher ICN scores than any of the H. hyaena specimens. This finding would seem to indicate that H. hyaena is more durophagous than the LBW hyaenids - and indeed more durophagous in general than its three confamilials although no dietary literature seems to allude to this difference.53
Discussion
The LBW hyaenids are fascinating from both an evolutionary and a functional perspective: how do they fit within their family phylogenetically, and how did they function ecologically? Much has been written about them from a phylogenetic perspective4,18,38 but, until recently, little attention has been paid to their diet through detailed metric analyses.39,40,44
With that said, the earliest descriptions of the LBW hyaenids indicated that these animals filled ecological niches similar to those occupied by modern canids. The present analyses partially confirm this belief -the LBW hyenas that were included in this study are certainly not as durophagous as the modern hyenas are. This observation is particularly interesting considering that I. abronia, the taxon that makes up the majority of this sample, was supposed to be the most durophagous of the LBW hyena species.30 However, in terms of their ROC and ICN, the LBW hyenas fall on the periphery of both the modern felid and modern canid morphospace, and indeed overlap slightly with the space occupied by two of the three durophagous modern hyenas (Figure 9). The LBW hyaenids have slightly sharper teeth (higher ROC) than most modern hyenas, although generally duller teeth (lower ROC) than almost all modern canids and felids. Their teeth have more notches (higher ICN scores) than almost all modern hyenas, and they have lower ICN scores than most modern felids. In this respect they are indeed more dog-like (with very similar mean ICN scores to those of the Canidae), showing higher notch scores (indicating greater hypercarnivory46) than some modern canids (like Canis mesomelas, C. simensis and Chrysocyon brachyurus) and lower notch scores than other modern canid species (like Canis lupus, C. rufus and Lycaon pictus). By this metric, the LBW hyenas appear to fall in the middle of the hypercarnivory-durophagy range of modern Canidae. However, in terms of their cusp sharpness they are much more hyena-like than canid-like. Perhaps this is a signal that ICN is more correlated with diet while ROC maintains a tighter phylogenetic signal - a finding that confirms patterns which emerged in previous analyses.43
Only two LBW felids were included in this study; nonetheless, interesting results were obtained. Although the two LBW specimens clearly fall- within the modern felid range for both ROC and ICN, they have higher ICN scores than most modern felids, suggesting, by this metric, that they were in fact more hypercarnivorous than many modern felid species are. Clearly, more work needs to be done to support this finding.
Obviously a broader LBW sample of both families would add to the robustness of this analysis as would the inclusion of some of the LBW specimens from other families (e.g. Agriotherium and Plesiogulo as well as the fox-like cf. Eucyon sp.). Likewise, a larger morphological sample would allow the use of more powerful multivariate tools like principal components analysis. However, with few exceptions, the LBW sample is fairly fragmentary, and most multivariate approaches (such as principal components analysis) require more complete specimens than those included in this study. If only the more complete specimens were analysed, the already small sample size would be reduced beyond most statistical approaches. With that said, this preliminary sample does seem to confirm a relatively late shift toward hyaenid durophagy while the LBW felids already exhibited the hypercarnivory that is the hallmark of their modern confamilials.
It is intriguing that both the LBW hyaenids and felids have similar ROC scores to their modern confamilials, yet both groups have relatively high ICN scores. A more complete analysis of the entire LBW carnivore guild would elucidate niche space, but these two lineages appear to be slightly less durophagous than their modern relatives. If the durophagous niche was occupied at LBW, which of the locality's species was most likely to have done so? While one cannot discount non-mammalian carnivores such as vultures, of which a single undescribed species has been discovered at LBW, two large mammalian carnivores, namely the giant wolverine Plesiogulo monspessulanus and African bear Agriotherium africanum, remain the strongest candidates. The latter species certainly had the capacity to consume large, mechanically demanding bone,54 although its ability to do so on a regular basis is still debated.55 Neither of these impressive species (or any close relatives) live in modern day Africa. Perhaps, if this bear and wolverine were the Mio-Pliocene durophages of Langebaanweg, then, as they receded toward extinction, the LBW hyaenids were replaced by more durophagous taxa to fill the newly freed niche space. Perhaps this move toward greater durophagy in the Hyaenidae actually caused these extinctions. Regardless, the adaptations of the modern bone-cracking hyenas are among the most impressive in all mammalian history, and understanding the more gracile anatomy of their Mio-Pliocene relatives is clearly informative of how these species operate within the carnivore guild.
In this paper, we have confirmed that the two methods, ROC and ICN analyses, contribute valuable insights into the diet of these species -insights that both corroborate previous findings (e.g. a more generalist diet for the LBW hyenas) but also suggest more complex interpretations (e.g. the meaning of the potentially conflicting ROC and ICN results). Future expansions of this study, by inclusion of more of the LBW taxa, will allow a broader guild-level analysis of the carnivorans from this site. Modifications of the methods to allow inclusion of disassociated premolars will allow a broadening of the hyena sample to include all of the LBW species. Furthermore, the application of these methods to other fossil samples will allow more complete discussion of the changing diets across geographic and temporal guilds. All of these further directions will allow greater insight into the dietary evaluation of fascinating fossil carnivoran species.
Acknowledgements
We thank Graham Avery for access to the specimens and informative conversations about them; Brittany Bovard, Kristen MacNeill, Elizabeth Simpler, Nathan Yasika and Gina Zappia for help with data collection; and four anonymous reviewers for helpful comments.
Authors' contributions
A.H-R. formulated the hypotheses and methods, collected the data, performed some of the analyses, and wrote the first draft and subsequent revisions of the manuscript. D.D.S. helped with the analyses, wrote major sections of the manuscript and helped address concerns posed by reviewers.
References
1. Koehelr CE, Richardson PRK. Proteles cristatus. Mamm Species. 1990;363:1-6. http://dx.doi.org/10.2307/3504197 [ Links ]
2. Cooper RL, Skinner JD. Importance of termites in the diet of the aardwolf Proteles cristatus in South Africa. S Afr J Zool. 1979;14:5-8.
3. Kruuk H, Sands WA. The aardwolf (Proteles cristatus Sparrman 1783) as a predator of termites. East Afr Wildlife J. 1972:10:211-227. http://dx.doi.org/10.1111/j.1365-2028.1972.tb00728.x [ Links ]
4. Werdelin L, Solounias N. The Hyaenidae: Taxonomy, systematics, and evolution. Fossils Strata. 1991;30:1-104. [ Links ]
5. Werdelin L, Peigné S. Carnivora. In: Werdelin L, Sanders WJ, editors. Cenozoic mammals of Africa. Berkeley: University of California Press; 2010. p. 609-663. http://dx.doi.org/10.1525/california/9780520257214.003.0032 [ Links ]
6. Ewer RF. The carnivores; Ithaca, NY: Cornell University Press; 1973. [ Links ]
7. Koepfli KP Jenks S, Eizirik E, Zahirpour T, Van Valkenburgh B, Wayne RK.
Molecular systematics of the Hyaenidae: Relationships of a relictual lineage resolved by a molecular supermatrix. Mol Phylogenet Evol. 2006;38:603-620. http://dx.doi.org/10.1016/j.ympev.2005.10.017
8. Tseng ZJ, Wang XM. Cranial functional morphology of fossil dogs and adaptation for durophagy in Borophagus and Epicyon (Carnivora, Mammalia). J Morphol. 2010;271:1386-1398. [ Links ]
9. Tseng ZJJ, Wang XM. Do convergent ecomorphs evolve through convergent morphological pathways? Cranial shape evolution in fossil hyaenids and borophagine canids (Carnivora, Mammalia). Paleobiology. 2011;37:470-489. http://dx.doi.org/10.1002/jmor.10881 [ Links ]
10. Kruuk H. The spotted hyena. A study of predation and social behavior. Chicago: University of Chicago Press; 1972. p. 336. [ Links ]
11. Ferretti MP. Evolution of bone-cracking adaptations in hyaenids (Mammalia, Carnivora). Swiss J Geosci. 2007;100:41-52. http://dx.doi.org/10.1007/s00015-007-1212-6 [ Links ]
12. Tseng ZJ, Antón M, Salesa MJ. The evolution of the bone-cracking model in carnivorans: Cranial functional morphology of the Plio-Pleistocene cursorial hyaenid Chasmaporthetes lunensis (Mammalia: Carnivora). Paleobiology. 2011;37:140-156. http://dx.doi.org/10.1666/09045.1 [ Links ]
13. Van Valkenburgh B. Feeding behavior in free-ranging, large African carnivores. J Mammal. 1996;77:240-254. http://dx.doi.org/10.2307/1382725 [ Links ]
14. Hendey QB. The late Cenozoic carnivora of the south-western Cape Province. Ann S Afr Mus. 1974;63:1-369. [ Links ]
15. Hendey QB. Late Tertiary Hyaenidae from Langebaanweg, South Africa, and their relevance to the phylogeny of the family. Ann S Afr Mus. 1978;76:265-297. [ Links ]
16. Werdelin L. Carnivoran ecomorphology: A phylogenetic perspective. In: Gittleman J, editor. Carnivore behavior, ecology, and evolution. Ithaca, NY: Cornell University Press; 1996. p. 582-624. [ Links ]
17. Werdelin L, Solounias N. The evolutionary history of hyenas in Europe and western Asia during the Miocene. In: Bernor RL, Fahlbusch V Rietschel S, editors. Later Neogene European biotic evolution and stratigraphic correlation. New York: Columbia University Press; 1996. p. 290-306. [ Links ]
18. Werdelin L, Turner A. The fossil and living Hyaenidae of Africa: Present status. In: Stewart KM, Seymour KL, editors. Palaeoecology and palaeoenvironments of Late Cenozoic mammals: Tributes to the career of C.S. (Rufus) Churcher. Toronto: University of Toronto Press; 1996. p. 635-659. [ Links ]
19. Hendey QB. A review of the geology and palaeontology of the Plio/Pleistocene deposits at Langebaanweg, Cape Province. Ann S Afr Mus. 1970;56:75-117. [ Links ]
20. Hendey QB. Further observations on the age of the mammalian fauna from Langebaanweg, Cape Province. Paleoecology. 1972;6:172-175. [ Links ]
21. Hendey QB. Fossil occurrences at Langebaanweg, Cape Province. Nature. 1973;244:13-14. http://dx.doi.org/10.1038/244013a0 [ Links ]
22. Hendey QB. New fossil carnivores from the Swartkrans australopithecine site (Mammalia, Carnivora). Ann Transv Mus. 1974;29:27-48. [ Links ]
23. Hendey QB. Fossil peccary from the Pliocene of South Africa. Sci Total Environ. 1976;192:787-789. [ Links ]
24. Harris JM. Pliocene Giraffoidea (Mammalia: Artiodactyla) from the Cape Province. Ann S Afr Mus. 1976;69:325-353. [ Links ]
25. Gentry A. Fossil Bovidae (Mammalia) from Langebaanweg, South Africa. Ann S Afr Mus. 1980;79:213-337. [ Links ]
26. Grine FE, Hendey QB. Earliest primate remains from South Africa. S Afr J Sci. 1981;77:374-376. [ Links ]
27. Rich P. Preliminary report on the fossil avian remains from late Tertiary sediments at Langebaanweg (Cape Province), South Africa. S Afr J Sci. 1980;76:166-170. [ Links ]
28. Olson S. Early Pliocene ibises (Aves, Plataleidae) from south western Cape Province, South Africa. Ann S Afr Mus. 1985;97:57-69. [ Links ]
29. Franz-Odendaal T, Kaiser T, Bernor R. Systematics and dietary evaluation of a fossil equid from South Africa. S Afr J Sci. 2003;99:453-459. [ Links ]
30. Werdelin L. The position of Langebaanweg in the evolution of Carnivora in Africa. Afr Nat Hist. 2006;2:201-202. [ Links ]
31. Sanders W. Taxonomic review of fossil Proboscidea (Mammalia) from Langebaanweg, South Africa. Trans Roy Soc S Afr. 2007;62:1-16. http://dx.doi.org/10.1080/00359190709519192 [ Links ]
32. Govender R, Avery G, Chinsamy A. Pathologies in the Early Pliocene phocid seals from Langebaanweg, South Africa. S Afr J Sci. 2011;107(1/2):72-77.
33. Govender R, Chinsamy A, Ackermann RR. Anatomical and landmark morphometric analysis of fossil phocid seal remains from Langebaanweg, West Coast of South Africa. Transactions of the Royal Society of South Africa 2012, 67, 135-149. http://dx.doi.org/10.1080/0035919X.2012.724471 [ Links ]
34. Hendey QB. Geological succession at Langebaanweg, Cape Province, and global events of the late Tertiary. S Afr J Sci. 1981;77:33-38.
35. Roberts D, Matthews T, Herries A, Boulter C, Scott LD, C Ntembi P et al. Regional and global palaeoenvironmental and sea level context of the Late Cenozoic Langebaanweg (LBW) palaeontological site: West coast of South Africa. Earth Sci Rev. 2011;106:191-214. http://dx.doi.org/10.1016/j.earscirev.2011.02.002 [ Links ]
36. Roberts D. Lithostratigraphy of the Varswater Formation. South African Committee for Stratigraphy-Lithostratigraphic Series. 2006;9:27-31. [ Links ]
37. Hendey QB. Langebaanweg: A record of past life. Cape Town: South African Museum; 1989. p. 71. [ Links ]
38. Werdelin L, Turner A, Solounias N. Studies of fossil hyaenids - the genera Hyaenictis gaudry and Chasmaporthetes hay, with a reconsideration of the Hyaenidae of Langebaanweg, South Africa. Zool J Linn Soc. 1994;111:197-217. http://dx.doi.org/10.1111/j.1096-3642.1994.tb01483.x [ Links ]
39. Stynder DD. Tooth crown form as an indicator of niche partitioning among Late Miocene/Early Pliocene hyenas from 'E' Quarry, Langebaanweg, South Africa. Paleogeogr Paleoclimatol Paleoecol. 2009;283:148-159. http://dx.doi.org/10.1016/j.palaeo.2009.09.017 [ Links ]
40. Stynder DD, Ungar PS, Scott JR, Schubert BW. A dental microwear texture analysis of the Mio-Pliocene hyaenids from Langebaanweg, South Africa. Acta Palaeontol Pol. 2012;57(3):485-496. http://dx.doi.org/10.4202/app.2011.0053 [ Links ]
41. Turner A, Antón M, Werdelin L. Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe. Geobios. 2008;41:677-687. http://dx.doi.org/10.1016/j.geobios.2008.01.001 [ Links ]
42. Tseng ZJ, Stynder D. Mosaic functionality in a transitional ecomorphology: Skull biomechanics in stem Hyaeninae compared to modern South African carnivorans. Biol J Linn Soc. 2011;102:540-559. http://dx.doi.org/10.1111/j.1095-8312.2010.01602.x [ Links ]
43. Hartstone-Rose A. Evaluating the hominin scavenging niche through analysis of the carcass-processing abilities of the carnivore guild [dissertation]. Durham, NC: Duke University; 2008. [ Links ]
44. Hartstone-Rose A, Simpler EM, Heckler AL. The diet of the Mio-Pliocene carnivores of Langebaanweg, South Africa. J Vertebr Paleontol. 2011;31:122A.
45. Simpler EM, Perry JMG, Hartstone-Rose A. Determining diet from the cross-sectional shape and intercuspid notches of the teeth of the carnivores of Rancho La Brea. J Vertebr Paleontol. 2011;31(Suppl. 3):195A. http://dx.doi.org/10.1111/j.1469-7998.2011.00821.x [ Links ]
46. Hartstone-Rose A. Reconstructing the diets of extinct South African carnivorans from premolar 'Intercuspid Notch' morphology. J Zool (Lond). 2011;285:1-10. [ Links ]
47. Hartstone-Rose A, Wahl S. Using radii-of-curvature for the reconstruction of extinct South African carnivoran masticatory behavior. C R Palevol. 2008;7:629-643. http://dx.doi.org/10.1016/j.crpv.2008.09.015 [ Links ]
48. Van Valkenburgh B. Trophic diversity in past and present guilds of large predatory mammals. Paleobiology. 1988;14:155-173.
49. Van Valkenburgh B. Carnivore dental adaptations and diet: A study of trophic diversity within guilds. In: Gittleman JL, editor. Carnivore behavior, ecology, and evolution. Ithaca, NY: Cornell University Press; 1989. p. 410-436. [ Links ]
50. Popowics TE, Fortelius M. On the cutting edge: Tooth blade sharpness in herbivorous and faunivorous mammals. Ann Zool Fenn. 1997;34:73-88.
51. Van Valkenburgh B. Skeletal and dental predictors of body mass in carnivores. In: Damuth J, MacFadden BJ, editors. Body size in mammalian paleobiology: Estimation and biological implications. Cambridge: Cambridge University Press; 1990. p. 181-205. [ Links ]
52. Lucas PW. Dental functional morphology; how teeth work. Cambridge: Cambridge University Press; 2004. p. 355. http://dx.doi.org/10.1017/CBO9780511735011 [ Links ]
53. Rieger I. Hyaena hyaena. Mamm Species. 1981;150:1-5. [ Links ]
54. Oldfield C, McHenry C, Clausen P Chamoli U, Parr W, Stynder D, et al. Finite element analysis of ursid cranial mechanics and the prediction of feeding behaviour in the extinct giant Agriotherium africanum. J Zool (Lond). 2012;286:93-171. http://dx.doi.org/10.1111/j.1469-7998.2011.00862.x [ Links ]
55. Stynder D, Kupczik K. Tooth root morphology in the early Pliocene African bear Agriotherium africanum (Mammalia, Carnívora, Ursidae) and its implications for feeding ecology. J Mamm Evol. In press 2012. http://dx.doi.org/10.1007/s10914-012-9218-x [ Links ]
56. Marean CW. Sabertooth cats and their relevance for early hominid diet and evolution. J Hum Evol. 1989;18:559-582. http://dx.doi.org/10.1016/0047-2484(89)90018-3 [ Links ]
 Correspondence:
Correspondence:
Adam Hartstone-Rose
University of South Carolina School of Medicine
CBA Building 1, Room C-36, 6439
Garners Ferry Road, Columbia, SC 29209, USA
Email: Adam.Hartstone-Rose@uscmed.sc.edu
Received: 16 Oct. 2012
Revised: 10 Jan. 2013
Accepted: 18 Jan. 2013


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}