










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.108 no.7-8 Pretoria ene. 2012
RESEARCH ARTICLE
Immobilisation of yeast cells on carbon nanotubes
Tirivaviri A. MamvuraI; Sunny E. lyukeI; Vusumuzi SibandaI; Clarence S. YahII
ISchool of Chemical and Metallurgical Engineering, University of the Witwatersrand, Johannesburg, South Africa
IIToxicology & Biochemistry Section, National Institute for Occupational Health, National Health Laboratory Service, Johannesburg, South Africa
ABSTRACT
Carbon nanotubes are increasingly finding application in a wide range of industries. The focus of this study was to investigate the immobilisation of yeast cells onto carbon nanotubes, using a flocculation method, for possible use in fermentation processes. Carbon nanotubes, which are long thin cylinders of carbon, were used as artificial agents to induce flocculation of yeast cells. The immobilisation experiments on carbon nanotubes were conducted under different process conditions and compared with control experiments done on free cells. The resultant immobilised cells or flocs were recovered and freeze dried before analysis was performed. The flocculated cells were characterised by scanning electron microscopy to confirm that flocculation had occurred. Conditions that gave optimum flocculation on carbon nanotubes were found to be: a pH between 5.0 and 5.8, a temperature between 25 °C and 30 °C, an agitation speed of about 110 rpm, and a concentration of carbon nanotubes (in powder form) of between 44 mg/mL and 54 mg/mL. The addition of calcium ions and glucose decreased the rate of flocculation and delayed the onset of flocculation. Our study has demonstrated that carbon nanotubes have great potential to improve the flocculation capacity of brewer's yeast.
Introduction
There are numerous biotechnological processes that make use of immobilisation techniques to manipulate cells. These techniques can be divided into four major categories based on the physical mechanism at play in bringing about the immobilisation of the cells. These categories are (1) attachment or adsorption on solid carrier surfaces, (2) entrapment within a porous matrix, (3) natural aggregation by flocculation, and (4) artificially induced cross-linking by agents and/ or cell containment behind barriers.1 Amongst the available yeast cell immobilisation techniques, the flocculation of microorganisms is very attractive, because of its simplicity and low cost. Flocculation does not involve any complex and costly mechanical devices or any supporting material in its operation, which represents a significant advantage over other immobilisation techniques.2
Flocculation is defined as the phenomenon of loose aggregation of free yeast cells as a result of random collisions by Brownian motion. The cell aggregates then rapidly settle from the medium in which they are suspended because of their increased mass.3 A typical example is the yeast cells flocculation that occurs at the end of a fermentation process in the stationary phase, where the flocculated cells either sink to the bottom of the fermenter or rise to the surface attached to carbon dioxide bubbles. Flocculation can also be induced by an artificial agent to increase the efficiency of the process and/or lower the overall process cost.
Carbon nanotubes (CNTs), which are widely used for catalysis, either as catalysts themselves, or as a catalyst support,4 were tested in this work as possible flocculation surfaces to immobilise brewer's yeast. Immobilisation often mimics what occurs in nature when cells grow on surfaces or within natural structures. Many microorganisms, including yeast cells, have the ability to adhere to and form a biofilm on different kinds of surfaces in nature.5 Multiwalled CNTs are relatively affordable materials, making them an attractive option as artificial flocculation agents.6
In fermentation, flocculation commonly occurs when the sources of fermentable sugars are exhausted. It has been suggested that, under such starvation conditions, the ability to form flocs may represent a stress response. Thus flocs provide a sheltered environment where the chance of survival of the population is enhanced. Disaggregation of flocs occurs if the cells are again exposed to a source of fermentable sugars. In this case, the re-adsorption of a single cell mode affords an unimpeded opportunity to utilise the supply of sugar.7
Live yeast cells have an intracellular negative charge because of the presence of a transmembrane potential and they can be attracted to cations or positively charged substances. However, dead cells, which have leaky membranes and cannot build a membrane potential, are not negatively charged and cannot be attracted to positively charged substances.8 In other words, during flocculation using positively charged CNTs, dead yeast cells cannot be attracted to CNTs and therefore cannot be flocculated by them.
This flocculation process can be applied in the ethanol industry to remove the suspended yeast cells after the fermentation process to reduce the turnaround time for the process.
Materials and methods
Microorganisms
Yeast cells were grown in a 500-mL flask held on a rotary shaking incubator working at 110 ± 2 revolutions per minute for 24 h. A rubber cork was used to cover the mouth of the flask throughout the experiment and temperature was maintained at 30 ± 0.5 °C.9,10
Growth curves were investigated for stationary phase and showed that the yeast cell concentrations were on average 65.75 x 106 colony forming units (CFU)/mL after growth and 62.63 x 106 CFU/mL on average before flocculation. The cells were freeze dried for later analysis and viability tests were performed.
Medium
Yeast extract (Merck, Johannesburg, South Africa) with additives was used as the medium for cell nutrition. The medium was sterilised at 121 °C and 1.1 bars in an autoclave for 20 min. The pH of the medium was 6.90 and was adjusted by addition of an acid to within fermentation pH range of between 4.00 and 6.00 before immobilisation experiments were resumed. Tests were also carried out at lower pH values (less than 4.00).
Carbon nanotubes
Carbon nanotubes were produced as reported in earlier work11 and their structure was confirmed by transmission electron microscopy (JEOL JEM 100S, Akishima, Tokyo, Japan).
Cell immobilisation
Yeast cell immobilisation was performed with CNTs whilst a control experiment was set up using yeast cells in the absence of CNTs. A colony of yeast was added to an Erlenmeyer flask containing 100 mL of sterilised medium and incubated in a shaker at 110 rpm and 30 °C for 24 h. After 24 h the yeast cells were used for immobilisation studies. Yeast broth (30 mL of 7.04 x 106 CFU/mL) was added to 250 mL of medium and incubated in a shaking incubator under the same conditions.12
Peinado et al.13 conducted immobilisation studies at 28 °C and 150 rpm for 7 days and were successful in producing yeast biocapsules. Sakurai et al.14 used conditions of 30 °C and 160 rpm during immobilisation studies of yeast cells on porous cellulose carriers. Öztop et al.15 immobilised Saccharomyces cerevisiae onto acrylamide-sodium acrylate hydrogels at 30 °C for 72 h.
Analytical methods
Yeast flocculation was analysed using two methods: a qualitative process to determine the quality of the flocs produced and a quantitative process to measure the flocculation weight. The first method was used to estimate the flocculation quality visually by looking at the sides and at the base of the Erlenmeyer flask. Flocculation was thus expressed qualitatively as: (-) no flocculation, (+) yeast slightly flocculent (poor), (++) yeast flocculent or (+++) yeast very flocculent.16,17,18 The second method - a quantitative method - involved the use of a centrifuge to concentrate the flocs, which were then recovered and dried at 40 °C for 24 h to determine their dry weight. The flocculated cells were recovered by a freeze dryer (VirTis, SP Industries, Warminster, PA, USA) and immobilisation was confirmed by scanning electron microscopy (JEOL JSM 840A, Akishima, Tokyo, Japan). The floc weight was then plotted against the variables under investigation to determine the effect of the respective variable on flocculation.
Results and discussion
Immobilisation of yeast cells
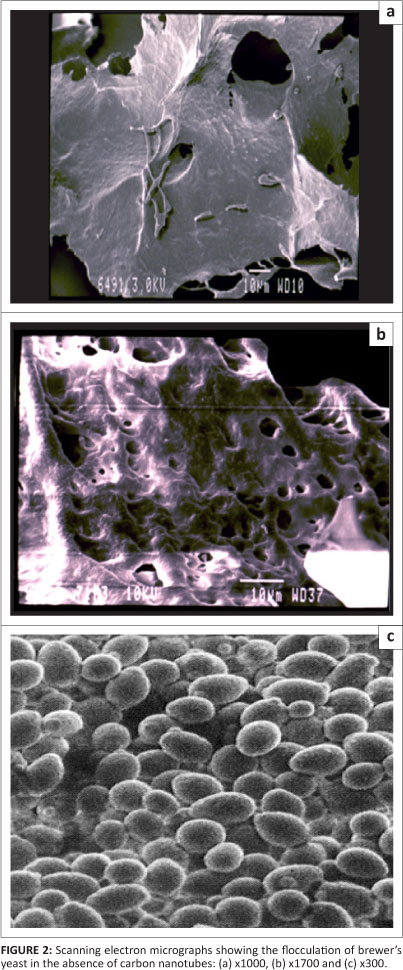
Figure 1 shows scanning electron micrographs of the immobilised cells or the flocculated yeast cells on the CNTs. The micrographs demonstrate that the immobilised yeast cells aligned themselves along the length of the CNTs. By comparing Figures 1c and 1d (which have the same magnification), it can be seen that the diameters of the CNTs increase when they are immobilised with yeast cells, that is, yeast cells are aligned along the length of the CNTs. This phenomenon was observed in all experiments where immobilisation was performed with CNTs. In contrast, free cells showed more planar flocculation structures, as seen in the micrograph in Figure 2. The free cells aligned on the surface forming a planar structure.
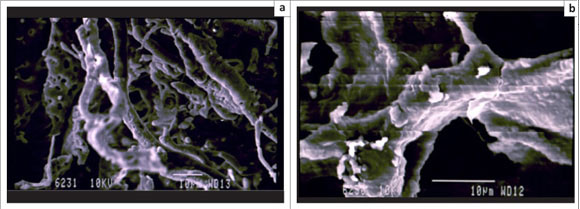
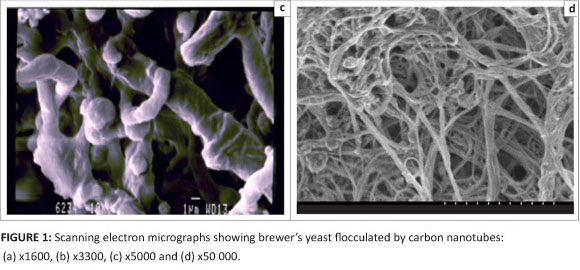
A number of factors which could affect the flocculation of yeast cells were investigated, for flocculation in the presence of CNTs and for free cell flocculation. These factors were: pH, temperature, concentration of CNTs, concentration of calcium ions, glucose and agitation speed.
Effect of pH
The study was conducted in the pH range of 1.30-6.50, which falls within the range used in fermentation processes (the target processes for these immobilised cells). The effects of pH are summarised in Figure 3.
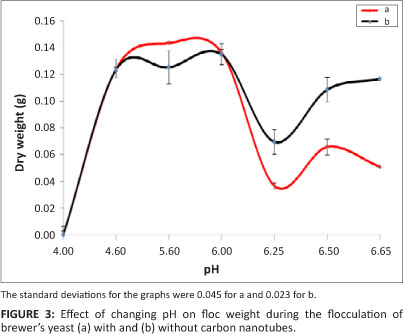
From Figure 3, it can be observed that the optimum pH for yeast flocculation was between 4.60 and 6.00 for both immobilisation on CNTs and flocculation of free cells. In this pH range, the floc weight obtained using CNTs was considerably higher than that obtained using free cells. This optimum pH range is close to the brewing pH range of 3.80-5.60 as reported in the literature.18,19,20,21,22,23 Above a pH of 6.00, the dry floc weight in both cases decreased rapidly until a pH of 6.25, before it increased slightly.
Flocculation became weak beyond a pH of 6.10; this finding could be as a result of the fact that yeast cells generally reverse their charge above a pH of 5.80. In aqueous suspensions, at the pH values of worts and beers (3.80-5.60), brewer's yeast cells migrate to the anode in electrophoresis experiments, thus behaving as negatively charged colloids. At more acidic pH values, reversal of the charge may take place,18 which may help to explain the decrease in flocculation weight at a pH below 4.60 and above 5.80, as was observed in this study. The fact that CNTs were able to flocculate yeast cells within the mentioned pH range showed that the CNTs are positively charged and repel the cells when the cells have a positive charge because of a change in pH. According to the literature, yeast cells should flocculate anywhere between a pH of 2.00 and 8.00, depending on the strain, with optimum flocculation occurring at a pH between 3.00 and 6.00. Our study showed similar results, with flocculation observed at a pH between 5.00 and 5.80. At low pH values (2.90-4.00), the cells might have been denatured, resulting in poor flocculation.19,24
Effect of immobilisation temperature
There is an apparent contradiction in the literature about the effect of temperature on flocculation: some authors report de-flocculation with increasing temperature while others report an increase in flocculation with increasing temperature. This discrepancy may be attributed to differences in the response of ale and lager strains.25 Jin et al.20,21 found that flocculation of a lager yeast strain varied from 24.1% at 5 °C to 66.8% at 25 °C, that is, that flocculation increased as temperature increased. However, there is little or no effect of temperature on the flocculation of brewer's yeast within the physiological temperature range of 15 °C-32 °C.19
Most brewing strains have an optimum temperature for growth that is between 30 °C and 34 °C,7,19,26 with viability losses during flocculation at 30 °C for 3 days considered negligible.27 Yeast autolysis normally occurs at elevated temperatures of between 40 °C and 60 °C.27,28,29,30 In this study, the effect of temperature on flocculation was investigated at 25 °C and 30 °C and the results are presented in Table 1.
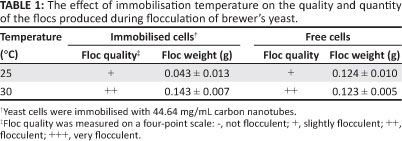
Table 1 indicates that the ideal temperature for flocculation using CNTs was close to 30 °C. That is, the weight of the floc produced at 30 °C was significantly greater (0.143 ± 0.007 g) than that produced at 25 °C (0.043 ± 0.013 g). In the absence of CNTs, flocculation weight was almost the same (i.e. 0.124 ± 0.010 g at 25 °C compared with 0.123 ± 0.005 g at 30 °C). Öztop et al.24 found the optimum temperature for immobilisation of yeast cells on a chitosan film to be 25 °C.
Similarly to our results which showed an increase in flocculation with an increase in temperature, Jin et al.20,21 and Hsu et al.31 also observed an increase in flocculation with an increase in temperature from 5 °C to 45 °C.
Effect of carbon nanotube concentration
Different concentrations (from 0 ìg/mL to 72 ìg/mL) of CNTs were added to the broth containing yeast cells and culture medium to investigate the effect of concentration of CNTs on flocculation. Table 2 and Figure 4 show the change in floc weight and quality observed with a change in CNT concentration.
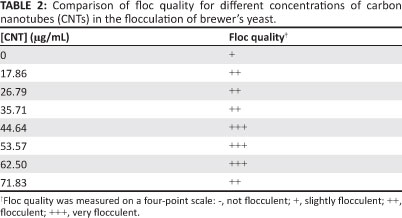
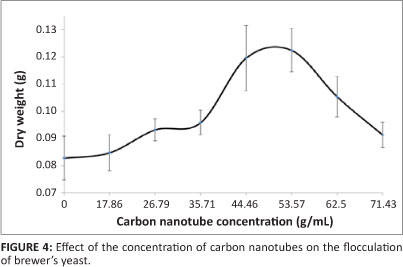
There was a general increase in floc weight with an increase in CNT concentration, with a peak at a CNT concentration of about 53 ìg/mL. Increases in CNT concentration beyond 53.57 ìg/mL caused a decrease in the floc weight, showing a negative effect of CNT concentration on flocculation beyond a certain concentration threshold. From concentrations of 0 ìg/mL to 35.71 ìg/mL, there was a negligible gain in floc weight (0.013 g). Increasing the CNT concentration to 53.57 ìg/mL resulted in a gain of 0.040 g.
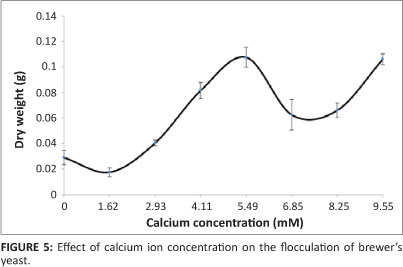
Effect of calcium ion concentration
Taylor and Orton32 observed that the presence of calcium ions at a very low concentration induced flocculation, whilst at high concentrations flocculation was inhibited. The influence of calcium ions was also tested in the present study. Calcium ion concentration was varied from 0 mM to 9.55 mM and introduced into the broth as anhydrous calcium chloride (CaCl2x2H2O). The calcium chloride weight was varied in steps of 0.05 g to yield seven concentrations for testing. The results are presented in Figure 5. The best flocculation quality was observed at Ca2+ concentrations of 5.49 mM and 9.55 mM. Figure 5 shows that flocculation typically increased from 0 mM to a peak at 5.49 mM, whereafter it decreased and increased again to the same peak at 9.55 mM.
Presence of glucose
The effect of glucose on flocculation of brewer's yeast cells was another parameter investigated. Generally, it has been found that maltose and mannose are the most effective inhibitors of flocculation whereas sucrose and glucose are less effective.19
The presence of glucose promoted yeast cell growth and delayed the stationary phase for yeast cells, thereby delaying the onset of flocculation. Ethanol was produced from the effect of the yeast cells on glucose, which decreased the pH of the broth and thus resulted in a delay in flocculation. The amount of glucose added is usually between 3 and 5 times the weight of the medium15,17,33,34; the glucose concentration used was 18 mg/mL.
The findings were plotted as pH and zeta potential against time (Figure 6). The results show a decrease in pH from 5.53 to 3.84 within a day and a progressive increase thereafter. A pH of 5.60, which is the optimum pH for the onset of flocculation, was reached after 3.95 days (~4 days).
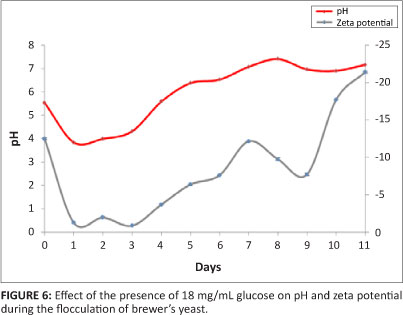
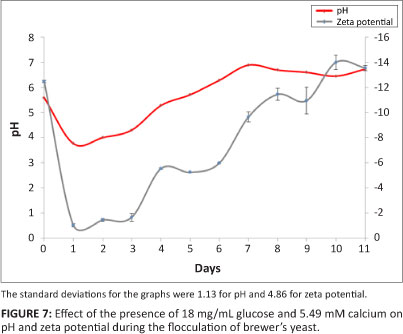
The experiment was repeated with 5.49 mM calcium ions (Figure 7) and there was a decrease in pH from 5.59 to 3.76 within a day and a progressive increase thereafter. A pH of 5.60 was reached after 4.90 days (~5 days), that is, there was a delay of 5 days before flocculation was observed. These observations were in agreement with previous reports which state that glucose inhibits flocculation.2,19,35 Several authors have indeed found that flocculation is triggered by carbon and/or nitrogen starvation and that the addition of these compounds to the growth medium delays flocculation.22,23,36
Effect of agitation speed
An analysis of the effect of agitation speed on the immobilisation of brewer's yeast was carried out by changing the speed from 0 to 200 revolutions per minute (rpm). A poor flocculation (+) was observed for speeds of 0 rpm, 50 rpm, 150 rpm and 200 rpm whilst good flocculation (++) was observed at a speed of 110 rpm (Table 3).
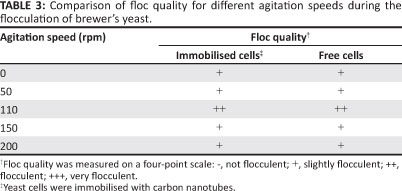
The weak flocculation observed at higher agitation speeds of 150 rpm and 200 rpm (Table 3) may be as a result of disintegration of the flocs. While an increase in collisions may help to grow the flocs, there is a limit to the agitation speed beyond which surface erosion or floc fracture sets in, which limits the stable floc to a certain optimum size.37
Stratford and Wilson37 showed that flocculation was observed at shaking speeds between 65 rpm and 115 rpm; these observation are in agreement with those of the present study where very little flocculation was observed at speeds lower than 50 rpm and optimum flocculation occurred at about 110 rpm.
The presence of CNTs increased the flocculation rate of brewer's yeast. In addition, the recovery of the flocs by freeze drying demonstrated that the flocs immobilised on CNTs were more stable than those produced by free cells.
The observation that CNTs increase the flocculation rate of yeast cells could possibly be explained by considering the Bridging Mechanism Theory. CNTs could be considered to be long chain particles which have large surface spikes which enable the neutralisation of the surface charge of brewer's yeast cells when there is contact made between the cells and the CNTs. This contact would allow the cells to adsorb onto the tubes such that an individual chain can become attached to two or more cells, 'bridging' them together. Spike structures accumulate tip-charge, but the energy required to push a spike tip through a repulsion field would be considerably less than that for cell-cell wall contact. The spike may contain a positive tip charge (as is the case with CNTs), which would most easily penetrate the negative charge repulsion of the yeast cells.37
The presence of CNTs also could have increased the water contact angle leading to an increase in cell surface hydrophobicity, which in turn initiated flocculation.38,39 A high level of cell surface hydrophobicity may facilitate cell-cell contact in an aqueous medium, resulting in more specific lectin-carbohydrate interactions.37
Conclusions
We have demonstrated the potential of CNTs for improving flocculation of brewer's yeast. The influence of various factors affecting flocculation of brewer's yeast on CNTs was studied. The optimum flocculation conditions were found to be: a pH between 5.00 and 5.80, a temperature of about 30 °C, an agitation speed of about 110 rpm, a concentration of CNTs of between 44 ìg/mL and 54 ìg/mL, and a calcium ion concentration of 5.49 mM. The addition of glucose decreased the flocculation rate and delayed the onset of flocculation.
Acknowledgements
The work presented was made possible with the assistance of the National Research Foundation (South Africa), Food-Bev Seta and the University of the Witwatersrand.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this paper.
Authors' contributions
T.A.M. was responsible for the experimental work and writing the manuscript. S.I., V.S. and C.S.Y. were responsible for supervision and guidance.
References
1. Pilkington PH, Margaritis A, Mensour NA, Russell I. Fundamentals of immobilized yeast cells for continuous beer fermentation: a review. J Inst Brew. 1998;104:19-31. [ Links ]
2. Domingues L, Vincente AA, Lima N, Teixeira JA. Applications of yeast flocculation in biotechnological processes. Biotechnol Bioprocess Eng. 2000;5:288-305. http://dx.doi.org/10.1007/BF02942185 [ Links ]
3. Stewart GG, Russell I. Brewing science. Toronto: Academic Press; 1981. [ Links ]
4. Berger M. Ethanol production inside carbon nanotubes [homepage on the Internet]. c2009 [updated 2007 June 08, cited 2009 Aug 03]. Available from http://www.nanowerk.com/spotlight/spotid=2053.php. [ Links ]
5. Kourkoutas Y, Bekatorou A, Banat IM, Marchant R, Koutinas AA. Immobilization technologies and support materials suitable in alcohol beverages production: A review. Food Microbiol. 2004;21:377-397. http://dx.doi.org/10.1016/j.fm.2003.10.005 [ Links ]
6. O'Connor L. Innovation in the lab: Emerging material could provide the breakthrough that nanotech needs. Global Research Technical Insights. 2009; 2(1), 3 pages. [ Links ]
7. Briggs DE, Boulton CA, Brookes PA, Stevens R. An outline of brewing: Brewing science and practice. Cambridge: Woodhead Publishing Ltd; 2004. [ Links ]
8. Van Zandycke S, Siddique R, Smart KA. The role of the membrane in predicting yeast quality. MBAA TQ. 2003;40(3):169-173. [ Links ]
9. Converti A, Borghi MD, Ferraiolo G, Sommariva C. Mechanical mixing and biological deactivation: The role of shear stress application time. Chem Eng J. 1996;62:155-167. [ Links ]
10. Firon N, Ofek I, Sharon N. Interaction of mannose-containing oligosaccharides with the fimbrial lectin of Escherichia coli. Biochem Biophys Res Commun. 1982;105:1426-1432. http://dx.doi.org/10.1016/0006-291X(82)90947-0 [ Links ]
11. Iyuke SE, Mamvura TA, Liu K, Sibanda V, Meyyappan M, Varadan VK. Process synthesis and optimisation for the production of carbon nanostructures. Nanotechnology. 2009;20:375602, 10 pages. [ Links ]
12. Brányik T, Vicente A, Oliveira R, Teixeira J. Physicochemical surface properties of brewing yeast influencing their immobilisation onto spent grains in a continuous reactor. Biotechnol Bioeng. 2004;88:84-93. http://dx.doi.org/10.1002/bit.20217,PMid:15389484 [ Links ]
13. Peinado RA, Moreno JJ, Maestre O, Mauricio JC. Use of a novel immobilization yeast system for winemaking. Biotechnol Lett. 2005;27:1421-1424. http://dx.doi.org/10.1007/s10529-005-0939-2, PMid:16215861 [ Links ]
14. Sakurai A, Nishida Y, Saito H, Sakakibara M. Ethanol production by repeated batch culture using yeast cells immobilized within porous cellulose carriers. J Biosci Bioeng. 2000;90(5):526-529. PMid:16232903 [ Links ]
15. Gztop HN, Öztop AY, Karadag E, I§ikver Y, Saraydin D. Immobilisation of Saccharomyces cerevisiae on to acrylamide-sodium acrylate hydrogels for production of ethyl alcohol. Enzyme Microb Technol. 2003;32:114-119. http://dx.doi.org/10.1016/S0141-0229(02)00244-2 [ Links ]
16. Hussain T, Salhi O, Lematre J, Charpentier C, Bonaly R. Comparative studies of flocculation and deflocculation of Saccharomyces uvarum and Kluyveromyces bulgaricus. Appl Microbiol Biotechnol. 1986;23:269-273. http://dx.doi.org/10.1007/BF00261927 [ Links ]
17. Nahvi I, Emtiazi G, Alkabi L. Isolation of a flocculating Saccharomyces cerevisiae and investigation of its performance in the fermentation of beet molasses to ethanol. Biomass Bioenergy. 2002;23:481-486. http://dx.doi.org/10.1016/S0961-9534(02)00070-3 [ Links ]
18. Ross AH, Harrison JS. The yeasts. London: Academic Press, 1970; p. 148156. [ Links ]
19. Jin Y-L, Speers RA. Flocculation of Saccharomyces cerevisiae. Food Res Int.1999;31:421-440. http://dx.doi.org/10.1016/S0963-9969(99)00021-6 [ Links ]
20. Jin Y-L, Speers RA. Effect of environmental conditions on the flocculation of Saccharomyces cerevisiae. J Am Soc Brew Chem. 2000;58:108-116. [ Links ]
21. Jin Y-L, Ritcey LL, Speers RAR, Dolphin PJ. Effect of cell surface hydrophobicity, charge and zymolectin density on the flocculation of Saccharomyces cerevisiae. J Am Soc Brew Chem. 2001;59:1-9. [ Links ]
22. Soares EV, Texeira JA, Mota M. Effect of cultural and nutritional conditions on the control of flocculation expression in Saccharomyces cerevisiae. Can J Microbiol. 1994;40:851-857. http://dx.doi.org/10.1139/m94-135, PMid:8000963 [ Links ]
23. Stratford M. Yeast flocculation: A new perspective. In: Rose AH, Tempest D, editors. Advances in microbial physiology. New York: Academic Press, 1992; p. 1-71. [ Links ]
24. Gztop HN, Saraydin D, Cetinus S. pH-sensitive chitosan films for baker's yeast immobilization. Appl Biochem Biotechnol. 2002;101:239-249. http://dx.doi.org/10.1385/ABAB:101:3:239 [ Links ]
25. Speers RA, Tung MA, Durances TD, Stewart GG. Biochemical aspects of yeast flocculation and its measurement: A review. J Inst Brew. 1992;98:293-300. [ Links ]
26. Verstrepen KJ, Derdelinckx G, Verachtert H, Delvaux FR. Yeast flocculation: What brewers should know. Appl Microbiol Biotechnol. 2003;61:197-205. PMid:12698276 [ Links ]
27. Zhao J, Fleet GH. Degradation of DNA during the autolysis of Saccharomyces cerevisiae. J Ind Microbiol Biotechnol. 2003;30:175-182. PMid:12715255 [ Links ]
28. Arnold WN. Autolysis in yeast cell envelopes: Biochemistry, biophysics and ultrastructure. New York: CRC Press, 1981; p. 129-137. [ Links ]
29. Hernawan T, Fleet G. Chemical and cytological changes during the autolysis of yeasts. J Ind Microbiol. 1995;14:440-450. http://dx.doi.org/10.1007/BF01573955, PMid:7662285 [ Links ]
30. Alexandre H, Guilloux-Benatier M. Yeast autolysis in sparkling wine - A review. Aust J Grape Wine Res. 2006;12:119-127. http://dx.doi.org/10.1111/j.1755-0238.2006.tb00051.x [ Links ]
31. Hsu JWC, Speers RA, Paulson AT. Modeling of orthokinetic flocculation of Saccharomyces cerevisiae. Biophys Chem. 2001;94:47-58. http://dx.doi.org/10.1016/S0301-4622(01)00236-8 [ Links ]
32. Taylor NW, Orton WL. Effect of alkaline earth metal salts on flocculence in Saccharomyces cerevisiae. J Inst Brew. 1973;79:294-297. [ Links ]
33. Dengis PB, Nelissen LR, Rouxhet PG. Mechanisms of yeast flocculation: Comparison of top- and bottom-fermenting strains. Appl Environ Microbiol. 1995;61:718-728. PMid:7574609, PMCid:167332 [ Links ]
34. Mallouchos A, Reppa P, Aggelis G, Kanellaki M, Koutinas AA, Komaitis M. Grape skins as a natural support for yeast immobilisation. BiotechnolLett. 2002;24:1331-1335. http://dx.doi.org/10.1023/A:1019892232296 [ Links ]
35. Verbelen PJ, De Schutter DP, Delvaux F, Verstrepen KJ, Delvaux FR. Immobilized yeast cell systems for continuous fermentation applications. Biotechnol Lett. 2006;28:1515-1525. http://dx.doi.org/10.1007/s10529-006-9132-5, PMid:16937245 [ Links ]
36. Soares EV, Mota M. Flocculation onset, growth phase and genealogical age in Saccharomyces cerevisiae. Can J Microbiol. 1996;42:539-547. http://dx.doi.org/10.1139/m96-073, PMid:8801005 [ Links ]
37. Stratford M, Wilson PDG. Agitation effects on microbial cell-cell interactions: A review. Lett Appl Microbiol. 1990;11:1-6. http://dx.doi.org/10.1111/j.1472-765X.1990.tb00122.x, PMid:1366670 [ Links ]
38. Smit G, Straver MH, Lugtenberg BJJ. Flocculence of Saccharomyces cerevisiae cells is induced by nutrient limitation with cell surface hydrophobicity as a major determinant. Appl Environ Microbiol. 1992;58(11):3709-3714. PMid:1482191, PMCid:183164 [ Links ]
39. Straver MH, Aar PC, Van der Smit G, Kijne JW. Determinants of flocculence of brewer's yeast during fermentation in wort. Yeast. 1993;9: 527-532. http://dx.doi.org/10.1002/yea.320090509, PMid:8322515 [ Links ]
 Correspondence to:
Correspondence to:
Tirivaviri Mamvura
School of Chemical and Metallurgical Engineering,
Faculty of Engineering and the Built Environment,
University of the Witwatersrand,
Wits 2050, South Africa
Email: atmamvura@yahoo.com
Received: 19 May 2011
Accepted: 10 Feb. 2012
Published: 17 July 2012
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.

