










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.108 no.1-2 Pretoria Jan. 2012
RESEARCH ARTICLE
The first animals: ca. 760-million-year-old sponge-like fossils from Namibia
C.K. 'Bob' BrainI; Anthony R. PraveII; Karl-Heinz HoffmannIII; Anthony E. FallickIV; Andre BothaV; Donald A. HerdII; Craig SturrockVI; Iain YoungVII; Daniel J. CondonVIII; Stuart G. AllisonII
IDitsong Museum, Northern Flagship Institution, Pretoria, South Africa
IIDepartment of Earth Sciences, University of St Andrews, St Andrews, UK
IIIGeological Survey, Ministry of Mines and Energy, Windhoek, Namibia
IVScottish Universities Environmental Research Centre, East Kilbride, UK
VLaboratory for Microscopy and Microanalysis, University of Pretoria, Pretoria, South Africa
VIEnvironmental Sciences, University of Nottingham, Nottingham, UK
VIISchool of Environmental and Rural Sciences, University of New England, Armidale, New South Wales, Australia
VIIINERC Isotope Geosciences Laboratory, Keyworth, UK
ABSTRACT
One of the most profound events in biospheric evolution was the emergence of animals, which is thought to have occurred some 600-650 Ma. Here we report on the discovery of phosphatised body fossils that we interpret as ancient sponge-like fossils and term them Otavia antiqua gen. et sp. nov. The fossils are found in Namibia in rocks that range in age between about 760 Ma and 550 Ma. This age places the advent of animals some 100 to 150 million years earlier than proposed, and prior to the extreme climatic changes and postulated stepwise increases in oxygen levels of Ediacaran time. These findings support the predictions based on genetic sequencing and inferences drawn from biomarkers that the first animals were sponges. Further, the deposition and burial of Otavia as sedimentary particles may have driven the large positive C-isotopic excursions and increases in oxygen levels that have been inferred for Neoproterozoic time.
Introduction
Evidence for the early evolution of animals has been documented from three main fossil assemblages of Ediacaran age (635-542 Ma). The first assemblage consists of phosphatised embryos, recovered principally from sedimentary successions in South China.1,2,3 The second is defined by large, complex fossils known collectively as the Ediacaran, of worldwide occurrence, which arose and largely went extinct through the last third of that geological period.4,5 The last is a group of small biomineralisers that first evolved about 550 Ma.6,7 These fossil discoveries reveal a relatively rich assemblage of organisms preserved in strata that predate by several tens of millions of years the base of the Cambrian at 542 Ma.8,9,10 Although older trace fossils have been assigned an animal origin,11,12 no undisputed body fossils of animals are known from rocks older than about 650 Ma. Nevertheless, molecular clock estimates place the origin of metazoans much deeper in time and predict that a major split in animal evolution occurred some 100 to 300 million years prior to the Cambrian explosion13,14,15 - an event that is thought to define the advent of sponges and subsequent divergence of all other animals. An exciting discovery in Australia in rocks 650-635 million years old of inferred animal-body fossils thought to be sponges16 supports the contention that Porifera may form the oldest group of animal lineages.
Ediacaran-age sponge embryos and/or siliceous spicules have been documented in the Doushantuo Formation in south China,17 Tsagaan Gol Formation in south-western Mongolia18 and the Ediacara fauna of south Australia (Palaeophragmodictya).19 These fossils are interpreted as either Demospongiae or Hexactinellida. In the Nama Group from southern Namibia, Namapoikia rietoogensis, a biomineralised metazoan that grew to as much as 1 m in diameter, is thought to have had a cnidarian or poriferan affinity.20 Molecular biomarkers of marine demosponges that are somewhat older than 635 Ma have been found in rocks of the Huqf Supergroup in south Oman.21 The forms reported herein and described below are interpreted as animal fossils but are much older than any yet reported: time constraints indicate that the organism evolved at least 760 Ma in the Cryogenian.
The marine carbonate rocks that formed in the Neoproterozoic oceans in which these organisms lived are marked by C-isotopic compositions much more enriched in 13C than found in the previous billion years, or in the subsequent half billion years of the Phanerozoic; these long-lived high d13C plateaux were themselves punctuated by shorter-lived, large-amplitude excursions.22 Although some workers have concluded that such patterns are as a result of diagenetic effects,23,24 the systematic nature and coherence of C-isotopic trends from numerous sections worldwide is not easily accommodated in such a model. We offer an additional and alternative idea: that the evolution of the organisms described herein, with an exterior shell, initiated a global-scale burial of organic matter that was encased in hard parts. This sedimentation would have removed isotopically light organic matter from the oceans, thereby contributing to enrichment of marine carbonates in 13C; such a perturbation of the global C cycle could also have influenced oxygen budgets.
Methods
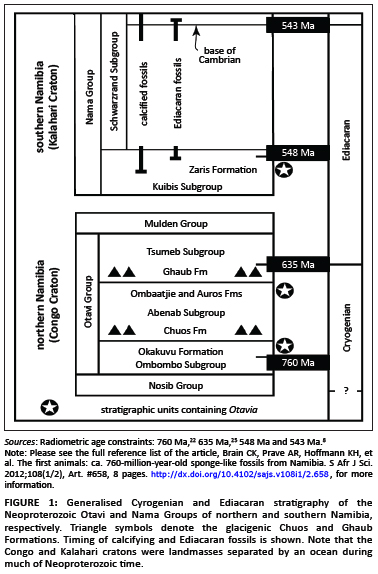
Individual hand-sized samples of black limestones from the Otavi and Nama Groups in Namibia (Figure 1) were collected and dissolved in acetic acid. More than 800 thin sections of rock were examined and over 1000 Otavia specimens were hand-picked from the acid residues.25 Representative Otavia were picked for analyses using scanning electron microscopy (SEM), electron probe imaging (BSE), cathode luminescence microscopy (CL) and X-ray microtomography (XMT) (see Box 1 for details). In addition, 153 rock samples spanning the lithologies and stratigraphy in which Otavia fossils have been found were analysed for carbonate C-O stable isotope ratios at the Scottish Universities Environmental Research Centre; those data will be reported in a separate article.
Results
Prior to a detailed discussion of the inferred fossil, we pose and answer two fundamental methodological questions related to our underlying assumptions:
1. Is there any evidence that the ensemble of objects (i.e. the inferred microfossils) is derived from one or more genetically related populations?
2. Is there a priori quantitative evidence against biogenicity of the objects?
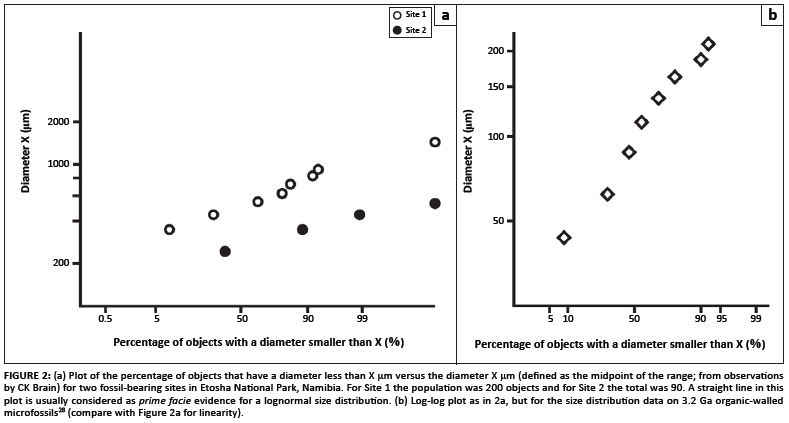
The measured size distribution of the objects recovered at two of the richest sampling sites (both in Etosha National Park, Namibia) provides an answer to the first question. The null hypothesis is that the objects are not genetically related, and so no internally consistent size distribution is expected at either site. In Figure 2a the diameter X (defined as the midpoint of the observed range of sizes) is plotted against the percentage of the observed population with diameter less than X. A linear relationship in this log-probability plot is usually taken as prima facie evidence of a log-normal distribution of sample sizes.26 Data from each site are, to first order, impressively linear and so the null hypothesis is rejected. This result deals satisfactorily with the issue of the ensembles of observed objects being part of a genetically related population at each locality. We remain uncertain as to why there are size differences, but it likely reflects an environmental control in that the larger specimens are recovered from a locality that represents relatively deeper, quieter conditions. Work is continuing on this issue.
We also stress that this conclusion provides no evidence of biogenicity: for example, Fallick et al.27 demonstrated convincing evidence for a truncated lognormal distribution of sizes for a suite of iron droplets in lunar soil 10084. Given that our positive size-distribution analysis cannot be taken as prima facie evidence of biogenicity, we then address the second of the above questions and ask: is a lognormal distribution of object sizes consistent with biogenicity?
That the answer to this question is positive is proved by data derived from Javaux et al.28 and plotted in Figure 2b. Those workers described specimens that are believed to be 3.2-billion-year-old organic-walled microfossils, and are thus accepted as biogenic. The scatter about the line in the log- probability plot is not distinctly different from the scatter associated with the objects we propose herein as biogenic fossils. Hence, our fundamental questioning of core assumptions has been satisfactorily dealt with. What follows is a detailed description of the putative fossils themselves.
Otavia fossils: Stratigraphy
We term the putative fossils Otavia, because they have been recovered most abundantly from black limestone units of the Otavi Group of Namibia (Figure 1). Otavia have also been found in the Nama Group of Namibia. Both Otavi and Nama Groups are several kilometres thick and comprised of siliciclastic and carbonate rocks deposited in shallow marine shelf and slope environments. High precision U-Pb ages on zircons from syndepositional volcanic ash beds provide geochronological constraints on these rocks. The oldest Otavia to have been discovered come from the Otavi Group, in units correlative with upper Ombombo Subgroup strata (Okakuyu Formation) that occur above an ash bed dated at 760±1 Ma and which lies beneath the glacigenic Chuos Formation of ca. 710 Ma.22 Otavia were also found in Otavi Group rocks (Ombaatjie and Auros Formations) underlying the glacigenic Ghaub Formation, the meltback phase of which is dated at 635.5±1.2 Ma.29 The stratigraphically youngest known Otavia come from rocks in the Nama Group (Zaris Formation, Mara Member) that are correlative with units beneath an ash bed dated at 548.8±1 Ma.8
Otavia fossils: Description
The general shape of Otavia is elongate ovoid to globular and, although individuals vary, the overall form of each remains similar (Figure 3). Sizes range from as small as 0.3 mm to as large as 5 mm in the longest dimension. The outer surface of Otavia is perforated and pierced by numerous small holes, typically 5 mm - 20 mm across, as well as a number of larger openings that form raised turrets or mounds several to many tens of microns in diameter (Figure 3).

The internal character of Otavia provides evidence for its biological origin. SEM, BSE, CL and XMT images show that Otavia consists of three components ( Figures 4 and 5): (1) an exterior wall that is several to many microns thick, pierced by numerous holes and comprised mostly of calcium phosphate and minor dolomite (secondarily silicified Otavia have been found in Nama Group limestones25); (2) a peripheral labyrinth (so termed because it encloses a large inner cavity) a few to many tens of microns in thickness, and composed of calcium phosphate (minor areas of calcite and replacement dolomite are also present); and (3) an irregularly shaped inner cavity that runs much of the length of Otavia and which is now filled with fine-grained carbonate sediment. In effect, Otavia resembles an elongate, irregularly shaped, hollow container(Figures 4 and 5).
The larger openings, which form raised turrets on the outer walls of Otavia, are linked to passageways that transect the peripheral labyrinth and open directly into the internal cavity; these openings provide a direct communication between the cavity and the outside ( Figure 4a, i, j; Figure 5a, b). The smaller holes or pores that pierce the exterior wall open into the peripheral labyrinth (Figure 4e; Figure 5c, d). The labyrinth consists of layered galleries of interlinked chambers bounded by several-micron-thick tabular to wavyparallel mineralised (mostly phosphatised) walls (Figure 4c, d, e, f; Figure 5d). The inner portion of the labyrinth adjacent to the internal cavity displays wavy to cuspate surfaces with many openings that link the chambers of the labyrinth to the internal cavity (Figure 4c, d, e, f; Figure 5b, d). The internal cavity can comprise lesser or greater volumes of the interior of Otavia and XMT reconstructions show that it can be complex in shape (XMT software can be programmed to eliminate solid material and highlight void spaces), reflecting the irregular form of the inner bounding surfaces of the peripheral labyrinth ( Figure 5e, f).
The mechanism of phosphate concentration and fixation remains to be ascertained. Phosphate preferentially occurs in the interior portions of Otavia, implying a genesis linked to conditions internal to Otavia and not as a result of an externally precipitated, microbially mediated layering, coating or binding. Parts of the walls and labyrinth are calcite and it is assumed that Otavia was originally composed of calcium carbonate and, to a lesser extent, dolomite.
Regardless of what original mineralogy Otavia used to construct itself, its overall structure indicates that it developed the capability to control mineral precipitation. The walls and peripheral labyrinth reveal sharp boundaries between internal and external areas and their surfaces show repeated patterns and complex shapes (Figure 4c, d). That Otavia was a hollow container is evident from the fact that its internal cavity can be filled with sediment different from the encompassing sedimentary matrix (i.e. a geopetal sediment) ( Figure 4f, g, i, j). In some Otavia, especially near to large external openings, the sediment within the interior cavity is mottled, having characteristics of both the internal (geopetal) and external sediment. This finding indicates that both sediment types were able to mix, additionally confirming that Otavia was hollow during life. In these cases, the cavities of Otavia were filled after death and then transported as a bioclast (fragments of Otavia can be seen in Figure 4i, j). In other cases, no distinction can be made between the internal and external sediment (Figure 4f), implying that those Otavia remained largely in situ after death and burial.
Otavia fossils: Interpretation
In describing the features that characterise Otavia, it is also important to stress what features have not been observed. To do so is important because microbial sedimentary structures such as thrombolites (mottled textures) and stromatolites (layered textures) are common in the Otavi and Nama Group rocks and it is necessary to reduce, and perhaps eliminate, the possibility that Otavia are bacterially mediated and/or influenced structures.
In the hundreds of Otavia specimens examined, not a single example of micron-scale globular clusters, botryoidal surfaces, or even singular ovoid (coccoid) forms was observed which would suggest an origin by bacteria or bacterial communities. Otavia do not display micron-scale layering and thus cannot be attributed to microbial sheets, veneers, coatings, layered cortices around clasts and grains, or to crystal shapes. Furthermore, the cavities and layered chambered labyrinths of Otavia are typically tens of microns in size and thus are not the micron-scale tubules of cyanobacteria. Nor is Otavia an aggregate or cluster of detrital grains and allochems (secondarily reworked and transported carbonate or other types of grains) bounded by microbial sheets (i.e. microphytollites). Irregularly shaped filamentous sheaths that trap and bind particles are readily distinguishable from Otavia (Figure 4j).
We have also considered the possibility that the Otavia organisms could have been foraminifera. However, documented examples of Cryogenian-age foraminifera (Amoebozoa) display agglutinated tests comprised of abundant Al-K-Mg silicate minerals embedded in carbonaceous matter.30 Otavia, in contrast, has a phosphatic shell. Further, the foraminifera are typically smaller (generally < 200 mm30) than Otavia and do not display the variably sized wall-piercing openings and passageways that seem to be diagnostic of Otavia. Hence, Neoproterozoic foraminifera are both compositionally and morphologically dissimilar to Otavia, and we thereby discount the possibility that Otavia could be a form of Amoebozoa.
Synthesising all of the above, the most parsimonious interpretation of Otavia is that it was a sponge-like organism. The small openings represent incurrent pores or ostia, the larger ones exhalent oscula, and the internal cavity a paragastric chamber (Figure 6). The limestones containing Otavia vary from grainstones to micrites and exhibit a variety of shallow-water features including oolitic lenses, tidal bundles, cross-stratification, stromatolitic and microbial laminites, intraclastic rip-up beds, hummocky cross-stratified bedforms and wave ripples. Hence, Otavia lived in normal marine conditions.

Otavia occur as unabraded fossils, as well as reworked bioclasts. The unabraded fossils are largely found in finely laminated micritic limestones representing relatively calm depositional conditions free from agitation and implying a setting of quiet, low-energy conditions. The reworked forms are recovered from intraclastic packstones and grainstones, indicating stronger current activity and transport of Otavia as part of the tractive bedload. As such, Otavia's skeleton was resilient enough to survive abrading during transport. Thus, the classification proposed here is that Otavia belong to the phylum Porifera. A prolonged search has been undertaken for spicules, as these would clarify the taxonomic relationship, but no unquestionable example has been found. The etymology of the name Otavia antiqua gen. et sp. nov. is derived from the Otavi Group in which the fossils occur and their age.
Discussion
Sponges require their ostia to be large enough to allow the entry of water drawn inwards by beating flagellae in the peripheral labyrinth and paragastric chambers. Assessing the efficiency at which Otavia would have been able to draw fluid into their bodies is speculative. However, if models are broadly correct in inferring that Neoproterozoic oceans were characterised by large pools of micron-scale particulate and dissolved organic matter,31 then the amount of fluid Otavia needed to filter for nutrition would have certainly been considerably less than that required for modern sponges.
Neoproterozoic time was marked by severe climatic conditions and widely fluctuating marine C-isotopic compositions.22,32 The pattern of C-isotopic values recorded in Neoproterozoic carbonate rocks worldwide has been well documented, with most workers interpreting the positive and negative excursions as globally generated secular variations that are useful in establishing correlations between regions and continents.22 As noted previously, others have concluded that such large-magnitude fluctuations are diagenetic overprints, post-depositional phenomena largely unrelated to original seawater compositions.23 Ascertaining which of these two opposing views is more correct is beyond the scope of this paper. However, it is interesting to consider the effect that the development of mineralogical hard parts enclosing organic matter would have had on the global C cycle. In effect, the genesis of hard parts would have made Otavia behave, particularly upon death, like any other sedimentary particle - it would have undergone transport and burial, thereby removing isotopically light organic C from the global oceanic reservoir.
The amount of biomass Otavia represented is unknown. However, the number of Otavia fragments that can be observed in thin section (e.g. see Figure 4i, j) and recovered from residues is impressive; if this number is representative of Otavia's global distribution, then the likelihood is that Otavia was abundant. The widespread burial of Otavia would have removed organic matter from the water column, driving d13Ccarbonate towards the high values that typify the Neoproterozoic22 and helping generate the inferred rise in Neoproterozoic oxygen levels.33,34 Until the evolution of burrowing metazoans, re-entrainment of sedimented Otavia would have been at the whim of physical processes stirring bottom layers. However, once metazoans evolved, their behavioural activities (such as burrowing and digging) would have reworked the sedimented organic matter back into the water column where it would have oxidised, driving C-isotopic values back towards 'normal'.
Given the existing time and stratigraphic constraints, Otavia predated and lived through the extreme environmental changes of later Neoproterozoic time - the snowball Earth events.32 Otavia evolved prior to the earliest of the documented Neoproterozoic global glaciations, survived through the repeated icehouse-greenhouse states, and existed at least to the dawn of metazoans at the end of the Precambrian. Hence, Otavia could not have been adversely affected by the severe environmental fluctuations and associated changing conditions. It is often postulated that the cessation of the climatic extremes concomitant with an inferred, stepwise global rise in oxygen levels near the close of Proterozoic time33,34 set the stage for the emergence of animals. The presence of Otavia in rocks 760 million years old requires this view to be revised. If our interpretation of Otavia is correct, then the early lineages of animals go back considerably further in time on the basis of molecular evidence and, now, on fossil evidence.
Conclusions
Microfossils, termed Otavia antiqua gen. et sp. nov., have been recovered from Cryogenian-Ediacaran successions in Namibia (the Otavi and Nama Groups) in rocks as old as 760 Ma to as young as 550 Ma. They are interpreted as calcareous sponges and occur as ovoid to irregularly elongate globular forms, ranging in size from 0.3 mm to 5 mm in the longest dimension. The outer surface of Otavia is perforated by numerous small and large openings (several to many tens of microns in size), with the latter commonly forming raised mounds. The outer wall of Otavia is thin (many microns to a few tens of microns thick), typically calcium phosphate in composition, and interior to the wall is a peripheral labyrinth, so named because it encases an irregularly shaped, longitudinally running inner cavity that is connected via irregular pathways through the peripheral labyrinth to the outer-wall openings. Combined, these features are interpreted as the ostia, oscula and paragastric chamber of an ancient calcareous sponge. The evolution of hard parts, effectively encasing organic matter, made Otavia a sedimentary particle. It is speculated that sedimentation and burial of Otavia helped drive the increase in the d13C composition of Neoproterozoic oceans, and perhaps even contributed to the inferred rise in oxygen levels. Otavia evolved prior to the first known Neoproterozoic global (snowball Earth) glaciation, and survived through the climatic extremes associated with the repeated snowball Earth episodes.
Acknowledgements
We thank the Namibian Ministry of Wildlife Management and Tourism, the Namibian Geological Survey, C. Brain, L. Brain, A. Gunzel, R. Newman, D. Newman and E. Voigt for research assistance. Discussions with S. Bengtson, M. Brasier, S. Conway Morris, C. Gaucher, J.G. Gehling, G. Germs, J. Grotzinger, A. Knoll, B. Runnegar, P. Vickers-Rich and R. Wood are much appreciated. C.K.B. thanks the Transvaal Museum and Northern Flagship Institution, the Palaeontology Scientific Trust and the National Research Foundation of South Africa for support. A.R.P. and A.E.F. acknowledge support from NERC, The Carnegie Trust for The Universities of Scotland and The Russell Trust. The proposed Otavia antiqua type specimens, picked from acetic acid residues of Auros Formation limestone from Halali Hill South, Etosha National Park, will be deposited in the collection of the Geological Survey of Namibia in Windhoek, together with thin sections.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
C.K.B. was the project leader. A.P. and K.H. were responsible for fieldwork and A.F., A.B., D.A., C.S. and I.Y. performed the various analyses. C.K.B., A.P., K.H., A.F., D.A., D.C. and S.A. made conceptual contributions. C.K.B., A.P. and A.F. wrote the manuscript.
References
1. Xiao S, Zhang Y, Knoll AH. Three-dimensional preservation of algae and animal embryos in a Neoproterozoic phosphorite. Nature. 1998;391(6667):553–558. http://dx.doi.org/10.1038/35318 [ Links ]
2. Hagadorn JW, Xiao S, Donoghue PCJ, et al. Cellular and subcellular structure of Neoproterozoic animal embryos. Science. 2006;314(5797):291-294. http://dx.doi.org/10.1126/science.1133129, PMid:17038620 [ Links ]
3. Yin L, Zhu M, Knoll AH, Yuan X, Zhang J, Hu J. Doushantuo embryos preserved inside diapause egg cysts. Nature. 2007;446(7136):661-663. http://dx.doi.org/10.1038/nature05682, PMid:17410174 [ Links ]
4. Narbonne GM. The Ediacara biota: Neoproterozoic origin of animals and their ecosystems. Ann Rev Earth Planet Sci. 2005;33:421–442. http://dx.doi.org/10.1146/annurev.earth.33.092203.122519 [ Links ]
5. Jensen S, Gehling JG, Droser ML. Ediacaran-type fossils in Cambrian sediments. Nature. 1998;393(6685):567-569. http://dx.doi.org/10.1038/31215 [ Links ]
6. Grotzinger J, Watters W, Knoll AH. Calcified metazoans in thrombolitestromatolite reefs of the terminal Proterozoic Nama Group, Namibia. Paleobiology. 2000;26(3):334-359. http://dx.doi.org/10.1666/0094-8373(2000)026<0334:cmitsr>2.0.CO;2
7. Amthor JE, Grotzinger JP, Schroeder S, et al. Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman. Geology. 2003;31(5):431-434. http://dx.doi.org/10.1130/0091-7613(2003)031<0431:eocana>2.0.CO;2
8. Grotzinger JP, Bowring SA, Saylor BZ, Kaufman AJ. Biostratigraphic and geochronologic constraints on early animal evolution. Science. 1995;270(5236):598–604. http://dx.doi.org/10.1126/science.270.5236.598 [ Links ]
9. Martin MW, Grazhdankin DV, Bowring SA, Evans DAD, Fedonkin MA, Kirschvink JL. Age of Neoproterozoic bilatarian body and trace fossils, White Sea, Russia: Implications for metazoan evolution. Science. 2000;288(5467):841-845. http://dx.doi.org/10.1126/science.288.5467.841, PMid:10797002 [ Links ]
10. Condon DJ, Maoyan Z, Bowring SA, Wei W, Aihua Y, Yugan J. U-Pb ages from the Neoproterozoic Doushantuo Formation, China. Science. 2005;308(5718):95-98. http://dx.doi.org/10.1126/science.1107765, PMid:15731406 [ Links ]
11. Seilacher S, Bose PK, Pflueger F. Triploblastic animals more than 1 billion years ago: Trace fossil evidence from India. Science. 1998;281(5386);80-83. http://dx.doi.org/10.1126/science.282.5386.80, PMid:9756480 [ Links ]
12. Rasmussen B, Bengston S, Fletcher IR, McNaughton NJ. Discoidal impressions and trace-like fossils more than 1200 million years old. Science. 2002;296(5570);1112-1115. http://dx.doi.org/10.1126/science.1070166, PMid:12004128 [ Links ]
13. Doolittle RF, Feng DF, Tsang S, Cho G, Little E. Determining divergence times of major kingdoms of living organisms with a protein clock. Science. 1996;271(5248);470-477. http://dx.doi.org/10.1126/science.271.5248.470, PMid:8560259 [ Links ]
14. Wray GA, Levinton JS, Shapiro LH. Molecular evidence for deep Precambrian divergence among metazoan phyla. Science. 1996;274(5287):568–573. http://dx.doi.org/10.1126/science.274.5287.568 [ Links ]
15. Peterson KJ, Butterfield NJ. Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record. Proc Natl Acad Sci USA. 2005;102(27):9547–9552. http://dx.doi.org/10.1073/pnas.0503660102, PMid:15983372, PMCid:1172262 [ Links ]
16. Maloof AC, Rose CV, Beach R, et al. Possible animal-body fossils in pre-Marinoan limestones from South Australia. Nat Geosci. 2010;3(9):653-659. http://dx.doi.org/10.1038/ngeo934 [ Links ]
17. Li CW, Chen JY, Hua TE. Precambrian sponges with cellular structures. Science. 1998;279(5352):879–882. http://dx.doi.org/10.1126/science.279.5352.879, PMid:9452391 [ Links ]
18. Brasier MD, Green O, Shields G. Ediacaran sponge spicule clusters from southwestern Mongolia and the origins of the Cambrian fauna. Geology. 1997;25(4):303-306. http://dx.doi.org/10.1130/0091-7613(1997)025<0303:esscfs>2.3.CO;2
19. Gehling JG, Rigby JK. Long expected sponges from the Neoproterozoic Ediacara fauna of South Australia. J Palaeont. 1996;70(2):185-195. [ Links ]
20. Wood RA, Grotzinger JP, Dickson JAD. Proterozoic modular biomineralized metazoan from the Nama Group, Namibia. Science. 2002;296(5577):2383-2386. http://dx.doi.org/10.1126/science.1071599, PMid:12089440 [ Links ]
21. Love GD, Grosjean E, Stalvies C, et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian. Nature. 2009;457(7230):718-723. http://dx.doi.org/10.1038/nature07673, PMid:19194449 [ Links ]
22. Halverson GP, Hoffman PF, Schrag DP, Maloof AC, Rice AHN. Toward a Neoproterozoic composite carbon-isotope record. Geol Soc Amer Bull. 2005;117(9):1181–1207. http://dx.doi.org/10.1130/B25630.1 [ Links ]
23. Knauth LP, Kennedy MJ. The late Precambrian greening of the Earth. Nature. 2009;460(7256):728-732. PMid:19587681 [ Links ]
24. Derry LA. A burial diagenesis origin for the Ediacaran Shuram-Wonoka carbon isotope anomaly. Earth Planet Sci Lett. 2010;294(1-2):152-162. http://dx.doi.org/10.1016/j.epsl.2010.03.022 [ Links ]
25. Brain CK, Hoffmann K-H, Prave AR, Fallick AE, Coetzee J, Botha AJ. Interpretive problems in a search for micro-invertebrate fossils from a Neoproterozoic limestone in Namibia. Palaeontologia Africana. 2001;37(1- 12):1-12. [ Links ]
26. Aitchison J, Brown JAC. The lognormal distribution. Cambridge: Cambridge University Press; 1957. [ Links ]
27. Fallick AE, Pillinger CT, Stephenson A, Housley RM. Concerning the size distribution of ultrafine iron in lunar soil. Abstracts of papers presented to the Fourteenth Lunar and Planetary Science Conference. 1983;14:185-186. [ Links ]
28. Javaux EJ, Marshall CP, Bekker A. Organic walled microfossils in 3.2 billion year old shallow-marine siliciclastic deposits. Nature. 2010;463(7283):934-938. http://dx.doi.org/10.1038/nature08793, PMid:20139963 [ Links ]
29. Hoffmann K-H, Condon DJ, Bowring SA, Crowley JL. U-Pb zircon date from the Neoproterozoic Ghaub Formation, Namibia: Constraints on Marinoan glaciation. Geology. 2004;32(9):817-820. http://dx.doi.org/10.1130/G20519.1 [ Links ]
30. Bosak T, Lahr DJG, Pruss SB, Macdonald FM, Dalton L, Matys E. Agglutinated tests in post-Sturtian cap carbonates of Namibia and Mongolia. Earth Planet Sci Lett. 2011;308(1-2):29-40. http://dx.doi.org/10.1016/j.epsl.2011.05.030 [ Links ]
31. Rothman DH, Hayes JM, Summons RE. Dynamics of the Neoproterozoic carbon cycle. Proc Nat Acad Sci USA. 2003;100(14):8124–8129. http://dx.doi.org/10.1073/pnas.0832439100, PMid:12824461, PMCid:166193 [ Links ]
32. Hoffman PF, Schrag DP. The snowball Earth hypothesis; testing the limits of global change. Terra Nova. 2002;14(3):129–155. http://dx.doi.org/10.1046/j.1365-3121.2002.00408.x [ Links ]
33. Fike DA, Grotzinger JP, Pratt LM, Summons RE. Oxidation of the Ediacaran Ocean. Nature. 2006;444(7120):744-747. http://dx.doi.org/10.1038/nature05345, PMid:17151665 [ Links ]
34. Canfield DE, Poulton SW, Narbonne GM. Late-Neoproterozoic deep-ocean oxygenation and the rise of animal life. Science. 2007;315(5808):92-95. http://dx.doi.org/10.1126/science.1135013, PMid:17158290 [ Links ]
 Correspondence to:
Correspondence to:
Anthony Prave
Postal address: Department of Earth Sciences, University of St Andrews
St Andrews KY16 9AL, UK
Email: ap13@st-andrews.ac.uk
Received: 09 Mar. 2011
Accepted: 30 July 2011
Published: 18 Jan. 2012
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}