










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.109 no.5-6 Pretoria ene. 2013
REVIEW ARTICLE
Pleistocene Homo and the updated Stone Age sequence of South Africa
Gerrit DusseldorpI; Marlize LombardII; Sarah WurzIII
ICentre for Anthropological Research, University of Johannesburg, Johannesburg, South Africa
IIDepartment of Anthropology and Development Studies, University of Johannesburg, Johannesburg, South Africa
IIIInstitute for Human Evolution, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
We provide a brief overview of how the rich South African Pleistocene Homo fossil record correlates with the recently revised Stone Age sequence. The overview and correlation of the data is intended to highlight gaps in the record and/or our understanding thereof, and to stimulate interdisciplinary research and debate on the Homo fossil and archaeological records spanning the Pleistocene. As an updated resource we present a complete inventory of known Pleistocene fossil material assigned to the genus Homo, and, where possible, its association with archaeological material. We demonstrate that (1) anatomical changes are not necessarily paralleled by changes in the archaeological sequence currently based on a range of technocomplexes, (2) the early Homo record of South Africa probably differs from that of East Africa, (3) mid-Pleistocene Homo might be associated with the Earlier to Middle Stone Age transitional phase and (4) the fossil record associated with the Middle Stone Age has wide anatomical variation. Also, hiatuses in the fossil record, such as that associated with the appearance of early Khoe-San-like populations, do not show concurrent hiatuses in the archaeological record. Thus, for a broader understanding of the demographic history of South Africa during the Pleistocene, both sources of information should be considered in tandem.
Keywords: Stone Age sequence; Homo; Pleistocene; South Africa; archaeology
Introduction
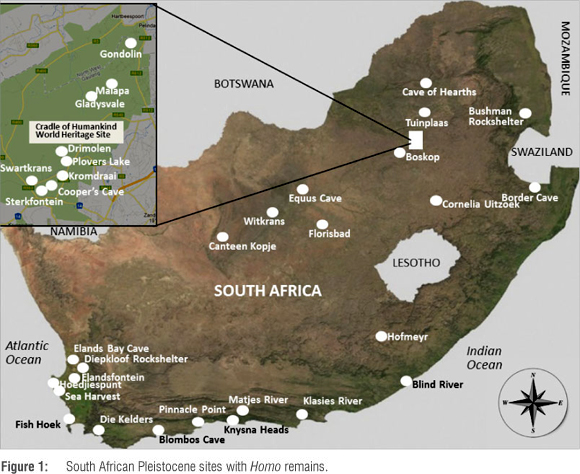
South Africa has a rich hominin fossil record and a seemingly uninterrupted archaeological sequence spanning at least the last 2 million years. A broad, up-to-date and integrated overview that is accessible to researchers from a range of disciplines is, however, lacking. Recently, the Stone Age archaeological sequence was updated, broadly aligned with the marine isotope stage (MIS) record, and adjustments to nomenclature proposed.1 Here we present an initial correlation of the revised Stone Age sequence with the known Pleistocene Homo fossil record of South Africa. This correlation is accompanied by an inventory of published South African Pleistocene Homo fossil material (over 200 fossils from 31 sites; see Table 1 and Supplementary Table 1 online). The aim of our short overview is to provide an updated database summarising human anatomical and behavioural evolutionary trends during the Pleistocene in South Africa. We also hope to stimulate interdisciplinary research and debate. By considering different data sets (i.e. the Stone Age sequence and the complete Pleistocene Homo fossil record), resolution, problems, and gaps in the records are underscored. This approach complements studies focusing separately on archaeology or palaeoanthropology or on either 'early Homo' or the evolution of 'modern Homo sapiens', which constitute the usual approaches in both disciplines.
Although our focus is on the genus Homo, it must be noted that before ~1 Ma Homo probably occurred sympatrically with robust australopithecines.2 The dating of the Malapa site suggests that earliest Homo existed concurrently with the newly described Australopithecus sediba.3,4 The presence of Paranthropus in many deposits said to contain early Homo makes ascribing isolated teeth and postcrania to Homo tentative at best.3,5 Furthermore, the co-existence of multiple hominin species complicates attributing cultural remains to any of these species. The production of the earliest Oldowan stone tools is usually ascribed to Homo,6,7 but circumstantial evidence suggests that species not assigned to Homo may also have produced stone tools. For example, metacarpal anatomy indicates that Paranthropus robustus from South Africa could knap.5,8 Moreover, the infill with Oldowan stone tools (Member 5 East) at Sterkfontein only contains fossils assigned to Paranthropus,9 whereas the breccia, associated with StW 53 that is generally ascribed to Homo,10 does not contain any stone tools.9 Correlation between the early archaeological and fossil Homo records thus remains imprecise, and by presenting sites yielding Homo fossils together with the MIS and archaeological records, we do not imply direct association between the data sets. Relationships between hominin/human remains, archaeological and/or environmental contexts throughout the sequence should be independently investigated.
Early Homo
Several sites have been reported to yield early Homo fossils (Table 1, Figure 1), but the interpretation of the record is complex (see Supplementary Table 1 online for sites, fossils and accession numbers). Firstly, the definition of the genus Homo is not clear-cut, because not all scholars agree that Homo habilis should be included.11 Secondly, the taxonomic identity of key fossils is debated. SK 847 from Swartkrans, for example, has been classified as A. africanus, P. robustus, H. habilis, H. erectus, H. leakeyi, Homo sp. nov. and H. gautengensis.10,12,13 Thirdly, taxonomic identification is further complicated by the fact that most remains comprise isolated teeth, or fragmented cranial or mandibular pieces. Some fossils from Sterkfontein and Swartkrans have been assigned to H. habilis (e.g. StW 53, SK 27, SK 847), some from Swartkrans Members 1 and 2 to H. erectus (e.g. SK 45, SK 847, SK 15), and fossils from Sterkfontein Member 5 West to H. ergaster (e.g. StW 80). Most of the fossils attributed to H. erectus or H. ergaster by some have been referred to as early Homo (e.g. H. habilis or H. gautengensis) by others (see Supplementary Table 1 online). It is possible that, similar to the East African record, several species of Homo co-existed.14,15
Recent cladistic analyses suggest that some important early Homo fossils (StW 53, SK 15, SK 27, SK 45, SK 847) do not fit within either the H. erectus/H. ergaster or H. habilis hypodigms.10,16 Curnoe13 proposes a new species, H. gautengensis, for this material, while others maintain that two early Homo species are represented at Sterkfontein and Swartkrans.17 StW 53 is the holotype for the proposed South African representative of early Homo, H. gautengensis13. StW 53 has previously been assigned to H. habilis together with SK 847,13,18 but is classified as A. africanus by some.9
The assignation of H. habilis to the genus Homo is contested.11 Because the dental characteristics of the South African material assigned to early Homo show closer affinities to East African H. habilis than to H. ergaster/H. erectus,16 a similar argument might apply to H. gautengensis. The strongest affinities are found within the South African material itself.16 Hence it may be inappropriate to equate the South African fossil sequence to an evolutionary scheme based on the East African record. The recent find of a tooth dated to ~1 Ma at Cornelia Uitzoek,19 associated with Acheulean artefacts and sharing most affinities with early South African Homo, reinforces the suggestion that the hominin succession of South Africa may have been unlike that of East Africa.
A cautious interpretation of the available evidence suggests that forms exhibiting derived anatomical characteristics that are also present in Homo, such as A. sediba and StW 151,3,20 are represented in South Africa from at least 2 Ma onwards. Thus far, no generally agreed upon fossils belonging to H. habilis and H. erectus/H. ergaster are known from South Africa. Given the anatomical variation in the reported fossil materials, a scenario in which multiple species occupied the region during the early Pleistocene is a strong possibility. The fragmented nature of most of- the material has, however, resulted in a lack of consensus on which fossils should be grouped together as subcategories of early Homo. The complicated taphonomic history of most deposits and the uncertain dating, with varying proposed age estimates for key deposits,2,9 could conceal diachronic trends within the fossil record. The taxonomic designation of the representation of the genus Homo before ~1 Ma in South Africa thus remains challenging, and most of the fossils from this group (see Supplementary Table 1 online) are only provisionally classified as Homo.
Mid-Pleistocene Homo
A second group of fossils (see Supplementary Table 1 online), dated to between ~1 Ma and ~200 ka, is mainly represented by the Elandsfontein skullcap and the Florisbad skull (Table 1). Such specimens have been ascribed to H. helmei, H. heidelbergensis, H. rhodesiensis and archaic H. sapiens.21,22 A distinction is sometimes made between an earlier group that includes the Elandsfontein skull, and a group exhibiting more modern morphology represented by the Florisbad skull (see Supplementary Table 1 online).22 Yet, it has been proposed that the African fossils from ~700 ka onwards represent a gradually evolving lineage - H. sapiens sensu lato.23 Another suggestion, however, is that the transition from mid-Pleistocene Homo to H. sapiens was a punctuated speciation event, during which the form of the cranium was re-organised.24 Both hypotheses await further exploration and corroboration.
The South African fossil inventory from this important phase in human development is limited, and dating resolution for most fossils remains poor.25 Determining the archaeological context of the fossils is problematic, because only the Cave of Hearths specimen was found in- direct association with archaeological materials. However, temporally, this group of fossils is generally associated with Acheulean, Earlier Stone Age to Middle Stone Age transitional and/or early Middle Stone Age assemblages (Table 1),1,9,26 encompassing important technological and behavioural change. Recent research suggests that transitional Earlier Stone Age-Middle Stone Age Fauresmith assemblages, possibly dating to ~500 ka at Kathu Pan 1, show systematic blade production and the possible hafting of stone artefacts interpreted as having tipped hunting weapons.27,28 If the dating and functional interpretation of the artefacts are accepted as accurate (although results have yet to be replicated), their reporting underscores previous suggestions that composite hunting weapons were not exclusive to more recent H. sapiens and H. neanderthalensis.29
Middle Stone Age anatomically modern Homo sapiens
Few South African hominin fossils can be placed between ~200 ka and 110 ka - the phase during which the transformation from mid-Pleistocene Homo to modern H. sapiens probably took place in the region. One of these fossils is the partial femur with modern morphology that was excavated at Blind River and dated to MIS 5e or ~118 ka.30 Another is the single archaic H. sapiens tooth from the Lincoln Cave, which was obtained from reworked deposits.31 The Sea Harvest fossils may also date from this phase, but their most likely age is considered to be somewhat younger, associated with MIS 5b starting at ~95 ka.32 Notwithstanding the sparse fossil record, more than 10 dated archaeological assemblages, including those from Pinnacle Point Cave 13B and Klasies River on the south coast, Kathu Pan in the Northern Cape, Florisbad in the Free State and Border Cave in KwaZulu-Natal, indicate human presence across the landscape at the time.1
Dating to between -110 ka and 40 ka, a third group of fossils, exemplified by the Klasies River sample (Figure 2), is classified as morphologically modern H. sapiens (Table 1, Figure 1, Supplementary Table 1 online). The fully modern BC 1 and BC 2 fossils from Border Cave are from secondary context and could date to -171-152 ka, but- also to -91-71 ka or even be as young as the Holocene; hence they cannot be considered here.33,34 The fossil sample from this period is relatively small and most specimens are isolated teeth. The population is recognised as modern H. sapiens, but some specimens, such as the proximal ulna from the Deacon excavations at Klasies River, exhibit archaic morphology.35 At several sites the buccolingual and mesiodistal diameters of the teeth exceed the average size of contemporary modern humans, and some specimens even fall outside the 2σ range from the modern average.36 Interestingly, the Klasies River sample also contains specimens with dimensions smaller than the modern average.37-39
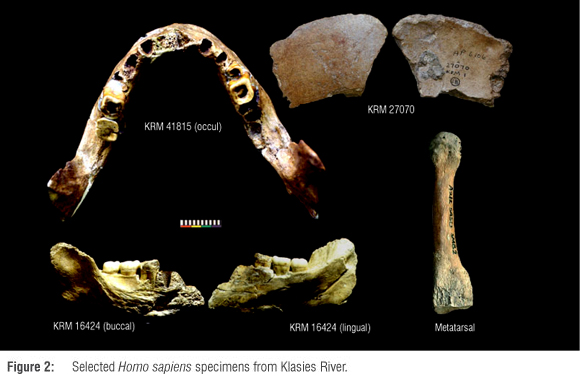
The fossils in this group are associated with a range of Middle Stone Age technocomplexes (Table 1, Supplementary Table 1 online),1 the majority of which date to MIS 5 and MIS 4. Archaeological material linked to this phase has been interpreted to indicate increased levels of technological, behavioural and cognitive complexity,29,40-44 perhaps related to shifts in demography.45-47 On the whole, the fossil record from this period suggests that South Africa was occupied by populations showing a wide range of anatomical variation. The populations were morphologically mostly modern, but the process of gracilisation, leading to the form and dimensions of contemporary populations, was not yet completed.
Final Middle Stone Age/early Later Stone Age Homo sapiens
Two fossils have been dated to the early stages of the final Middle Stone Age/early Later Stone Age period: the Hofmeyr skull (~36 ka) and the mandible from Bushman Rock Shelter (~30 ka).48,49 Both fossils show fully modern morphology within the metric range of modern humans, but their dimensions fall outside the range exhibited by modern Khoe-San. Metrically, the Hofmeyr skull resembles Upper Palaeolithic Europeans most closely.49 Unfortunately the Hofmeyr skull was found in secondary context, and the archaeological association of the Bushman Rock Shelter remains is unclear.48,49 The transition from late MIS 3 H. sapiens to late MIS 2/early MIS 1 Khoe-San-like populations is obscured by a seeming hiatus in the fossil record. The fossil record, however, largely depends on accidental discovery, and is hampered by sampling, dating and- preservation biases. A previously suggested population decline during MIS 3, partly based on the perceived fossil hiatus, is not supported by a recent synthesis of the MIS 3 archaeology of southern Africa.50
Resolution for final Middle Stone Age and early Later Stone Age technocomplexes, spanning MIS 3 and the initial stages of MIS 2, is poor and assemblages overlap in time.1 However, it is important to understand the division between the Middle Stone Age and the Later Stone Age as a heuristic strategy, rather than an absolute boundary.1,51 Material from sites with sequences that span the two phases often demonstrate gradual technological transformation of Middle Stone Age into Later Stone Age industries,52-54 which might not have occurred simultaneously at all sites or in all areas in the region.55 The most recent confirmation of this interpretation stems from the re-analysis of Border Cave lithic material.56
The Canteen Kopje skull, with an uncertain age,57 and the Tuinplaas skeleton, tentatively dated to ~20~11 ka, but probably only slightly older than the minimum age estimate of ~11 ka,58 are anatomically fully modern. Slightly younger skeletons from Elands Bay Cave and Matjes River, possibly dating to the terminal Pleistocene/early Holocene at ~11 ~10 ka, are anatomically similar to modern Khoe-San,59 and the deliberate burial of individuals now seems common. Finally, a number of fossils that were conventionally thought to date to the Pleistocene, and have been grouped as 'Boskopoid', have been redated. Of these, the Fish Hoek skeleton proved to date to the mid-Holocene,60 and the Cape Flats skull to ~150 BP61
Conclusion
The South African Homo fossil record, as presented in the inventory in Supplementary Table 1 online, combined with the revised Stone Age sequence1 arguably provides the most comprehensive current data set documenting human anatomical and technological developments for the region. This review and the accompanying inventory demonstrate that a diverse and large scholarship developed around the South African palaeoanthropological and archaeological records. By presenting this synthesis and inventory in a user-friendly format, we aim to provide an expedient analytical tool for multidisciplinary research and discussion around the evolution of our genus in the region. The inclusive approach has the potential to clarify patterns and/or highlight problem areas that are often obscured when particular issues and controversies are debated, because focused discourse often precludes presenting empirical information not immediately relevant to the particular question at hand. The lacunae in the fossil record do not imply that the South African landscape was void of hominin occupation during such times, as the archaeological record attests to their presence during most phases. We therefore emphasise that it is important to base demographic interpretations on both the fossil and archaeological data sets. Hiatuses and/or discrepancies in either data set, however, draw attention to potential future excavation, analytical and research focus areas and the need to revisit previously excavated archaeological assemblages that may yield more human fossil material.
Acknowledgements
The work of Marlize Lombard and Sarah Wurz is supported by the African Origins Platform of the National Research Foundation (NRF). Gerrit Dusseldorp is supported by the NRF micro-TrACKS programme directed by Marlize Lombard, and previously received support from the NRF SARCHI programme directed by Christopher Henshilwood. Any opinions expressed in this paper are those of the authors and not the NRF.
Authors' contributions
All authors contributed equally to drafting and editing the manuscript. M.L. suggested the concept and G.L.D. collated the online supplementary material, which served as a foundation for the contribution.
References
1. Lombard M, Wadley L, Deacon J, Wurz S, Parsons I, Mohapi M, et al. South African and Lesotho Stone Age sequence updated. S Afr Archaeol Bull. 2012;67:123-144. [ Links ]
2. Herries AIR, Curnoe D, Adams JW. A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa. Quat Int. 2009;202:14-28. http://dx.doi.org/10.1016/j.quaint.2008.05.017 [ Links ]
3. Pickering R, Dirks PHGM, Jinnah Z, De Ruiter DJ, Churchill SE, Herries AIR, et al. Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo. Science. 2011;333:1421-1423. http://dx.doi.org/10.1126/ science.1203697 [ Links ]
4. Pickering TR, Heaton JL, Clarke RJ, Sutton MB, Brain CK, Kuman K. New hominid fossils from Member 1 of the Swartkrans formation, South Africa. J Hum Evol. 2012;62:618-628. http://dx.doi.org/10.1016/j.jhevol.2012.02.003 [ Links ]
5. Tocheri MW, Orr CM, Jacofsky MC, Marzke MW. The evolutionary history of the hominin hand since the last common ancestor of Pan and Homo. J Anat. 2008;212:544-562. http://dx.doi.org/10.1111/j.1469-7580.2008.00865.x [ Links ]
6. Tobias PV. Australopithecus, Homo habilis, tool-using and tool-making. S Afr Archaeol Bull. 1965;20:167-192.
7. Clark JD. The prehistory of Africa. London: Thames & Hudson; 1970. [ Links ]
8. Susman RL. Fossil evidence for early hominid tool use. Science. 1994;265:1570-1573. http://dx.doi.org/10.1126/science.8079169
9. Kuman K, Clarke RJ. Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5. J Hum Evol. 2000;38:827-847. http://dx.doi.org/10.1006/jhev.1999.0392
10. Smith HF, Grine FE. Cladistic analysis of early Homo crania from Swartkrans and Sterkfontein, South Africa. J Hum Evol. 2008;54:684-704. http://dx.doi.org/10.1016/j.jhevol.2007.10.012
11. Wood B, Collard M. The human genus. Science. 1999;284:65-71. http://dx.doi.org/10.1126/science.284.5411.65
12. Olson TR. Hominid phylogenetics and the existence of Homo in Member I of the Swartkrans formation, South Africa. J Hum Evol. 1978;7:159-178. http://dx.doi.org/10.1016/S0047-2484(78)80008-6
13. Curnoe D. A review of early Homo in southern Africa focusing on cranial, mandibular and dental remains, with the description of a new species (Homo gautengensis sp. nov.). HOMO - J Comp Hum Biol. 2010;61:151-177. http:// dx.doi.org/10.1016/j.jchb.2010.04.002
14. Spoor F, Leakey MG, Gathogo PN, Brown FH, Anton SC, McDougall I, et al. Implications of new early Homo fossils from Ileret, east of Lake Turkana, Kenya. Nature. 2007;448:688-691. http://dx.doi.org/10.1038/nature05986 [ Links ]
15. Leakey MG, Spoor F, Dean MC, Feibel CS, Antón SC, Kiarie C, et al. New fossils from Koobi Fora in northern Kenya confirm taxonomic diversity in early Homo. Nature. 2012;488:201-204. http://dx.doi.org/10.1038/nature11322 [ Links ]
16. Grine FE, Smith HF, Heesey CP Smith EJ. Phenetic affinities of Plio-Pleistocene Homo fossils from South Africa: Molar cusp proportions. In: Grine FE, Fleagle JG, Leakey RE, editors. The first humans: Origin and early evolution of the genus Homo. Dordrecht, Heidelberg, New York, London: Springer; 2009. p. 49-62. http://dx.doi.org/10.1007/978-1-4020-9980-9_6 [ Links ]
17. Ungar PS, Grine FE, Teaford MF, El Zaatari S. Dental microwear and diets of African early Homo. J Hum Evol. 2006;50:78-95. http://dx.doi.org/10.1016/j.jhevol.2005.08.007
18. Curnoe D, Tobias PV. Description, new reconstruction, comparative anatomy, and classification of the Sterkfontein Stw 53 cranium, with discussions about the taxonomy of other southern African early Homo remains. J Hum Evol. 2006;50:36-77. http://dx.doi.org/10.1016/j.jhevol.2005.07.008
19. Brink JS, Herries AIR, Moggi-Cecchi J, Gowlett JAJ, Bousman CB, Hancox JP et al. First hominine remains from a ~1.0 million year old bone bed at Cornelia-Uitzoek, Free State Province, South Africa. J Hum Evol. 2012;63:527-535. http://dx.doi.org/10.1016/j.jhevol.2012.06.004 [ Links ]
20. Moggi-Cecchi J, Grine FE, Tobias PV. Early hominid dental remains from Members 4 and 5 of the Sterkfontein Formation (1966-1996 excavations): Catalogue, individual associations, morphological descriptions and initial metrical analysis. J Hum Evol. 2006;50:239-328. http://dx.doi.org/10.1016/j.jhevol.2005.08.012
21. Brãuer G. Middle Pleistocene diversity in Africa and the origin of modern humans. In: Hublin J-J, McPherron Sp editors. Modern origins: A North African perspective. Dordrecht, Heidelberg, New York, London: Springer; 2012. p. 221-240. http://dx.doi.org/10.1007/978-94-007-2929-2_15 [ Links ]
22. Rightmire GP Middle and later Pleistocene hominins in Africa and Southwest Asia. Proc Natl Acad Sci USA. 2009;106:16046-16050. http://dx.doi.org/10.1073/pnas.0903930106
23. Brãuer G. The origin of modern anatomy: By speciation or intraspecific evolution. Evol Anthropol. 2008;17:22-37. http://dx.doi.org/10.1002/evan.20157
24. Lieberman DE, Bar-Yosef O. Apples and oranges: Morphological versus behavioural transitions in the Pleistocene. In: Lieberman DE, Smith RJ, Kelley L, editors. Interpreting the past: Essays on human, primate and mammal evolution. Boston: Brill Academic Publishers; 2005. p. 275-296. [ Links ]
25. Millard AR. A critique of the chronometric evidence for hominid fossils: I. Africa and the Near East 500-50 ka. J Hum Evol. 2008;54:848-874. http://dx.doi.org/10.1016/j.jhevol.2007.11.002
26. Beaumont PB, Vogel JC. On a timescale for the past million years of human history in central South Africa. S Afr J Sci. 2006;102:217-228.
27. Wilkins J, Chazan M. Blade production ~500 thousand years ago at Kathu Pan 1, South Africa: Support for a multiple origins hypothesis for early Middle Pleistocene blade technologies. J Arch Sci. 2012;39:1883-1900. http://dx.doi.org/10.1016/j.jas.2012.01.031
28. Wilkins J, Schoville BJ, Brown KS, Chazan M. Evidence for early hafted hunting technology. Science. 2012;338:942-946. http://dx.doi.org/10.1126/science.1227608
29. Lombard M, Haidle MN. Thinking a bow-and-arrow set: Cognitive implications of Middle Stone Age bow and stone-tipped arrow technology. Camb Archaeol J. 2012;22:237-264. http://dx.doi.org/10.1017/S095977431200025X
30. Wang Q, Tobias P Roberts D, Jacobs Z. A re-examination of a human femur found at the Blind River Site, East London, South Africa: Its age, morphology, and breakage pattern. Anthropol Rev. 2008;71:43-61. http://dx/doi. org/10.2478/v10044-008-0009-1
31. Reynolds SC, Clarke RJ, Kuman KA. The view from the Lincoln Cave: Mid- to late Pleistocene fossil deposits from Sterkfontein hominid site, South Africa. J Hum Evol. 2007;53:260-271. http://dx.doi.org/10.1016/j. jhevol.2007.02.004
32. Grine FE, Klein RG. Late Pleistocene human remains from the Sea Harvest site, Saldanha Bay, South Africa. S Afr J Sci. 1993;98:145-152.
33. Schwartz JH, Tattersall I. The human fossil record Volume two: Craniodental morphology of genus Homo (Africa and Asia). Hoboken: Wiley-Liss; 2003. [ Links ]
34. Millard AR. Bayesian analysis of ESR dates, with application to Border Cave. Quat Geochronol. 2006;1:159-166. http://dx.doi.org/10.1016/j.quageo.2006.03.002
35. Churchill SE, Pearson OM, Grine FE, Trinkaus E, Holliday TW. Morphological affinities of the proximal ulna from Klasies River main site: Archaic or modern? J Hum Evol. 1996;31:213-237. http://dx.doi.org/10.1006/jhev.1996.0058 [ Links ]
36. Grine FE. Middle Stone Age human fossils from Die Kelders Cave 1, Western Cape Province, South Africa. J Hum Evol. 2000;38:129-145. http://dx.doi.org/10.1006/jhev.1999.0353
37. Singer R, Wymer J, editors. The Middle Stone Age at Klasies River Mouth in South Africa. Chicago: Chicago University Press; 1982. [ Links ]
38. Rightmire GP, Deacon HJ. New human teeth from Middle Stone Age deposits at Klasies River, South Africa. J Hum Evol. 2001;41:535-544. http://dx.doi.org/10.1006/jhev.2001.0500
39. Grine FE. Observations on Middle Stone Age human teeth from Klasies River Main Site, South Africa. J Hum Evol. 2012;63:750-758. http://dx.doi.org/10.1016/j.jhevol.2012.08.005 [ Links ]
40. Brown KS, Marean CW, Herries AIR, Jacobs Z, Tribolo C, Braun D, et al. Fire as an engineering tool of early modern humans. Science. 2009;325:859-862. http://dx.doi.org/10.1126/science.1175028 [ Links ]
41. d'Errico F, Henshilwood C, Vanhaeren M, Van Niekerk K. Nassariuskraussianus shell beads from Blombos Cave: Evidence for symbolic behaviour in the Middle Stone Age. J Hum Evol. 2005;48:3-24. http://dx.doi.org/10.1016/j. jhevol.2004.09.002 [ Links ]
42. Henshilwood CS, d'Errico F, Watts I. Engraved ochres from the Middle Stone Age levels at Blombos Cave, South Africa. J Hum Evol. 2009;57:27-47. http://dx.doi.org/10.1016/j.jhevol.2009.01.005
43. Wadley L, Sievers C, Bamford M, Goldberg P Berna F, Miller C. Middle Stone Age bedding construction and settlement patterns at Sibudu, South Africa. Science. 2011;334:1388-1391. http://dx.doi.org/10.1126/science.1213317 [ Links ]
44. Wadley L, Hodgskiss T, Grant M. Implications for complex cognition from the hafting of tools with compound adhesives in the Middle Stone Age, South Africa. Proc Natl Acad Sci USA. 2009;106:9590-9594. http://dx.doi. org/10.1073/pnas.0900957106 [ Links ]
45. Powell A, Shennan S, Thomas MG. Late Pleistocene demography and the appearance of modern human behavior. Science. 2009;324:1298-1301. http://dx.doi.org/10.1126/science.1170165 [ Links ]
46. Wurz S. Modern behaviour at Klasies River. Goodwin Ser S Afr Archaeol Soc. 2008;10:150-156.
47. d'Errico F, Stringer CB. Evolution, revolution or saltation scenario for the emergence of modern cultures? Phil Trans R Soc B. 2011;366:1060-1069. http://dx.doi.org/10.1098/rstb.2010.0340
48. Protsch R, De Villiers H. Bushman Rock Shelter, Origstad, Eastern Transvaal, South Africa. J Hum Evol. 1974;3:387-396. http://dx.doi.org/10.1016/0047-2484(74)90201-2
49. Grine FE, Bailey RM, Harvati K, Nathan RP Morris AG, Henderson GM, et al. Late Pleistocene human skull from Hofmeyr, South Africa, and modern human origins. Science. 2007;315:226-229. http://dx.doi.org/10.1126/ science.1136294 [ Links ]
50. Mitchell PJ. Developing the archaeology of Marine Isotope Stage 3. Goodwin Ser S Afr Archaeol Soc. 2008;10:52-65.
51. Mitchell PJ. The archaeology of southern Africa. Cambridge: Cambridge University Press; 2002. [ Links ]
52. Kaplan J. The Umhlatuzana Rock Shelter sequence: 100 000 years of Stone Age history. Natal Mus J Human. 1990;2:1-94.
53. Clark AMB. Late Pleistocene technology at Rose Cottage Cave: A search for modern behavior in an MSA context. Afr Archaeol Rev. 1999;16:93-119. http://dx.doi.org/10.1023/A:1021955013009
54. McCall GS, Thomas JT. Re-examining the South African Middle-to-Later Stone Age transition: Multivariate analysis of the Umhlatuzana and Rose Cottage Cave stone tool assemblages. Azania. 2009;44:311-330. http:// dx.doi.org/10.1080/00672700903337519
55. Wadley L. The Pleistocene Later Stone Age south of the Limpopo River. J World Prehist. 1993;7:243-296. http://dx.doi.org/10.1007/bf00974721
56. Villa P Soriano S, Tsanova T, Degano I, Higham TFG, d'Errico F, et al. Border Cave and the beginning of the Later Stone Age in South Africa. Proc Natl Acad Sci USA. 2012;109:13208-13213. http://dx.doi.org/10.1073/ pnas.1202629109 [ Links ]
57. Smith P Nshimirimana R, De Beer F, Morris D, Jacobson L, Chazan M, et al. Canteen Kopje: A new look at an old skull. S Afr J Sci. 2012;108(1/2), Art #738, 9 pages. http://dx.doi.org/10.4102/sajs.v108i1/2.738 [ Links ]
58. Pike AWG, Eggins S, Grün R, Thackeray F. U-series dating of TP1, an almost complete human skeleton from Tuinplaas (Springbok Flats), South Africa. S Afr J Sci. 2004;100:381-383. [ Links ]
59. Stynder DD, Ackermann RR, Sealy JC. Early to mid-Holocene South African Later Stone Age human crania exhibit a distinctly Khoesan morphological pattern. S Afr J Sci. 2007;103:349-352.
60. Stynder DD, Brock F, Sealy JC, Wurz S, Morris AG, Volman TP. A mid-Holocene AMS 14C date for the presumed upper Pleistocene human skeleton from Peers Cave, South Africa. J Hum Evol. 2009;56:431-134. https://dx.doi.org/10.1016/j.jhevol.2008.11.004 [ Links ]
61. Singer R. Mythical African 'Australoids' and triangular bricks: The Cape Flats skull in retrospect. S Afr Archaeol Bull. 1993;48:105-112.
62. Morris AG. A master catalogue: Holocene human skeletons from South Africa. Johannesburg: Witwatersrand University Press; 1992. [ Links ]
 Correspondence:
Correspondence:
Gerrit Dusseldorp
Centre for Anthropological Research, University of Johannesburg
PO Box 524, Auckland Park 2006, South Africa
Email:g.dusseldorp@gmail.com
Received: 16 Oct. 2012
Revised: 10 Jan. 2013
Accepted: 18 Jan. 2013


{kind=link}