










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.108 n.3-4 Pretoria Jan. 2012
RESEARCH ARTICLE
New hominin fossils from Kanapoi, Kenya, and the mosaic evolution of canine teeth in early hominins
Fredrick K. ManthiI; J. Michael PlavcanII; Carol V. WardIII
IDepartment of Earth Sciences, National Museums of Kenya, Nairobi, Kenya
IIDepartment of Anthropology, University of Arkansas, Fayetteville, USA
IIIDepartment of Pathology and Anatomical Sciences, University of Missouri, Columbia, USA
ABSTRACT
Whilst reduced size, altered shape and diminished sexual dimorphism of the canine-premolar complex are diagnostic features of the hominin clade, little is known about the rate and timing of changes in canine size and shape in early hominins. The earliest Australopithecus, Australopithecus anamensis, had canine crowns similar in size to those of its descendant Australopithecus afarensis, but a single large root alveolus has suggested that this species may have had larger and more dimorphic canines than previously recognised. Here we present three new associated dentitions attributed to A. anamensis, recently recovered from the type site of Kanapoi, Kenya, that provide evidence of canine evolution in early Australopithecus. These fossils include the largest mandibular canine root in the hominin fossil record. We demonstrate that, although canine crown height did not differ between these species, A. anamensis had larger and more dimorphic roots, more like those of extant great apes and Ardipithecus ramidus, than those of A. afarensis. The canine and premolar occlusal shapes of A. anamensis also resemble those of Ar. ramidus, and are intermediary between extant great apes and A. afarensis. A. afarensis achieved Homo-like maxillary crown basal proportions without a reduction in crown height. Thus, canine crown size and dimorphism remained stable during the early evolution of Australopithecus, but mandibular root dimensions changed only later within the A. anamensis-afarensis lineage, coincident with morphological changes in the canine-premolar complex. These observations suggest that selection on canine tooth crown height, shape and root dimensions was not coupled in early hominin evolution, and was not part of an integrated adaptive package.
Introduction
One of the earliest derived features of the hominin clade is canine tooth size reduction, with a decrease in sexual dimorphism in canine crown height, and the loss of maxillary canine tooth 'honing' against the lower third premolar that occurs in most primate species. Canine tooth crown reduction was originally thought to have first appeared in Australopithecus,1 but now is known to have characterised even earlier taxa - Sahelanthropus,2 Orrorin,3 Ardipithecus kadabba4,5,6 and Ardipithecus ramidus.7,8,9,10 However, the morphology of the Australopithecus canine-premolar complex is derived morphologically relative to these earlier hominins. Furthermore, canine tooth form appears to have changed throughout the early evolution of Australopithecus.9,11,12 The pattern and timing of canine evolution is significant for understanding early hominin evolution because alterations in canine tooth size and dimorphism constitute evidence of social and/or dietary adaptations.13,14
The earliest member of the Australopithecus-human clade is Australopithecus anamensis (4.17 Ma - 3.9 Ma).9,15,16,17,18 A. anamensis appears to represent the initial part of a lineage culminating in the better-known Australopithecus afarensis (3.77 Ma - 3.0 Ma).9,11,19 Compared to A. afarensis, A. anamensis had larger canine basal crown dimensions relative to postcanine tooth size, more ape-like canine and premolar shapes, and altered topography of the maxilla and mandible in the regions of the canine juga.11,16,17,18 The canine tooth crowns known for A. anamensis appear no more variable in their dimensions than those of either A. afarensis,17,18 or Ar. ramidus,7 which would seem to suggest that absolute canine crown height and breadth remained stable with minimal dimorphism throughout the origin and evolution of early Australopithecus.
However, a single large A. anamensis mandibular canine alveolus (KNM-KP 29287), and to some extent a large canine root with heavily worn crown from Fejej, Ethiopia (FJ-4-SB-1a),20 has led to the suggestion that there may have been more canine sexual dimorphism early in this lineage than is represented in the fossil record of preserved canine tooth crowns.17,18 This suggestion would be surprising, given that data from Ardipithecus, Orrorin and Sahelanthropus indicate that reduction in canine tooth crown height and breadth, as well as a decrease in dimorphism, are basal hominin traits.
The intermediate temporal position of A. anamensis, between the earlier Ardipithecus, Orrorin and Sahelanthropus, and A. afarensis, makes this species of great interest in documenting the rate and timing of changes in the canine-premolar complex. Whilst a gradual, integrated change in the complex5,9 might suggest a single vector of selective change to integrate canine function with that of the incisors, a mosaic pattern of change11 implies a pattern of sequential selective pressures and possible unappreciated functional diversity over time.
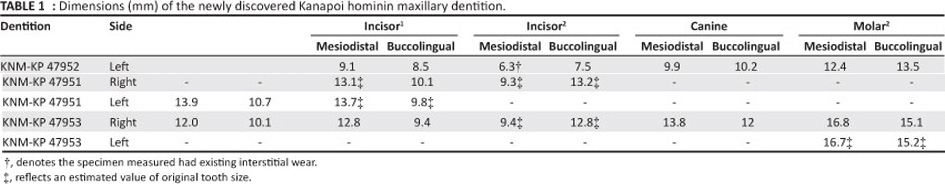
From 2003 to 2007, several new fossils attributed to A. anamensis were recovered from the type site of Kanapoi, Kenya by a team led by one of us (F.K.M.). The new Kanapoi hominin fossils include three partial, associated dentitions, each including a canine tooth (Figure 1; Table 1). All are from the lower fluvial sequence at the site, and are dated to between 4.195 Ma and 4.108 Ma.21 There are two mandibular dentitions: KNM-KP 47951 is a mandibular canine with associated premolars and KNM-KP 47953 is a mandibular dentition preserving the right canine and premolars, along with the second and third molar. KNM-KP 47952 is a maxillary dentition with two maxillary canines and an incisor.
These fossils provide important new evidence of canine evolution in early Australopithecus. Here we present these fossils and consider their implications for understanding canine tooth evolution in early hominins.
Materials and methods
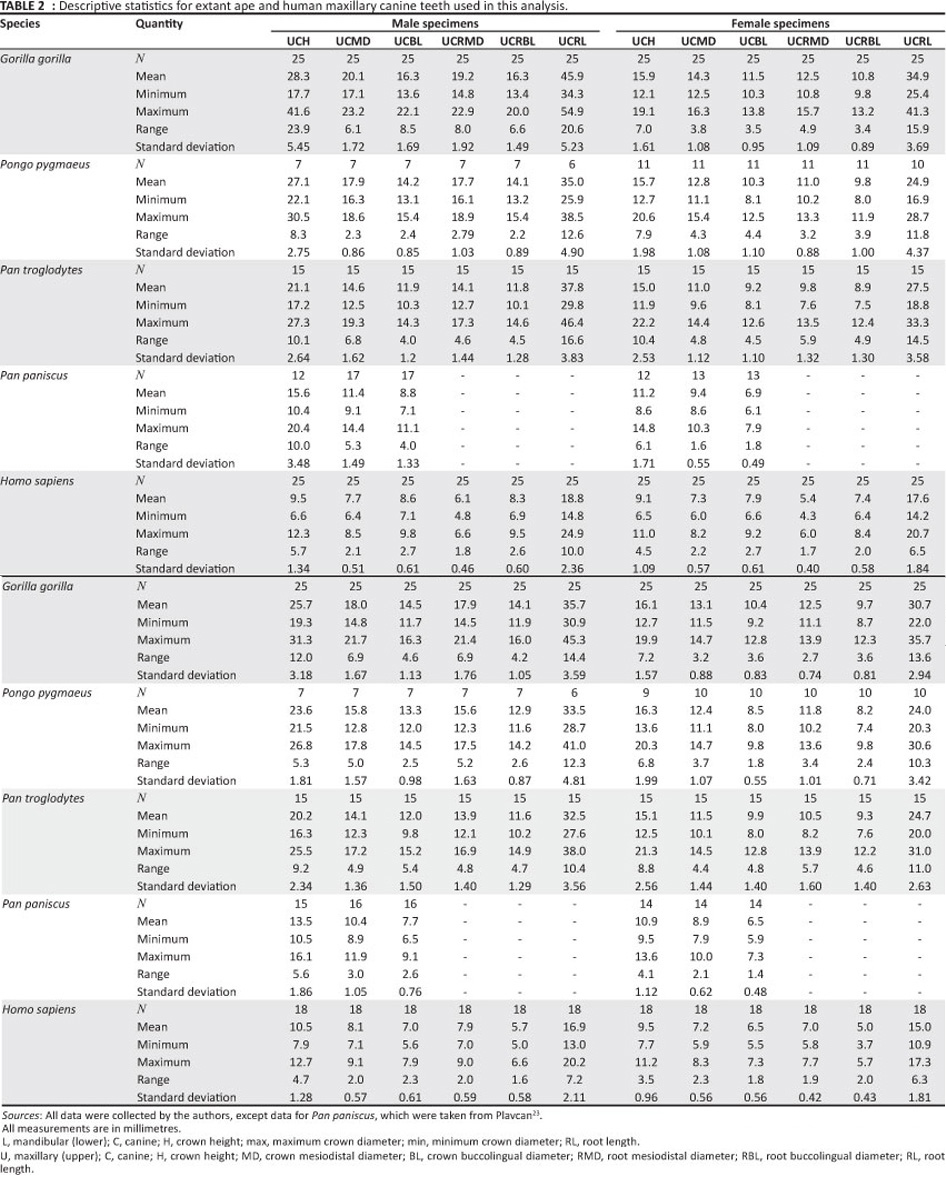
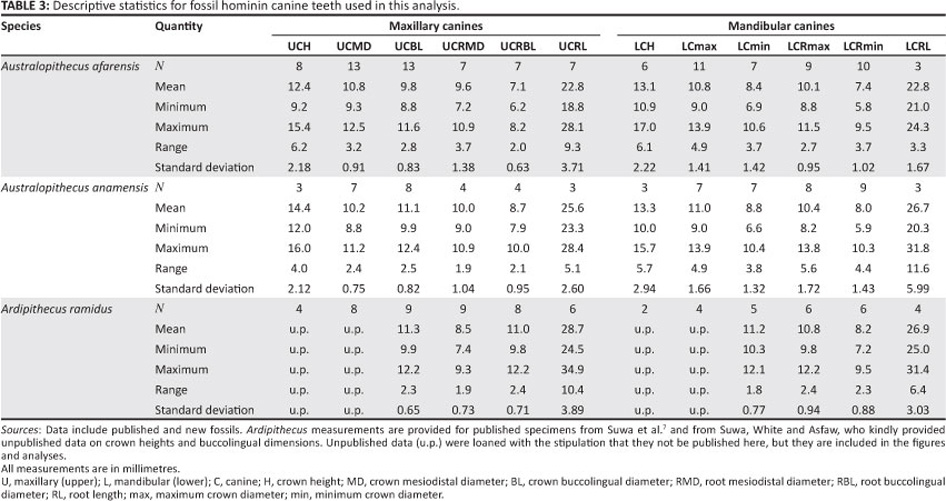
The newly discovered A. anamensis teeth were compared with those of extant great apes (Gorilla gorilla n = 25, Pongo pygmaeus n = 7, Pan troglodytes n = 15, Pan paniscus n = 17 and Homo sapiens n = 25) (Table 2), as well as A. afarensis (n=11),previously described A. anamensis (n=9) and Ar. ramidus (n = 6) (Table 3).
Linear data from fossils were taken on original Kenyan A. anamensis specimens by one of us (C.V.W.). A. afarensis data were kindly provided by William Kimbel and checked against measurements from casts taken by CVW to ensure consistency amongst data sets. Data for the Fejej fossils were taken by C.V.W. from casts kindly provided by John Fleagle and checked against the originals by C.V.W. Ar. ramidus data were taken from Suwa et al.7 with supplementary data on crown heights kindly provided by Gen Suwa, Tim White and Berhane Asfaw (2009, personal communication, December 1), with the stipulation that the unpublished numbers are not for reproduction. Data for the Asa Issie A. anamensis specimens were taken from White et al.9 and checked against the originals by C.V.W.
Maxillary canine basal dimensions are measured as 'mesiodistal' and 'buccolingual'. Morphologically, the maximum diameter of the canine embraces, or nearly embraces, the base of the mesial and distal crests of the tooth in most non-human primates. Because the human tooth is mesiodistally compressed relative to its buccolingual diameter, the maximum diameter of the tooth is not homologous to its mesiodistal dimensions, as in other species. However, mandibular canine basal dimensions are presented using 'maximum' and 'minimum' (being the greatest dimension perpendicular to the maximum) diameters, because, in hominins, the relative position of the tubercles are not located at the mesial and distal margins of the tooth. The same definitions apply to the mandibular canine measurements for non-human primates. Similarly, because the P3 is normally oriented obliquely relative to the tooth row, basal dimensions of the P3 are also measured as maximum and minimum in all human and non-human primates.
We measured all available crown heights for A. anamensis and A. afarensis (Table 4). For many primates, wear is a normal and necessary part of canine function, and in some species the apex of the tooth is worn before the tooth is finished erupting. Canine crown height data from Plavcan23 for 89 extant primates demonstrate that 'moderately worn' (teeth showing some blunting of the apex) and unworn canines do not significantly differ in crown height.24 Given that the criteria used for excluding worn canines for this study were more stringent than for the Plavcan23 data set, apical wear had no significant impact on our results. Here, we did not correct for wear, but have noted those teeth that clearly show apical blunting. Whilst including worn specimens slightly depresses the mean canine height for the hominin sample, and increases the variance, the overall change in variation and the range of crown height is small by comparison to interspecific differences in canine size. Even adding several millimeters to canine dimensions for worn teeth will not affect the results of this study. Given that the canine crowns of A. anamensis and A. afarensis were measured by us in the same way, the interspecific differences in canine size are robust, and our conclusions would not be altered by attempting to estimate the unworn size of the canine teeth.
Canine data for Ar. ramidus were, as reported to us, 'corrected' for wear and damage. Having not studied the original specimens, we cannot quantify whether the measurements are exactly comparable to ours or not. Nevertheless, restricting crown height comparisons to only unworn teeth does not alter any of our results and conclusions. Here we report only the results for the entire sample. Therefore, conclusions drawn from comparisons between the Ardipithecus and Australopithecus canine crown heights appear to be robust.
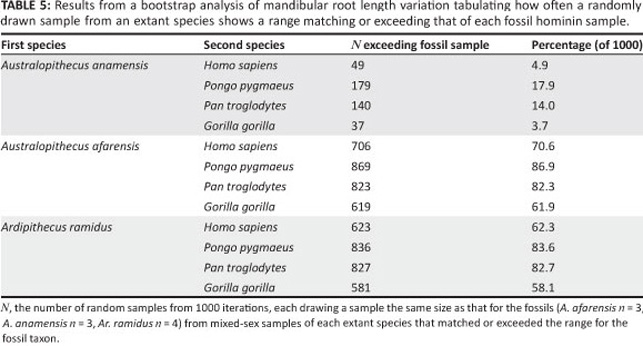
Standard parametric and non-parametric statistical tests were used for most comparisons, as noted where appropriate. To compare the range of variation in root length between hominins and extant apes and Homo, a bootstrap analysis was carried out using a program written in Matlab.25 For each fossil taxon comparison, 1000 random samples from each extant taxon were selected with replacement, and without regard to sex, selecting the same number of specimens as available for the fossil sample. The number of samples with a range equal to or exceeding that of the fossil sample was tabulated.
To evaluate basal canine shape proportions, we used the SMATR26 software package to test for differences in both slope and elevation of reduced major axis lines fit through ln-transformed mesiodistal and buccolingual canine tooth dimensions amongst all extant ape species, A. anamensis and A. afarensis, using 1000 iterations (following Wharton et al.27). To confirm these results, we also performed a least-squares regression through ln-transformed canine mesiodistal versus buccolingual dimensions of apes only. We calculated the analysis of variance of residuals for extant hominoids, A. anamensis and A. afarensis derived from this least-squares regression, using Tukey's honestly significantly different two-tailed tests for post-hoc contrasts between groups at an alpha level of 0.05. In no case did the results differ from the SMATR results, so only results from the latter analysis are reported here as they are statistically most appropriate.27
Results
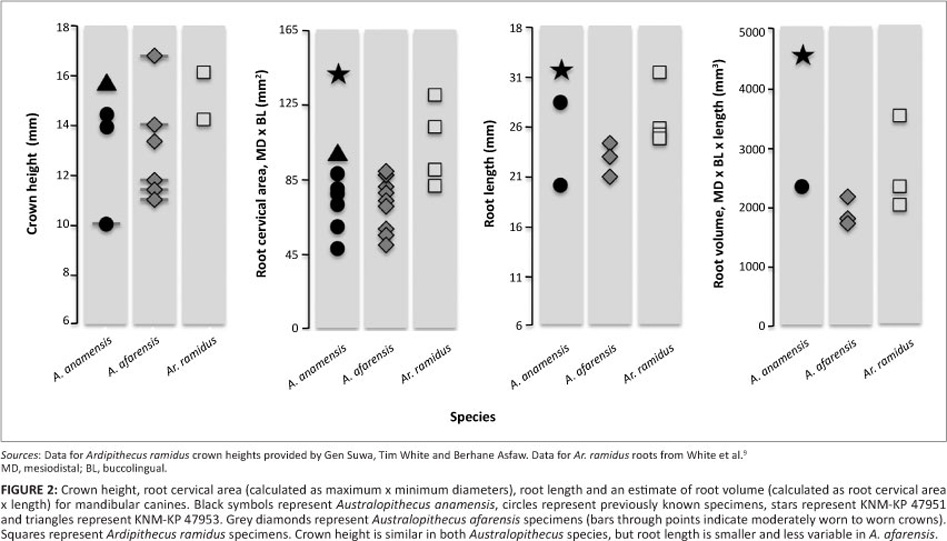
KNM-KP 47951 has a strikingly large and robust mandibular canine root that is the largest known for any early hominin, in length, cervical dimensions and volume (Figure 2). The KNM-KP 47951 canine root is substantially larger than the alveolus for KNM-KP 29287, which was sufficiently large to suggest greater canine size and variation in A. anamensis compared with all later hominins.17,18 KNM-KP 47951 demonstrates that neither KNM-KP 29287 nor FJ-4-SB-1a have unusually large canines, nor would KNM-KP 29287 even have belonged to a particularly large male individual. The large root of KNM-KP 47951 increases the observed range of variation in length and occlusal dimensions in A. anamensis canine teeth, and so also in overall size (Figure 2). It is far greater in length and size than any A. afarensis specimen. Long, large mandibular canine roots also are seen in Ar. ramidus (9) and extant great apes, suggesting that this is a primitive trait for the hominin clade (Figure 2).
Even though sample sizes of complete root lengths are small, A. anamensis has greater variation than minimally dimorphic Homo (Figure 2; Table 5). A. afarensis and Ar. ramidus root length variation, by contrast, is minimal, although too few specimens are preserved with which to assess degrees of variation in either species. Mandibular root size in both A. anamensis and Ar. ramidus is similar, and both are substantially greater than A. afarensis. There is no overlap in mandibular root volume between A. anamensis and A. afarensis (Figure 2). In combination, the data suggest a decline in mandibular root size from the primitive size in A. anamensis to a derived condition in A. afarensis.
The canine crown of the other new mandibular dentition, KNM-KP 47953, supports the observation that although canine crowns were not absolutely taller or broader in A. anamensis than in A. afarensis (11, 16, 17, 18), A. anamensis canines have larger basal dimensions relative to the size of their postcanine teeth (Figure 3). This shift in relative size ratios apparently continues a general trend seen when comparing Ar. ramidus to Australopithecus, and at least partly reflects increasing postcanine tooth size.7,9,13
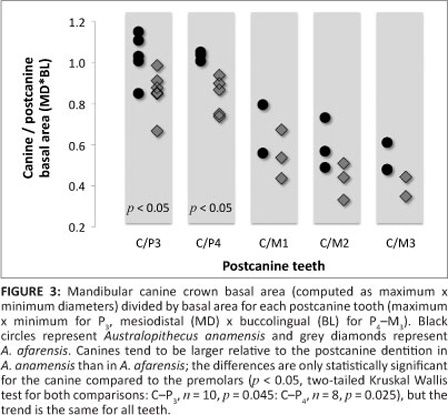
Together, KNM-KP 47951 and KNM-KP 47953, along with previously known specimens, suggest that mandibular canine crown height, breadth and root size variation were not coupled in early Australopithecus. Specifically, the new specimens suggest that whilst canine crowns appear to have reduced in height and dimorphism prior to the appearance of the genus Australopithecus,5,7 root size and variation decreased within the A. anamensis―afarensis lineage independently of crown dimensions. Thus, the apparently large alveolus of KNM-KP 29287 reflects the relatively large roots in the earlier species, and not greater canine crown size as previously hypothesised.17,18
Preserved A. anamensis and A. afarensis fossils do not differ in relative maxillary canine basal crown and root size, but all four A. afarensis specimens are small, suggesting that they may be female individuals, thereby obscuring comparisons. Maxillary canine crown shape does differ as part of an overall shift in morphology of the C-P3 complex during the evolution of A. anamensis into A. afarensis.5,7,8,9,11,19 KNM-KP 47952 demonstrates the previously documented A. anamensis condition of having mesiodistally longer maxillary crowns and roots than does A. afarensis (Figure 4a; Table 6) (see also Leakey et al.15 and Ward et al.18). Notably, A. anamensis is nearly identical to Ar. ramidus in the absolute size and occlusal proportions of the maxillary canines, but both differ from A. afarensis, which has a shape equivalent to that of the more diminutive human canines (Figure 4a; Tables 2 and 3). Thus, A. anamensis retained the primitive condition, and shape change occurred during the evolution of A. afarensis.
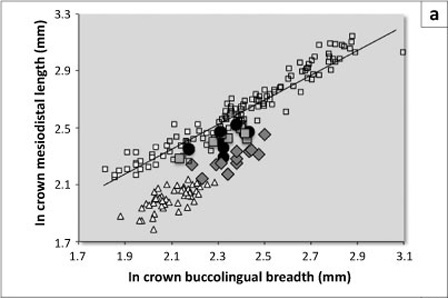

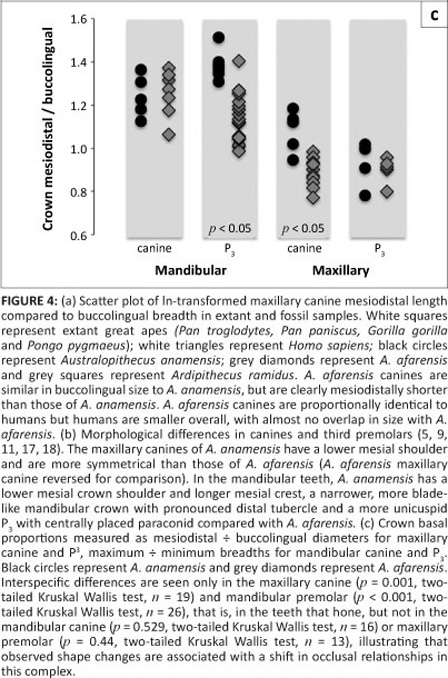
Accompanying this shift in maxillary canine crown proportions, mesial crest length reduced as a function of the mesial shoulder of the tooth shifting apically5,7,9,11 (Figure 4b). This change mirrors the broadening of the mandibular canine, which also experienced morphological alterations through time, becoming less blade-like. Additionally, the mandibular premolar transformed, with the protoconid shifting buccally, affecting the fovea form, and the metaconid expanded in size.11,17,19
Metric changes in basal shape from A. anamensis to A. afarensis occurred in the maxillary canine and mandibular premolar, the honing pair, but not in the mandibular canine or maxillary premolar (Figure 4c). This shape change reflects change in C-P3 function, increasing transverse contact area between maxillary and mandibular teeth, most logically due to increased use of the canine in food acquisition or preparation. This shape change suggests that any associated change in function occurred between A. anamensis and A. afarensis, and not with the origin of Australopithecus. Also, it is now clear that shape changes in the canine-premolar complex did not accompany selection for reduced canine crown height, which was already diminished in earlier hominins (Ardipithecus, Orrorin and Sahelanthropus).2,4,6,9
Discussion
Canine crown reduction is one of the hallmarks of hominin evolution and so plays an important role in identifying potential adaptive changes at the origins of the clade. Multiple hypotheses have been put forward to explain canine reduction amongst hominins in general, including the loss of canine teeth as weapons, dental crowding and selection altering the canines for food ingestion and/or processing.13,22,28,29,30,31,32,33 Hypotheses concerning the co-option of the canine for food processing or gathering either implicitly or explicitly link canine reduction directly to selection for a change in dietary function. Furthermore, whilst large canine roots have been noted in early hominins, the relationship between root and crown reduction (and dimorphism) has not been evaluated.
These new A. anamensis fossils help demonstrate that whilst canine tooth size reduction probably occurred basally in hominin evolution prior to the evolution of Australopithecus, changes in canine shape, in both crowns and roots, occurred in a mosaic fashion throughout the A. anamensis-afarensis lineage. Whatever selective pressure led to canine tooth crown size reduction in human evolution did not occur at the same time as that leading to tooth crown shape change. This finding suggests, in turn, that multiple independent factors altered the complex over time. These phenomena, therefore, appear to have been a result of different pressures. These pressures, in turn, suggest at least the possibility that the canines of australopithecines may have served a different function from those of either their ancestors or their descendants.
Canine crown size and dimorphism were already reduced in all earlier hominins (Ardipithecus, Sahelanthropus and Orrorin) prior to the appearance of Australopithecus,7,8 suggesting that the ancestor of Australopithecus probably had reduced crown size and dimorphism as well. However, substantial shape change did not accompany this crown height reduction. This observation stands in contrast to the hypothesis that shape changed in association with crown height reduction and incorporation of the tooth into an incisal functional field.13 The short canine crowns imply that canines no longer played a role as weapons for intrasexual or intraspecific aggression early in hominin evolution. It follows that changes in canine shape almost certainly do not signal changes in social behaviour in later hominins.
Therefore, further alterations in canine shape within early Australopithecus by default probably reflect changes in food processing. Shorter canine crowns also did not accompany a shift towards thicker tooth enamel and enhanced mastication with the origins of Australopithecus. Rather, canine crown reduction in earlier hominins likely exapted the canines to serve a unique, derived function in Australopithecus, probably in food acquisition and/or processing. The development of the mesial cristid, which contacts the lateral maxillary incisor, and the elevation of the shoulders of the maxillary canine accompanies the shift in canine occlusal shape in Australopithecus, strongly suggesting a dietary function of the canines.13 However, the lack of simultaneous canine size reduction suggests that this change did not reflect a gradual integration of the canine into an integrated anterior incisal mechanism. Rather, it suggests a dietary function unique to Australopithecus, and not simply human-like. Unfortunately, little is known about anterior tooth use and function in early hominins. Recent work suggesting similar overall diets in A. anamensis and A. afarensis is based on molar morphology and microwear.34 These data demonstrate that the diets of both species involved heavy mastication of tough food items with similar material properties, but do not address possible variation in incision or ingestion behaviours, nor do they provide evidence of canine use, which may have differed. To date, evidence of canine use in early hominin canines, such as with microwear, has not been evaluated.
The addition of the new fossils of A. anamensis presented here reveals a dissociation of canine root morphology that appears to have accompanied morphological shifts in the C-P3 complex, but not canine crown height reduction. Canine tooth root size likely accounts, at least in part, for the inflated anterolateral margins of the mandible seen in A. anamensis as compared with A. afarensis, in which the canines are set directly anterior to the postcanine tooth rows in A. anamensis, but more medially in A. afarensis. Relatively large canine roots may also contribute to the inflated canine jugal area and rounded lateral nasal aperture seen in A. anamensis and possibly the earliest A. afarensis (Garusi 1).17 These changes in facial and mandibular form, which may in turn affect masticatory biomechanics, may be spatially linked to the reduction in canine root size and dimorphism. Unfortunately, little is known about the functional significance of variation in canine root size or morphology. At the least, these results suggest that crown and root size are not tightly integrated functionally.
The new Kanapoi fossils underscore the complex, mosaic nature of evolution in the hominin canine honing complex during early hominin evolution, and highlight new questions about hominin dentognathic adaptations. Identifying the order and timing of these morphological and proportional changes provides a basis for developing accurate hypotheses to explain the selective factors acting on crown height, crown shape and root dimensions in Australopithecus that will play a key role in understanding the adaptive transition from A. anamensis to A. afarensis.
Acknowledgements
We thank the curators and staff of the National Museums of Kenya, National Museum of Ethiopia, Turkana Basin Institute, Cleveland Museum of Natural History, National Museum of Natural History and Royal Museum of Central Africa for access to collections in their care and assistance. We would also like to thank Gen Suwa, Tim White, Berhane Asfaw and William Kimbel for data and access to original fossils; John Fleagle and William Kimbel for casts; and William Kimbel, Milford Wolpoff, Alan Walker, Bernard Wood, Luke Delazene, Yohannes Haile-Selassie, Scott Simpson, John Fleagle and anonymous reviewers for helpful comments and discussion. This project was supported by the Leakey Foundation, PAST (South Africa), Turkana Basin Institute and the Wenner Gren Foundation for Anthropological Research.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
F.K.M. led the team that recovered the fossils; J.M.P. conducted the statistical analyses; C.V.W. described the fossils; and all authors contributed to writing the manuscript.
References
1. Dart RA. Australopithecus africanus: The man-ape of South Africa. Nature. 1925;115:195. http://dx.doi.org/10.1038/115195a0 [ Links ]
2. Brunet M, Guy F, Pilbeam D, et al. A new hominid from the Upper Miocene of Chad, Central Africa. Nature. 2002;418:145-151. http://dx.doi.org/10.1038/nature00879, PMid:12110880 [ Links ]
3. Senut B, Pickford M, Gommery D, Mein P, Cheboi K, Coppens Y. First hominid from the Miocene (Lukeino Formation, Kenya). C R Acad Sc Paris. 2001;332:137-144. [ Links ]
4. Haile-Selassie Y. Late Miocene hominids from the Middle Awash, Ethiopia. Nature. 2001;412:187-191. [ Links ]
5. Haile-Selassie Y, Suwa G, White TD. Late Miocene teeth from middle Awash, Ethiopia, and early hominid dental evolution. Science. 2004;303:1503-1505. http://dx.doi.org/10.1126/science.1092978, PMid:15001775 [ Links ]
6. Haile-Selassie Y, WoldeGabriel G, editors. Ardipithecus kedabba: Late Miocene evidence from the Middle Awash, Ethiopia. Berkeley: University of California Press; 2009. [ Links ]
7. Suwa G, Kono R, Simpson S, Asfaw B, Lovejoy C, White T. Paleobiological implications of the Ardipithecus ramidus dentition. Science. 2009;236:94-99. [ Links ]
8. White T, Asfaw B, Beyene Y, et al. Ardipithecus ramidus and the paleobiology of early hominids. Science. 2009;326:64-86. http://dx.doi.org/10.1126/science.1175802 [ Links ]
9. White T, WoldeGabriel G, Asfaw B, et al. Assa Issie, Aramis and the origin of Australopithecus. Nature. 2006;440:883-889. http://dx.doi.org/10.1038/nature04629, PMid:16612373 [ Links ]
10. White TD, Suwa G, Asfaw B. Australopithecus ramidis, a new species of early hominid from Aramis, Ethiopia. Nature. 1994;371:306-312. http://dx.doi.org/10.1038/371306a0, PMid:8090200 [ Links ]
11. Kimbel W, Lockwood C, Ward CV, Leakey M, Rak Y, Johanson D. Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. J Hum Evol. 2006;51:134-152. http://dx.doi.org/10.1016/j.jhevol.2006.02.003, PMid:16630646 [ Links ]
12. White TD. Earliest hominids. In: Hartwig WC, editor. The primate fossil record. Cambridge: Cambridge University Press, 2002; p. 407-417. [ Links ]
13. Greenfield L. Origins of the human canine; a new solution to an old enigma. Yearb Phys Anthropol. 1992;35:153-185. http://dx.doi.org/10.1002/ajpa.1330350607 [ Links ]
14. Plavcan JM. Inferring social behavior from sexual dimorphism in the fossil record. J Hum Evol. 2000;39:327-344. http://dx.doi.org/10.1006/jhev.2000.0423, PMid:10964532 [ Links ]
15. Leakey MG, Feibel CS, MacDougall I, Ward CV, Walker A. New specimens and confirmation of an early age for Australopithecus anamensis. Nature. 1998;363:62-66. http://dx.doi.org/10.1038/29972, PMid:9590689 [ Links ]
16. Leakey MG, Feibel CS, McDougall I, Walker A. New four-million-year-old hominid species from Kanapoi and Alia Bay, Kenya. Nature. 1995;376:565-571. http://dx.doi.org/10.1038/376565a0, PMid:7637803 [ Links ]
17. Ward CV, Leakey MG, Walker A. Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya. J Hum Evol. 2001;41:255-368. http://dx.doi.org/10.1006/jhev.2001.0507, PMid:11599925 [ Links ]
18. Ward CV, Walker A, Leakey MG. The new hominid species Australopithecus anamensis. Evol Anthropol. 1999;7:197-205. http://dx.doi.org/10.1002/(SICI)1520-6505(1999)7:6<197::AID-EVAN4>3.0.CO;2-T [ Links ]
19. Haile-Selassie Y, Saylor B, Deino A, Alene M, Latimer B. New hominid fossils from Woranso-Mille (Central Afar, Ethiopia) and taxonomy of early Australopithecus. Am J Phys Anthropol. 2010;141:406-417. PMid:19918995 [ Links ]
20. Fleagle JG, Rasmussen DT, Yirga S, Bown TM, Grine FE. New hominid fossils from Fejej, southern Ethiopia. J Hum Evol. 1991;21:145-152. http://dx.doi.org/10.1016/0047-2484(91)90005-G [ Links ]
21. McDougall I, Brown F. Geochronology of the pre-KBS Tuff sequence, Omo Group, Turkana Basin. J Geol Soc. 2008;165:549-562. http://dx.doi. org/10.1144/0016-76492006-170 [ Links ]
22. Wolpoff M. Paleoanthropology. New York: Knopf; 1980. [ Links ]
23. Plavcan J. Sexual dimorphism in the dentition of extant anthropoid primates. Durham: Duke University; 1990. [ Links ]
24. Plavcan J, Ward CV, Paulus F. Estimating tooth crown height in early Australopithecus. J Hum Evol. 2009;57:2-10. http://dx.doi.org/10.1016/j.jhevol.2009.04.005, PMid:19482334 [ Links ]
25. Matlab. Version 7. Natick, MA: Mathworks; 2010. [ Links ]
26. SMATR (Standardized Major Axis Tests and Routines). Version 2. Sydney: Falster D, Wharton D, Wright I; 2006. [ Links ]
27. Wharton D, Wright I, Falster D, Westoby M. Bivariate line-fitting methods for allometry. Biol Rev. 2006;81:259-291. http://dx.doi.org/10.1017/S1464793106007007, PMid:16573844 [ Links ]
28. Bailit H, Friedlaender J. Tooth size reduction: A hominid trend. Am Anthropol. 1966;68:665-72. http://dx.doi.org/10.1525/aa.1966.68.3.02a00030 [ Links ]
29. Brace C. Structural reduction in evolution. Am Nat. 1963;97:39-49. [ Links ]
30. Calcagno J, Gibson K. Human dental reduction: Natural selection or probable mutation effect? Am J Phys Anthropol. 1988;77:505-517. http://dx.doi.org/10.1002/ajpa.1330770411, PMid:3223518 [ Links ]
31. Wolpoff M. The effect of mutations under conditions of reduced selection. Soc Biol. 1969;16:11-23. PMid:5803566 [ Links ]
32. Jungers W. On canine reduction in early hominids. Curr Anthropol. 1978(19):155-156. http://dx.doi.org/10.1086/202027 [ Links ]
33. Szalay FS. Hunting-scavenging protohominds: A model for hominid origins. Man. 1975;10:420-429. http://dx.doi.org/10.2307/2799811 [ Links ]
34. Ungar P, Scott R, Grine F, Teaford M. Molar microwear textures and the diets of Australopithecus anamensis and Australopithecus afarensis. Philos Trans R Soc London. 2010;365:3345-3354. http://dx.doi.org/10.1098/rstb.2010.0033, PMid:20855308 [ Links ]
 Correspondence to:
Correspondence to:
Carol Ward
Postal address: Department of Pathology and Anatomical Sciences
M263 Medical Sciences Building
University of Missouri
Columbia, MO 65212, USA
Email: wardcv@missouri.edu
Received: 25 Apr. 2011
Accepted: 15 Sept. 2011
Published: 02 Mar. 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}