










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.108 n.5-6 Pretoria Jan. 2012
RESEARCH ARTICLES
Molecular characterisation of human peripheral blood stem cells
Ruzanna Ab KadirI; Shahrul Hisham Zainal AriffinI; Rohaya Megat Abdul WahabII; Sahidan SenafiI
ISchool of Biosciences and Biotechnology, Faculty of Science and Technology, National University of Malaysia, Bangi, Selangor, Malaysia
IIDepartment of Orthodontics, Faculty of Dentistry, National University of Malaysia, Bangi, Selangor, Malaysia
ABSTRACT
Peripheral blood mononucleated cells consist of haematopoietic stem cells (HSCs) and mesenchymal stem cells (MSCs). To date there is no well-defined isolation or characterisation protocol of stem cells from human adult peripheral blood mononucleated cells. Our aim in this study was to isolate and characterise mononucleated cells from human peripheral blood. Peripheral blood mononucleated cells were isolated using the Ficoll-Paque density gradient separation method and cultured in complete medium. After 4 days of culture, adherent and suspension mononucleated cells were separated and cultured for 14 days in an in-vitro culture selection. Stem cells in the isolated mononucleated cells were characterised using a multidisciplinary approach which was based on the expression of stem cell markers, morphology and the capacity to self-renew or proliferate and differentiate into specialised cells. Reverse transcription polymerase chain reaction was used to identify the expression of an HSC marker (signalling lymphocytic activation molecule family member 1, SLAMF1) and a MSC marker (CD105). Results revealed that adherent mononucleated cells were positive for MSC markers, whereas mononucleated cells in suspension were positive for HSC markers. The isolated adherent and suspension mononucleated cells were able to maintain their stem cell properties during in-vitro culture by retaining their capacity to proliferate and differentiate into osteoclast and osteoblast cells, respectively, when exposed to the appropriate induction medium. The isolated mononucleated cells consisted of suspension HSCs and adherent MSCs, both of which have the capability to proliferate and differentiate into mature cells. We have shown that suspension HSCs and adherent MSCs can be obtained from an in-vitro culture of peripheral blood mononucleated cells.
Introduction
Stem cells are unspecialised cells with the capacity to self-renew, differentiate and repopulate a host in vivo.1 Stem cells are widely studied as they are well known to have potential in the development of innovative therapeutic strategies. In general, there are two types of stem cells: embryonic and adult stem cells. Embryonic stem cells originate from the inner cell mass of the blastocyst stage during embryonic development and can differentiate into all cell types of the body because they are pluripotent.2,3 Adult stem cells are categorised as multipotent stem cells because they can differentiate into cell types different from their tissue of origin. Embryonic stem cells are more versatile in that they can develop into a greater variety of tissues than can adult stem cells. However, the therapeutic use of embryonic stem cells is still controversial because of ethical concerns, as well as immunological incompatibilities and concerns about uncontrolled development of malignancies or teratomas from administered cells.4 In contrast, the use of adult stem cells is free of such ethical concerns, and, because the use is autologous, there are no concerns regarding incompatibility and rejection.5
Mesenchymal stem cells (MSCs) and haematopoietic stem cells (HSCs) are adult stem cells present in peripheral blood. MSCs are generally defined as self-renewable, multipotent progenitor cells with the ability to differentiate into several mesenchymal lineages including bone, cartilage, adipose and muscle tissues. In addition, MSCs have also demonstrated the potential to transdifferentiate into other types of tissues such as hepatic, renal, cardiac and neuronal tissues, thus indicating the cells pluripotency.6 HSCs are defined by their ability to repopulate all of the haematopoietic lineages in vivo and sustain the production of these cells for the lifespan of the individual.7 HSCs have been or are being used for the restoration of lymphohaematopoietic function after myeloablative, near myeloablative or non-myeloablative treatment.8
Our main objectives for this study were the isolation, culture and molecular characterisation of mononucleated cells from human peripheral blood. Stem cells in the isolated mononucleated cells were characterised using a multidisciplinary approach which was based on the expression of stem cell markers, morphology and the capacity to self-renew or proliferate and differentiate into specialised cells as evidenced by proliferation and molecular analysis. Mononucleated cells consisting of MSCs, HSCs and their progenitors, along with monocytes, lymphocytes and macrophages were isolated from the buffy coat of blood. Our results demonstrated that suspension and adherent mononucleated cells showed molecular expressions of HSC markers and MSC markers, respectively. Furthermore, suspension and adherent mononucleated cells were still able to maintain their stem cell properties during in-vitro culture.
Material and methods
Isolation of human mononucleated cells
Mononucleated cells were isolated from healthy donors aged between 18 and 25 years old by density gradient centrifugation using Ficoll-PaqueTM PLUS (GE Healthcare, Uppsala, Sweden). Human peripheral blood samples were obtained after consent from the donors and approval from the ethical committee of the Faculty of Science and Technology, National University of Malaysia. The blood samples were diluted three times with Hanks' Balanced Salt Solution (Sigma, St Louis, MO, USA). The diluted blood samples were carefully layered 1:1.5 on Ficoll-PaqueTM PLUS and centrifuged at 400 g for 20 min at room temperature. The mononucleated cell layer at the plasma-Ficoll interface was washed three times with phosphate buffered saline and cultured in six-well plates with complete medium containing alpha medium essential medium, 2% (v/v) penicillin-streptomycin and 10% (v/v) newborn calf serum at 37 °C in a humidified atmosphere containing 5% CO2 for 4 days. After 4 days, non-adherent (suspension) mononucleated cells were transferred into new plates. Both adherent and suspension mononucleated cells were maintained in the complete medium with twice weekly medium exchange for 14 days in in-vitro culture before further use.
May-Grunwald Giemsa staining
May-Grunwald Giemsa staining was carried out with May-Grunwald's eosin methylene blue solution (Merck, Darmstadt, Germany) and Giemsa solution (Sigma, St Louis, MO, USA). May-Grunwald Giemsa staining started with slides immersed in 100% (v/v) May-Grunwald for 2.5 min. The slides were then transferred directly to 4% (v/v) Giemsa for 2.5 min and briefly rinsed with distilled water. Excess dye was wiped off and the slides were air-dried. The May-Grunwald Giemsa stained areas were viewed by light microscopy.
Proliferation of suspension and adherent mononucleated cells
Suspension and adherent mononucleated cells were cultured in complete medium. For suspension mononucleated cells, the trypan blue exclusion assay was used to observe the proliferation of the cells, whereas the 3-[4,5-dimethylthiazol-2-y]-2,5-diphenyltetrazolium bromide (MTT) assay was used to observe the proliferation of adherent mononucleated cells.
Statistical analysis
Data were statistically analysed using paired t-tests. Effects were considered statistically significant at p < 0.05.
In-vitro differentiations into osteoclast and osteoblast
The ability of adherent and suspension mononucleated cells to differentiate into specialised cells from their lineages was evaluated by differentiating the suspension and adherent mononucleated cells into osteoclast and osteoblast cells, respectively. Cells cultured in complete medium were supplemented with 50 ng/mL recombinant soluble receptor activator of nuclear factor kappa-B ligand (sRANKL) and 25 ng/mL macrophage colony-stimulating factor (M-CSF) to induce osteoclast differentiation, while 50 ìg/mL ascorbic acid and 10 mM â-glycerophosphate were added to induce differentiation into osteoblasts. The cultures were maintained at 37 °C in a fully humidified atmosphere containing 5% CO2. Some of the cells were cultured with complete medium without supplementation of growth factors to act as controls.
Reverse transcription polymerase chain reaction amplification
Total RNA was extracted using TRI ReagentTM (Sigma, St Louis, MO, USA) following the manufacturer's instructions. Total RNA was extracted from adherent and suspension mononucleated cells that had been cultured in complete medium and differentiation medium. Total RNA (1 ìg) was subjected to reverse transcription polymerase chain reaction (RT-PCR) amplification using an Access RT-PCR System kit provided by Promega (Madison, WI, USA). First-strand complementary DNA (cDNA) was synthesised by reverse transcription at 45 °C for 45 min, followed by avian myeloblastosis virus reverse transcriptase inactivation at 94 °C for 2 min. Second-strand cDNA synthesis and PCR amplification consisted of 40 cycles of denaturation at 94 °C for 30 s, primer annealing at 55 °C (GAPDH), 54 °C (SLAMF1) or 61 °C (CD105) for 1 min and extension at 68 °C for 2 min, with a final cycle at 68 °C for 7 min. The specific primer sequences used are shown in Table 1.
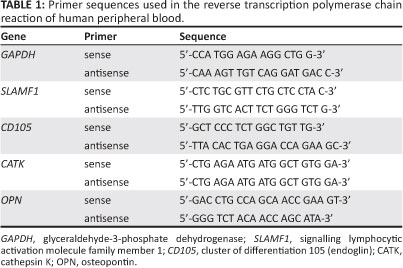
GAPDH was used as a positive control. RT-PCR amplifications were performed in a Mastercycler Gradient thermocycler (Eppendorf, Hamburg, Germany). The PCR products were separated by 1% (w/v) agarose gel electrophoresis. The separated DNA fragments were visualised by ethidium bromide staining and photographed using the Alpha Imaging System (Alpha Innotech, San Leandro, CA, USA) under UV light.
Results and discussion
Isolation of mononucleated cells
Mononucleated cells consisted of MSCs, HSCs and their progenitors, along with leukocytes and macrophages. Mononucleated cells were isolated from the buffy coat of peripheral blood using a density gradient centrifugation on Ficoll-Paque where differences in density separate mononucleated cells from other blood cells. However, isolated mononucleated cells are a heterogeneous population.
In this study, we further separated the mononucleated cells according to their physical characteristics. Non-adherent (suspension) and adherent cells were cultured separately after 4 days of isolation. Both adherent and suspension mononucleated cells were maintained in the complete medium for 14 days of culture for the purpose of separating adherent and suspension cells and also to deplete most of the unwanted cells (e.g. differentiated and precursor cells) that have a short lifespan, for example, granulocytes (30 min -40 min in peripheral blood with a total lifespan of 7-13 days depending on pathological state) and monocytes (5-7 days).9 Figure 1 shows the morphology of suspension and adherent mononucleated cells after 14 days of culture. Suspension cells without staining appear morphologically rounded (Figure 1a). The morphology of the suspension cells after staining included a large single nucleus that occupied much of the cytoplasmic space, resembling that of lymphocytes. MSCs are cells that adhere to the tissue culture plastic and display a fibroblastic appearance10; adherent mononucleated cells showing spindle-shaped fibroblast-like morphology can be seen in Figure 1b. After May-Grunwald Giemsa staining was carried out on the suspension cells fraction, the nuclei were stained dark purple and the cytoplasms were stained a lighter purple (Figure 1c).
Expression of stem cell markers
Physical characteristics alone are not sufficient to prove that HSCs and MSCs are present in mononucleated cells. The existence of HSC and MSC mononucleated cells was further characterised by a molecular approach (RT-PCR analysis) using specific HSC and MSC markers. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is one of the most commonly used housekeeping genes used in comparisons of gene expression data as it is constitutively expressed at the same level in mammalian cells and tissues.11 Therefore in this study, GAPDH was used as a positive control for RT-PCR analysis.
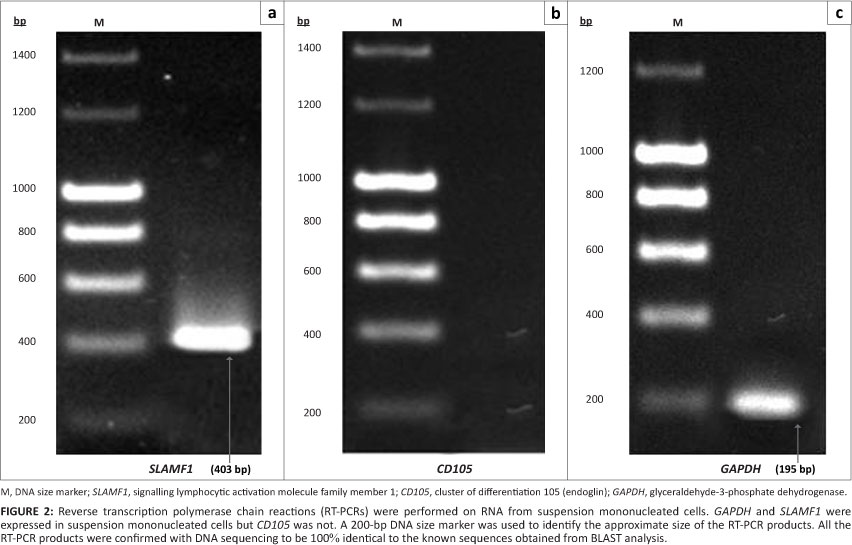
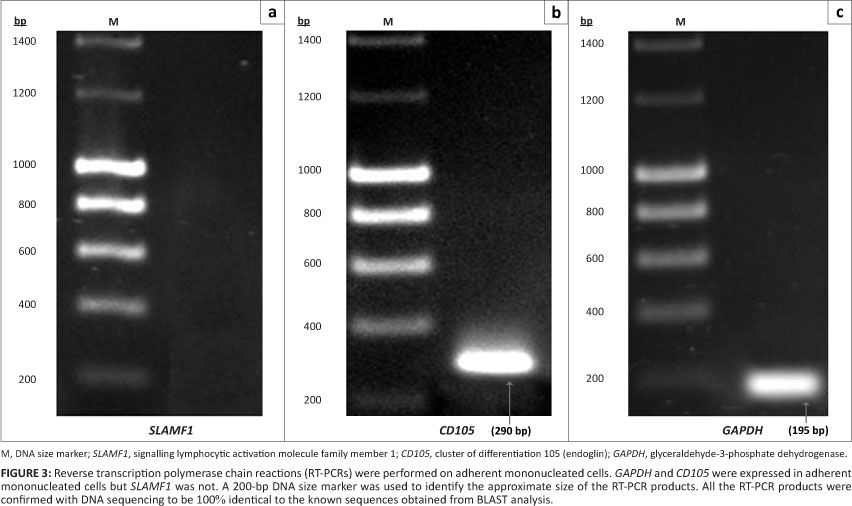
As shown in Figure 2, the suspension mononucleated cells were positive for the presence of an HSC marker (signalling lymphocytic activation molecule family member 1, SLAMF1) as well as GAPDH with RT-PCR products of the expected size for SLAMF1 (403 bp) and GAPDH (195 bp). Figure 3 shows amplification of the RT-PCR products of GAPDH and a MSC marker (CD105) in RNA from adherent mononucleated cells. SLAMF1 and CD105 were not expressed in adherent and suspension mononucleated cells, respectively.
CD34 is the most commonly used marker for haematopoietic stem cells. However, not all haematopoietic stem cells are CD34+ cells. According to Bhatia et al.12 and Guo et al.13, the CD34 antigen does not appear to be expressed on all HSCs. Although CD34 is considered to be a HSC marker, the CD34 antigen is also expressed on committed progenitors14 and non-haematopoietic progenitors.15 Therefore, in this study, an alternative HSC marker (SLAMF1) was used and evaluated. SLAMF1 is a cell surface receptor that is important during the self-renewal of HSCs.16 SLAMF1 is the founding member of the SLAM family of cell surface receptors.17,18 According to Kiel et al.19, SLAMF1 is only expressed by HSCs and not by multipotent haematopoietic progenitors or haematopoietic progenitor cells. Moreover, SLAM family members regulate the proliferation and activation of lymphocytes.20,21 Like SLAMF1, CD105 is also involved in regulating proliferation but only in MSCs.22 CD105 or endoglin is a type I membrane glycoprotein, which is located on the cell surface and is also part of the TGF-â receptor complex.23
Proliferation and differentiation of mononucleated cells
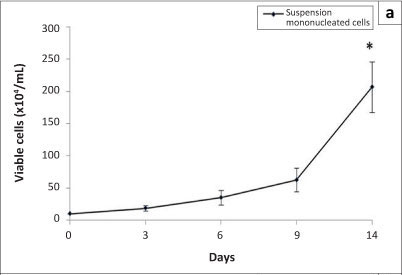
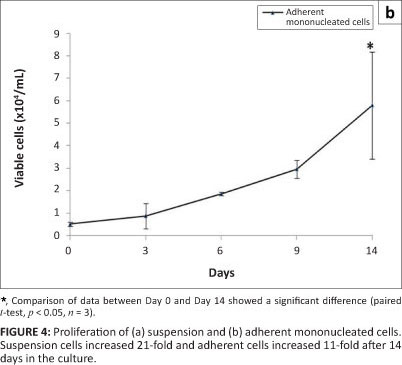
Stem cells possess two properties: the ability to self-renew or proliferate and the ability to differentiate into specialised cells. In this study, the proliferation of suspension and adherent mononucleated cells in an in-vitro culture was investigated using the trypan blue exclusion assay and the MTT assay, respectively. Figure 4 shows the proliferation of suspension and adherent mononucleated cells. Statistical analyses comparing data from Day 0 and Day 14 showed a significant increase (p < 0.05) in viable cell numbers in both cell populations. From Figure 4a, a significant increase of an average of 21-fold in viable cells was observed on Day 14 compared to Day 0. The number of adherent cells also increased significantly by an average of 11-fold (Figure 4b). These results show that suspension and adherent mononucleated cells were able to proliferate during in-vitro culture conditions.
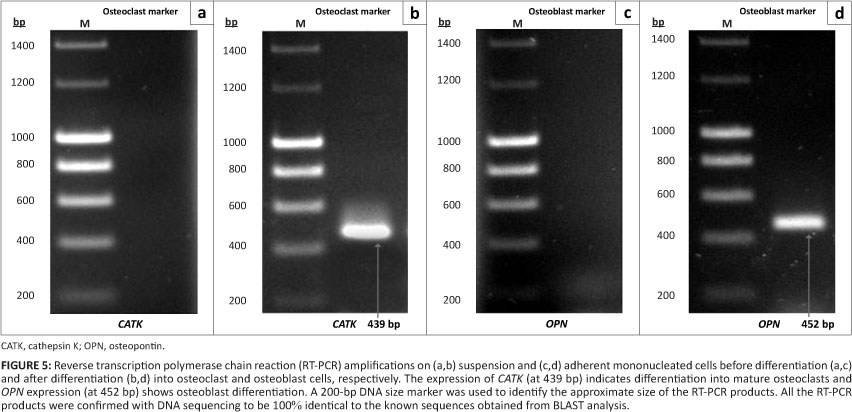
The differentiation of stem cells into specialised cell types is one of the important properties of stem cells. In order to examine the ability of MSCs from adherent mononucleated cells and HSCs from suspension mononucleated cells to differentiate into specialised cells from their lineages, suspension and adherent mononucleated cells were differentiated toward osteoclast and osteoblast cells, respectively, by adding specific differentiation factors to the medium. sRANKL and M-CSF were added to induce differentiation into osteoclasts while ascorbic acid and -glycerophosphate were added to induce differentiation into osteoblasts. Molecular analysis was performed to determine in-vitro differentiation of suspension and adherent mononucleated cells into mature osteoclast and osteoblast cells, respectively (Figure 5).
A number of genes such as tartrate resistant acid phosphatase (TRAP) and cathepsin K (CATK) are expressed during osteoclastogenesis.24 All of these genes have been functionally implicated in osteoclastic bone resorption.25 As shown in Figure 5b, expression of CATK was observed by RT-PCR analysis in suspension mononucleated cells cultured in osteoclast differentiation medium, which contained M-CSF and sRANKL. The results presented here also show that suspension mononucleated cells in complete medium without supplementation of growth factors were negative for CATK. CATK expression is important during osteoclast differentiation26 as the function of CATK in osteoclasts accounts for their osteoclastic activities.27 Osteoclast cells function in bone resorption by removing the mineralised matrix and breaking up the organic bone. Osteoclast cells secrete hydrochloric acid, which is involved in hydroxyapatite resorption and protein degradation by proteases such as CATK. CATK functions to degrade collagen and other bone matrix proteins.28
Osteopontin (OPN) is considered an important marker in osteoblastic differentiation.29 Using RT-PCR analysis, we observed that adherent mononucleated cells cultured in osteoblast differentiation medium were positive for OPN expression, in contrast to cells cultured in complete medium without supplementation of growth factors (Figure 5b). Osteoblast cells are essential in bone development. During bone development, OPN is expressed at an early stage of bone formation,30 laid into unmineralised matrix before calcification, and localised at matrix-matrix and matrix-cell interfaces as well as between collagen fibrils of fully matured hard tissue.31
Conclusion
Our results indicate that there is a population of stem cells in the peripheral blood mononucleated cell fraction. The isolated mononucleated cells consist of HSC (suspension) and MSC (adherent) components. Moreover, the isolated stem cells amongst adherent and suspension mononucleated cells are still able to maintain their stem cell properties during in-vitro culture by retaining their capacity to self-renew or proliferate and differentiate into specialised cells.
Acknowledgements
This study was supported by grants from the Ministry of Higher Education, Malaysia (FRGS/1/2011/SG/ UKM/02/13 and UKM-DD-03-FRGS0030-2010), the Ministry of Science, Technology and Innovation (MOSTI), Malaysia (09-05-MGI-GMB002), the Department of Fishery Malaysia (STGL-007-2010/9) and the National University of Malaysia (UKM-OUP-KPB-33-170/2011 and UKM-GUP-2011-093).
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
R.A.K. conducted the laboratory work, generated and analysed the data, and wrote the manuscript. R.M.A.W. and S.S. assisted in the analysis of the data. S.H.Z.A. designed the study, analysed the data and contributed to the writing of the manuscript.
References
1. Weissman I, Anderson D, Gage F. Stem and progenitor cells: Origins, phenotypes, lineage commitments, and transdifferentiations. Annu Rev Cell Dev Biol. 2001;17:387-403. http://dx.doi.org/10.1146/annurev.cellbio.17.1.387, PMid: 11687494 [ Links ]
2. Shahrul Hisham ZA, Intan Zarina ZA, Sahidan S, Nor Muhammad M, Rohaya MAW, Zaidah ZA. Stem cells in blood development. Sains Malaysiana. 2005;34(1):21-26. [ Links ]
3. Shahrul Hisham ZA, Rohaya MAW, Ismanizan I, Nor Muhammad M, Zaidah ZA. Stem cells, cytokines and their receptors. As Pac J Mol Biol Biotechnol. 2005;13(1):1-13. [ Links ]
4. Hentze H, Graichen R, Colman A. Cell therapy and the safety of embryonic stem cell-derived grafts. Trends Biotechnol. 2007;25(1):24-32. http://dx.doi.org/10.1016/j.tibtech.2006.10.010, PMid:17084475 [ Links ]
5. Togel F, Westenfelder C. Adult bone marrow-derived stem cells for organ regeneration and repair. Dev Dyn. 2007;236(12):3321-3331. http://dx.doi.org/10.1002/dvdy.21258, PMid:17685479 [ Links ]
6. Alhadlaq A, Mao JJ. Mesenchymal stem cells: Isolation and therapeutics. Stem Cells Dev. 2004;13(4):436-448. http://dx.doi.org/10.1089/scd.2004.13.436, PMid: 15345137 [ Links ]
7. Wognum AW, Eaves AC, Thomas TE. Identification and isolation of hematopoietic stem cells. Arch Med Res. 2003;34(6):461-475. http://dx.doi.org/10.1016/j.arcmed.2003.09.008, PMid:14734086 [ Links ]
8. Arslan O, Moog R. Mobilization of peripheral blood stem cells. Transfus Apher Sci. 2007;37(2):179-185. http://dx.doi.org/10.1016/j.transci.2007.08.002, PMid:17980665 [ Links ]
9. Ariffin SH, Abidin IZ, Yazid MD, Wahab RM. Differentiation analyses of adult suspension mononucleated peripheral blood cells of Mus musculus. Cell Commun Signal. 2010;8:29. http://dx.doi.org/10.1186/1478-811X-8-29, PMid:20969794 [ Links ]
10. Bobis S, Jarocha D, Majka M. Mesenchymal stem cells: Characteristics and clinical applications. Folia Histochem Cytobiol. 2006;44(4):215-230. PMid:17219716 [ Links ]
11. Barber RD, Harmer DW, Coleman RA, Clark BJ. GAPDH as a housekeeping gene: Analysis of GAPDH mRNA expression in a panel of 72 human tissues. Physiol Genomics. 2005;21(3):389-395. http://dx.doi.org/10.1152/physiolgenomics.00025.2005, PMid:15769908 [ Links ]
12. Bhatia M, Bonnet D, Murdoch B, Gan OI, Dick JE. A newly discovered class of human hematopoietic cells with SCID-repopulating activity. Nat Med. 1998;4(9):1038-1045. http://dx.doi.org/10.1038/2023, PMid: 9734397 [ Links ]
13. Guo Y, Lübbert M, Engelhardt M. CD34 hematopoietic stem cells: Current concepts and controversies. Stem Cells. 2003;21(1):15-20. http://dx.doi.org/10.1634/stemcells.21-1-15, PMid: 12529547 [ Links ]
14. Larochelle A, Vormoor J, Hanenberg H, et al. Identification of primitive human hematopoietic cells capable of repopulating NOD/SCID mouse bone marrow: Implications for gene therapy. Nat Med. 1996;2(12):1329-1337. http://dx.doi.org/10.1038/nm1296-1329, PMid: 8946831 [ Links ]
15. Peichev M, Naiyer AJ, Pereira D, et al. Expression of VEGFR-2 and AC133 by circulating human CD34(+) cells identifies a population of functional endothelial precursors. Blood. 2000;95(3):952-958. PMid: 10648408 [ Links ]
16. Kent DG, Copley MR, Benz C, et al. Prospective isolation and molecular characterization of hematopoietic stem cells with durable self-renewal potential. Blood. 2009;113(25):6342-6350. http://dx.doi.org/10.1182/blood-2008-12-192054, PMid:19377048 [ Links ]
17. Engel P, Eck MJ, Terhorst C. The SAP and SLAM families in immune responses and X-linked lymphoproliferative disease. Nat Rev Immunol. 2003;3(10):813-821. http://dx.doi.org/10.1038/nri1202, PMid:14523387 [ Links ]
18. Sidorenko SP, Clark EA. The dual-function CD150 receptor subfamily: The viral attraction. Nat Immunol. 2003;4(1):19-24. http://dx.doi.org/10.1038/ni0103-19, PMid:12496974 [ Links ]
19. Kiel MJ, Yilmaz OH, Iwashita T, Yilmaz OH, Terhorst C, Morrison SJ. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell. 2005;121(7):1109-1121. http://dx.doi.org/10.1016/j.cell.2005.05.026, PMid: 15989959 [ Links ]
20. Howie D, Okamoto S, Rietdijk S, et al. The role of SAP in murine CD150 (SLAM)-mediated T-cell proliferation and interferon gamma production. Blood. 2002;100(8):2899-2907. http://dx.doi.org/10.1182/blood-2002-02-0445, PMid:12351401 [ Links ]
21. Wang N, Satoskar A, Faubion W, et al. The cell surface receptor SLAM controls T cell and macrophage functions. J Exp Med. 2004;199(9):1255-1264. http://dx.doi.org/10.1084/jem.20031835, PMid:15123745 [ Links ]
22. Gaebel R, Furlani D, Sorg H, et al. Cell origin of human mesenchymal stem cells determines a different healing performance in cardiac regeneration. PLoS One. 2011;6(2):e15652. http://dx.doi.org/10.1371/journal.pone.0015652, PMid:21347366 [ Links ]
23. Duff SE, Li C, Garland JM, Kumar S. CD105 is important for angiogenesis: Evidence and potential applications. FASEB J. 2003;17(9):984-992. http://dx.doi.org/10.1096/fj.02-0634rev, PMid:12773481 [ Links ]
24. Shi Z, Silveira A, Patel P, Feng X. YY1 is involved in RANKL-induced transcription of the tartrate-resistant acid phosphatase gene in osteoclast differentiation. Gene. 2004;343(1):117-126. http://dx.doi.org/10.1016/j.gene.2004.08.009, PMid:15563837 [ Links ]
25. FengX, Novack DV, Faccio R, et al. A Glanzmann's mutation in beta 3 integrin specifically impairs osteoclast function. J Clin Inves. 2001;107(9):1137-1144. http://dx.doi.org/10.1172/JCI12040, PMid:11342577 [ Links ]
26. Boyle WJ, Simonet WS, Lacey DL. Osteoclast differentiation and activation. Nature. 2003;423(6937):337-342. http://dx.doi.org/10.1038/nature01658, PMid:12748652 [ Links ]
27. Yazid MD, Ariffin SH, Senafi S, Razak MA, Wahab RM. Determination of the differentiation capacities of murines' primary mononucleated cells and MC3T3-E1 cells. Cancer Cell Int. 2010;10:42. http://dx.doi.org/10.1186/1475-2867-10-42, PMid:20979664 [ Links ]
28.Matsuo K, Irie N. Osteoclast-osteoblast communication. Arch Biochem Biophys. 2008;473(2):201-209. http://dx.doi.org/10.1016/j.abb.2008.03.027, PMid:18406338 [ Links ]
29. Kartsogiannis V, Ng KW. Cell lines and primary cells cultures in the study of bone cell biology. Mol Cell Endocrinol. 2004;228(1-2):79-102. http://dx.doi.org/10.1016/j.mce.2003.06.002, PMid:15541574 [ Links ]
30. Valenti MT, Dalle Carbonare L, Donatelli L, Bertoldo F, Zanatta M, Lo Cascio V. Gene expression analysis in osteoblastic differentiation from peripheral blood mesenchymal stem cells. Bone. 2008;43(6):1084-1092. http://dx.doi.org/10.1016/j.bone.2008.07.252, PMid:18761114 [ Links ]
31. Li Z, Zhou Z, Saunders MM, Donahue HJ. Modulation of connexin43 alters expression of osteoblastic differentiation markers. J Cell Physiol. 2006;290:1248-1255. http://dx.doi.org/10.1152/ajpcell.00428.2005, PMid:16319124 [ Links ]
 Correspondence to:
Correspondence to:
Shahrul Hisham Zainal Ariffin
Postal address: School of Biosciences and Biotechnology
Faculty of Science and Technology
National University of Malaysia
43600 Bangi, Selangor Darul Ehsan, Malaysia
Email: hisham@ukm.my
Received: 19 Sept. 2011
Accepted: 23 Dec. 2011
Published: 24 May 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}