










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.108 no.1-2 Pretoria ene. 2012
RESEARCH ARTICLE
Nyctereutes terblanchei: the raccoon dog that never was
Sally C. Reynolds
Institute for Human Evolution, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
Fossils of the raccoon dog (genus Nyctereutes) are particularly rare in the African Plio-Pleistocene record, whilst the sole living representative, Nyctereutes procyonoides, is found in eastern Asia and parts of Europe. In southern Africa, only one fossil species of raccoon dog has been identified - Nyctereutes terblanchei. N. terblanchei is recognised from a handful of Plio-Pleistocene sites in South Africa: Kromdraai, Kromdraai-Coopers and Sterkfontein in Gauteng, as well as Elandsfontein in the Western Cape Province. The validity of this species identification was questioned on the basis of the rarity of southern African fossils assigned to Nyctereutes, that is, fewer than 10 specimens have been identified as Nyctereutes. This study examined this fossil sample of the raccoon dog from the Gauteng sites and compared dental and cranial metrics of the fossil with samples of modern canids and published data. Morphological traits used to distinguish Nyctereutes, such as the pronounced subangular lobe on the mandible and the relatively large size of the lower molars, were observed to be variable in all samples. Analysis showed that the size of the dentition of the southern African fossil samples was larger than that of living raccoon dogs, but fell well within the range of that of African jackals. These results suggest that fossil Nyctereutes cannot be distinguished from other canid species based on metric data alone, and may only be diagnosable using combinations of non-metric traits of the dentition and skull. However, based on the degree of morphological variability of the traits used to diagnose Nyctereutes, as well as the rarity of this genus in the African fossil record, these fossils are more likely to belong to a species of jackal or fox.
Introduction
Modern raccoon dogs belong to a single Asian species, Nyctereutes procyonoides, with six recognised subspecies that are well adapted to colder climates.1,2 The raccoon dog has also been introduced to European countries such as Russia and Finland for commercial farming in the fur industry. Raccoon dogs share many characteristics with medium-sized canids, including their omnivorous diets and the morphology of their skulls and dentition (Figure 1). They are, however, unique amongst canids in that they hibernate in the winter.2 Extant raccoon dogs are the size of large foxes; the raccoon dog possesses a smaller average body size than the black-backed jackal, with a median mass of about 4.0 kg,1 compared with ranges of 6.6 kg - 7.9 kg for the black-backed jackal and 8.3 kg - 9.4 kg for the side-striped jackal.3

At present, there are three jackal species in Africa: the arid-adapted golden jackal (Canis aureus) that occurs in northern and eastern Africa, the black-backed or silver-backed jackal (Canis mesomelas) that is found in eastern and southern Africa and the side-striped jackal (Canis adustus) that is found in sub-Saharan Africa up to the borders of the equatorial forest.3 DNA studies examining the phylogenetic relationships of 23 extant Canidae species suggest that Canis mesomelas and Canis adustus cluster close together, within a larger group of 'wolf-like canids', whilst the raccoon dog clusters with the African bat-eared fox (Otocyon megalotis) in a 'basal canid' position.4
Fossil canids, such as C. mesomelas and C. adustus and smaller foxes, such as the Cape fox (Vulpes chama), are common in the deposits from the Pliocene and well into the late Pleistocene and early Holocene in southern Africa.5 Elsewhere on the continent, Nyctereutes appears in the deposits of north-western Africa. The presence of a small canid with clearly defined mandibular traits indicative of Nyctereutes has been reported from the Moroccan site of Ahl al Oughlam.6 Recently, a new species of fossil raccoon dog has been announced from the Lower Awash site of Dikika, Ethiopia. This species, Nyctereutes lockwoodi, comes from deposits dated to approximately 3.20 Ma.7 A possible primitive Nyctereutes has also recently been reported from Laetoli.8 Elsewhere, several fossil raccoon dog species are known from Pliocene deposits of Western Europe and China and these specimens are larger than modern day raccoon dogs, suggesting a size decrease in the lineage of Nyctereutes from the Pliocene to the present.9
Morphologically, the genus Nyctereutes closely resembles the African jackals and foxes (genera Canis and Vulpes, respectively). In general, the raccoon dog can be distinguished by its smaller body size, shorter tooth row, reduced carnassial blade and relatively larger molars, suggestive of a greater degree of crushing ability, than can be observed in either Canis or Vulpes.10,11 The presence of a marked subangular lobe on the mandible is observed in modern raccoon dogs, as well as in the samples of later, more derived fossils of this genus. However, primitive fossil samples attributed to the raccoon dog genus do not appear to have this trait.8
The South African fossil raccoon dog species, Nyctereutes terblanchei, was first identified as Thos terblanchei from collections by Robert Broom. The skull and mandibular specimen from Kromdraai A (KA 1290), in the Sterkfontein Valley (now part of the Cradle of Humankind World Heritage Site), is the type specimen for this species.12 A second area from where these fossils were collected, was a locality known only as 'KA 2' which was labelled as Kromdraai-Coopers (two sites lying very close to Sterkfontein). Subsequently, the exact location of this site has been lost, and the isolated Nyctereutes material (n = 8 fossils) is now housed in the Palaeontology Department of the Ditsong Natural History Museum in Pretoria, South Africa (at that time called the Transvaal Museum). The seven specimens from the KA 2 site comprise five partial mandibles (KA. 2 3 YY; K2.82; K2.81; KA.72; KA2. 82) and two maxillary specimens (KA. 2 83 and KA. 2 3). Other specimens assigned to this species include a left maxilla from Sterkfontein Member 5 (SE 125) and one additional specimen (Specimen SAM 15605) from the Middle Pleistocene site of Elandsfontein (lying approximately 100 km from Cape Town).11 The specimens from Sterkfontein and Elandsfontein were not considered as part of this study.
In the original description of the material, Ficcarelli and colleagues11 noted that N. terblanchei showed a marked subangular lobe on the mandible and an ascending ramus with a more anterior orientation. When viewed as a whole, the ascending ramus and the horizontal corpus of the mandible appear to intersect at right angles.11 The basal portion of the ascending ramus is massive, but the horizontal portion shows a distinct taper towards the mandibular symphysis, whilst retaining a straight inferior border. However, the authors concluded that, of the small sample of southern African fossils, only KA 1290 and K2.82 could confidently be assigned to N. terblanchei.11
Despite the many new discoveries in the Cradle of Humankind and various studies which have reviewed and synthesised the fossil carnivores of this important hominin region,5,13,14,15,16,17 no further fossil raccoon dog material has come to light since the study by Ficcarelli and colleagues11. This absence suggests either: (1) that the raccoon dog is underdiagnosed in the South African fossil sites, as was previously suggested,11 or (2) that the sample assigned to N. terblanchei may represent another canid species. Certainly, there are carnivore species that are only rarely recovered in the Cradle of Humankind sites, such as the cheetah (Acinonyx jubatus) and certain sabre-toothed cat species (Homotherium latidens).5 This study compared extant and fossil jackals, as well as extant and fossil raccoon dog data, to the fossil sample from the Sterkfontein Valley to assess the likelihood of N. terblanchei being a rare, but valid, species identification or, alternatively, that the fossils represent a different canid species. The question is of importance to the reconstruction of fossil faunal communities and the understanding of the causes of Pleistocene extinctions.
Materials and methods
Specimens and measurements
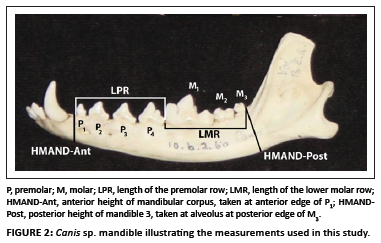
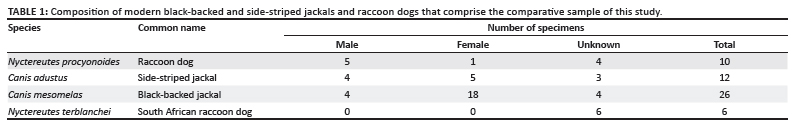
The skulls and mandibles of modern African jackals belonging to C. adustus and C. mesomelas (n = 38) and raccoon dogs from China, Vietnam and Korea (n = 10) were measured in the Natural History Museum (London, UK) (Table 1). The fossil specimens assigned to N. terblanchei are held at the Palaeontology Department of the Ditsong Natural History Museum (Pretoria, South Africa). Measurements were taken of a sample of fossil canids from Sterkfontein, Swartkrans and Kromdraai for comparison; these specimen numbers are provided in Table 2. The measurements used in this study are defined in Table 3 and illustrated in Figure 2. Additional published data for the Ethiopian Nyctereutes specimens, as well as dental measurements from the Chinese and Western European raccoon dog fossils, are also included.7,9 Means and ranges of the measurements of the samples analysed are presented in Tables 4a-f.

Site dates
The N. terblanchei type site, Kromdraai A, is younger than the Olduvai Event, that is, younger than 1.95 Ma.18 New dates for Sterkfontein Member 4 show ages of 2.65 Ma - 2.01 Ma, based on uranium-lead and uranium-thorium isotopic methods,19 whilst a second study used palaeomagnetic methods to estimate a new age of 2.58 Ma - 2.16 Ma.20 Sterkfontein Member 5 dates to between 2.0 Ma and 1.4 Ma.21 The Sterkfontein Dump 7 fossils most likely derive from the Lincoln Cave deposits; these sediments have been dated to between 353 ka and 115 ka, using uranium-thorium methods.22 The Post-Member 6 material is undated, but is presumed here to be contemporaneous with the Lincoln Cave.22 Finally, Swartkrans Member 2 has been dated to 1.1 Ma based on faunal characteristics.23
Results
Morphological traits of the mandible
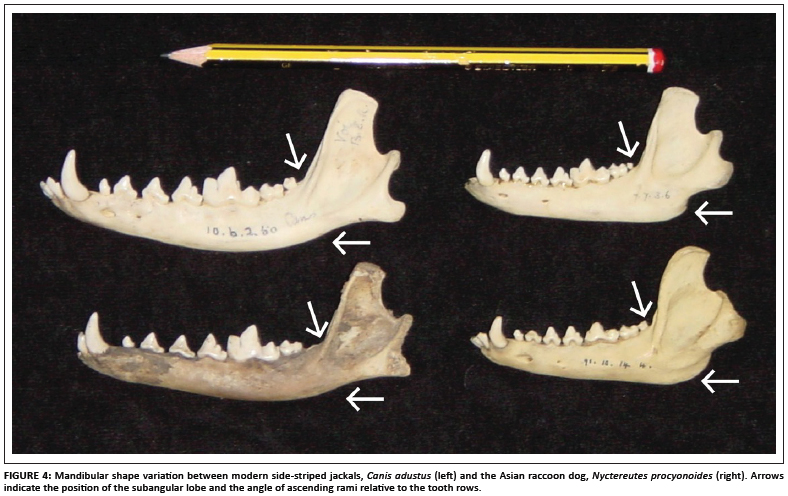
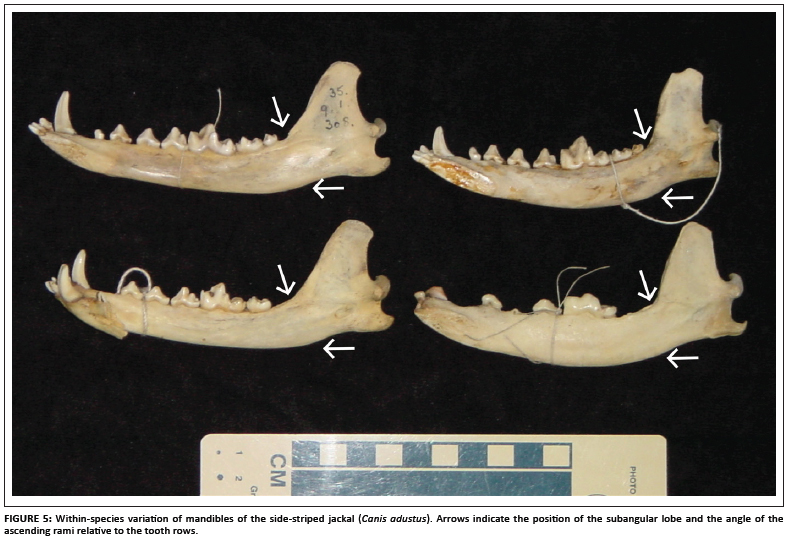
The presence of the subangular lobe of the mandible is an important characteristic of the fossil sample of Nyctereutes (Figure 3). Certain of the fossil specimens exhibit strong subangular lobes (e.g. specimens KA2. 81 and K2.82, as well as the type specimen KA 1290 illustrated elsewhere),11 whilst other specimens possess a lesser subangular lobe which overlaps with the range of variation seen in modern jackals (such as specimen KA. 72). The fossil species from Ethiopia possesses a well-developed subangular lobe.7 The subangular lobe is well developed in the modern raccoon dogs, especially when compared to the side-striped jackal (Figure 4). However, within a sample of side-striped jackals, there is also variation in the degree to which the subangular lobe is evident (Figure 5). The putative Nyctereutes from Laetoli has no subangular lobe (but a very Nyctereutes-like dentition).8
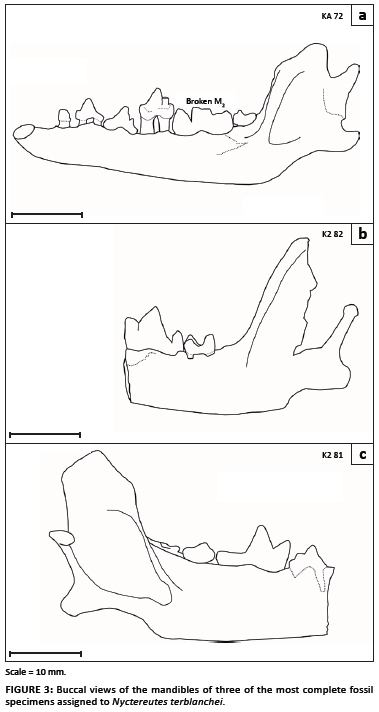
The second important characteristic is the orientation of the ascending ramus, which is almost 90 degrees in the fossil Nyctereutes11 (Figure 3). However, this angle is variable, both in the modern samples (Figures 4 and 5), as well as the fossils, with specimens such as KA.82 showing a relatively acute angle that is not so clearly marked in the other specimens (KA.72 and K2.81). The type specimen also shows an acute angle.11 In contrast, the angle observed in the published image of N. lockwoodi from Ethiopia is quite obtuse.7 Both the occurrence of the subangular lobe and the angle of the coronoid process are variable, thereby diminishing the overall usefulness of these traits as indicators of Nyctereutes. However, the development of the increasingly marked subangular lobe through time may be a fruitful avenue for future research on the fossil raccoon dogs.
Metric traits of the mandible
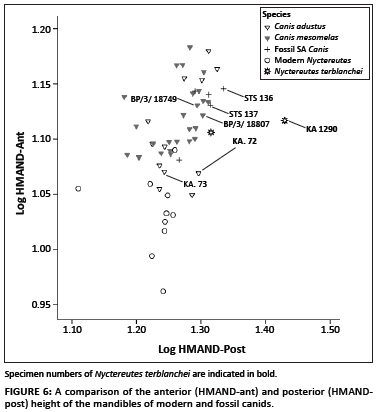
To compare the taper between the posterior and anterior portions of the ramus, log 10 values of these heights (HMAND-ant and HMAND-post) were plotted against each other (Figure 6). In modern raccoon dogs, the shape of the horizontal ramus of the mandible is relatively straight with some sloping towards the anterior of the mandible being evident. Within the relatively small sample of raccoon dogs, there is quite a marked amount of variation evident. Although variation in the mandibular heights is visible within the samples of C. mesomelas and C. adustus, the fossil Nyctereutes specimens (KA.72 and, in particular, the type specimen KA 1290) fall outside of the range of modern jackals and raccoon dogs (Figure 6). In contrast, the proportions of the mandibular traits of the fossil Canis sp. overlap with those of modern jackals, and fall particularly close to those of specimens of C. adustus, the side-striped jackal (Figure 6). There are, however, only five fossil jackal specimens available for comparison: specimen KA.73 from Kromdraai A, STS 137 and STS 136 from Sterkfontein Member 4 'Type Site' and specimens BP/3/18734 and BP/3/18807 from Lincoln Cave 'Dump 7'. The specimen assigned to Nyctereutes (KA.72) does possess a gradually tapering mandible (i.e. low anterior height and greater posterior height of the mandible).
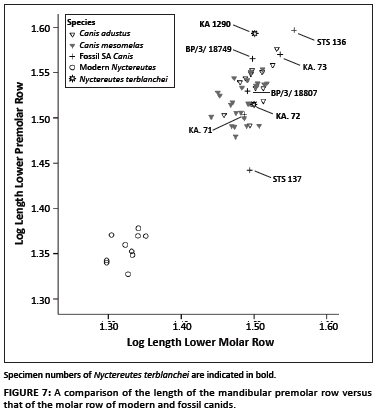
For the log 10 lengths of the lower premolar and molar rows, there is a clear separation between the cluster of modern raccoon dogs and the modern and fossil jackals and fossil N. terblanchei (Figure 7). One fossil specimen (KA. 72) falls within the latter group, whilst the type specimen, KA 1290, is larger than that of the modern black-backed jackals and side-striped jackals. The majority of the fossil Canis specimens fall well within the size range of modern jackals, with the exception of the STS 137 jackal from Sterkfontein Member 4 that is markedly smaller (Figure 7), and the STS 136 specimen which is markedly larger. The STS 137 specimens were originally published as a new species, Canis brevirostris, and the STS 136 specimen as part of the type specimen of Canis antiquus.24,25 Whilst the taxonomic attributions of these two species are not well understood, this lack of understanding does not affect the position of the N. terblanchei specimens relative to C. mesomelas, C. adustus and the other fossil canids.
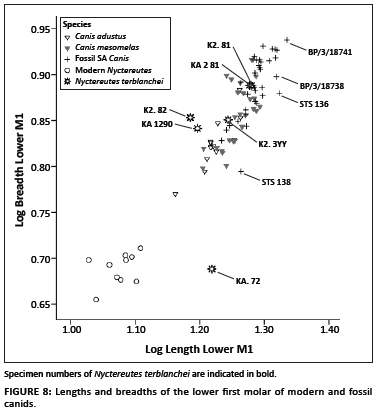
The dimensions of the log 10 lengths and breadths of the lower first molars show that three N. terblanchei specimens (K2.81; KA2. 81 and K2. 3 YY) fall within the range of variation of modern African side-striped and black-backed jackals (Figure 8). Two Nyctereutes specimens, namely K2.82 and the type specimen KA 1290, have slightly greater breadths relative to the remaining fossil Nyctereutes and Canis sp. sample. In contrast, the Nyctereutes specimen with the very narrow breadth (KA.72) is from an old individual with advanced wear on the dentition, which may have contributed to the narrowness of this specimen.
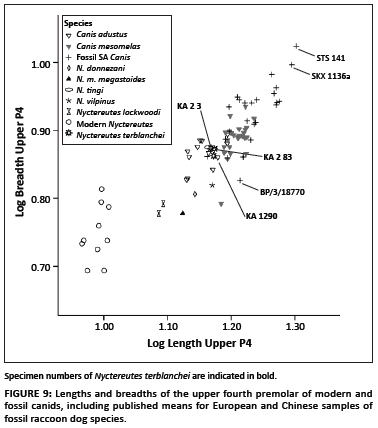
The log transformed dimensions of the upper fourth premolar show very similar patterns to those of the lower first molars (Figure 9). Modern raccoon dogs have significantly smaller upper fourth premolars (in both length and breadth) than the fossil Nyctereutes sample from South Africa. There is a certain degree of overlap in the breadths between the modern Nyctereutes specimens and those of the published specimens of N. lockwoodi from Ethiopia (Figure 9). There is also a similarity between the fossil Nyctereutes specimens from Ethiopia and the published means reported for the European fossil Nyctereutes (N. m. megastoides and N. donnezani). In contrast, the three fossil raccoon dog specimens from South Africa (KA. 2 3; KA. 2 83 and KA 1290) show overlap with the modern jackals. The largest upper fourth premolar dimensions are those of Sterkfontein Member 4 assigned to C. antiquus (STS 141) and the Swartkrans Canis sp. specimen (SKX 1136a). These specimens fall well outside the range of variation for the modern samples of black-backed and side-striped jackals (Figure 9).
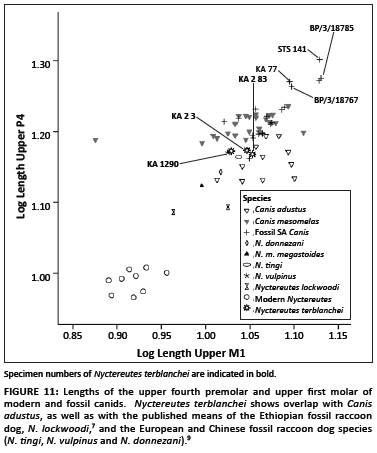
When combinations of dental metrics are considered, a similar pattern becomes evident. The log transformed length of the upper fourth premolar against the length of the upper first molar (Figure 10) shows that modern Nyctereutes are once again the smallest of all the species considered here, with the published N. lockwoodi fossils from Ethiopia having an intermediate position between the modern raccoon dogs and the modern jackals (C. mesomelas and C. adustus). The three specimens of N. terblanchei (KA 1290, KA. 2 3 and KA. 2 83) fall close to the published means of the European raccoon dog species, but also well within the range of the C. adustus and C. mesomelas specimens. The specimens with the largest metrics examined here are those derived from Sterkfontein Member 4, which were previously assigned to C. antiquus, and the unpublished Canis sp. specimens from Sterkfontein Dump 7 (BP/3/18785 and BP/3/18767).

A comparison of the log transformed lower first and second molar lengths shows a clear separation between C. adustus and C. mesomelas, with the former possessing larger lower second molar lengths (Figure 11). The five N. terblanchei specimens overlap largely with the C. adustus specimens, with K2.81 and KA2. 81 being slightly larger than the extant jackals. The largest metrics were those of the type specimens of C. antiquus (STS 136) and C. brevirostris (STS 137). The remaining fossil Canis spp. overlap with the extant C. mesomelas (Figure 11).
Discussion
The Plio-Pleistocene carnivore guild of Africa contains numerous species which are extinct today, including the sabre-toothed cats (genera Megantereon, Homotherium and Dinofelis) and the long-legged hunting hyaenas (genus Chasmaporthetes). Within extant carnivore genera, such as spotted hyaenas (Crocuta crocuta), lions (Panthera leo) and leopards (Panthera pardus), there are changes through time in body size related to climate changes.26 Specifically, body-size increases are related to deteriorating climatic conditions, or increasing seasonality.26 Thus the process of identifying within-lineage size shifts, versus the existence of completely different species or genera, is complicated given the fragmentary nature of the fossil material with which to choose between such alternatives.
The diagnosis of the genus Nyctereutes rests largely on the presence of three primary features: (1) the subangular lobe on the mandible, (2) the forward rotation of the ascending rami and (3) the progressive distomedial taper of the mandibular ramus.11 However, not all of these traits need necessarily be present: for instance, certain north-west African specimens assigned to Nyctereutes lack the subangular lobe.6 The enlarged subangular lobe provides a larger insertion point for the digastric muscle in fossil and modern canids,7 suggesting that this feature is related primarily to dietary behaviour, and is perhaps less useful for distinguishing between species that behave similarly. In modern raccoon dogs and jackals, these traits are present in the majority of specimens, but the degree to which they are pronounced is variable, thus suggesting that they are of limited usefulness for distinguishing between species. The results indicate that, in terms of size and shape even for combinations of dental metrics, N. terblanchei falls well within the range of variation of the African jackals, in particular the side-striped jackal (Canis adustus). Whilst it is possible that N. terblanchei can only be definitively diagnosed using non-metric traits, such as the morphology of the entoconid and hypoconid on the talonid of the lower first molar, or subtle variations in the cranial morphology, this present study, using only metric traits, could find no metric or combinations of metrics that could be used to distinguish successfully between the fossil raccoon dog and C. mesomelas and C. adustus.
The second point for discussion is that fossil jackals examined in this study were larger in the past relative to modern Canis conspecifics. Larger Plio-Pleistocene jackals may mean that: (1) environments in which they were living were more seasonal or arid, thus promoting selection for larger body sizes as has been documented in other African faunas of this time period26 or (2) selective forces led to a different pattern of community-wide character displacement in the Pleistocene than today. Jackal species in eastern Africa demonstrate morphological variation in areas where their distributions overlap, presumably to reduce competition between sympatric species.27 Given the presence of such size variability, it is highly possible that the sample of Nyctereutes represents the lower end of the fossil variation in jackals, instead of a completely different species.
Although the southern African 'raccoon dog' dentition appears very similar to other raccoon dog tooth metrics published from Pliocene deposits in France and China (Figures 7 and 8), this similarity may simply reflect that these canids were living in comparable environments, and not necessarily that they are taxonomically related to each other. The probability of raccoon dogs dispersing into sub- Saharan Africa from Europe and China, whilst slight, cannotbe discounted. However, even larger carnivores, such as the African wild dog (Lycaon pictus), with a larger body size and a greater degree of mobility, show that the gene flow has been affected by barriers, such as the African Rift, that have contributed to the genetic divergence observed in the wild dogs of eastern and southern Africa.28 It is therefore improbable that this small sample of raccoon dogs represents a valid species, but rather that these samples represent a morphological variant of a more common canid species.
This conclusion may also be true of the two other fossil canid species: C. brevirostris and C. antiquus.
Another carnivore species?
In the Plio-Pleistocene, a densely populated carnivore guild existed in the Sterkfontein Valley, although not all carnivore species were continuously present in all parts of the Cradle of Humankind over the same time period.5,17 Within the modern African carnivore guild, limited resources (such as prey and denning sites) mean that only a certain number of carnivores can be accommodated. Increased numbers of carnivore species would mean reduced availability of food resources, and more competition between large and small carnivore species. Where carnivores occur in high densities, they exhibit a range of behaviours to diminish competition between predators, such as avoidance, kleptoparasitism and interspecific aggressive behaviour.17 Adding another small carnivore species to this guild, such as N. terblanchei, would result in increased competition for denning sites and food resources with other small carnivores such as the black-backed and side-striped jackals, as well as the Cape fox (Vulpes chama). Additionally, the bat-eared fox (Otocyon megalotis) and the specialised ant-eating aardwolf species (Proteles cristatus) would have increased competition for insect food resources.
Conclusions
The validity of the South African raccoon dog as a species is not only vital to the reconstruction of carnivore guilds which existed in the past, but also to understanding the numerous extinctions of Pleistocene faunas. The traits used to define the fossil raccoon dog included a pronounced subangular lobe on the mandible and the ascending ramus with an anterior orientation, as well as small premolars in combination with large molars. However, this study indicated that these traits show reasonable amounts of variation in modern populations of Asian raccoon dogs (N. procyonoides), as well as in the African jackals (C. mesomelas and C. adustus). For all metrics considered in this study, the fossil sample of the raccoon dog fell well within the range of variation for the African jackals. Future work will need to focus on the generic traits which separate the Canis lineage from the Nyctereutes lineage, such as the transverse crest linking the entoconid and hypoconid on the lower first molar talonid, to determine if N. terblanchei most resembles racoon dogs or members of the Canis lineage. However, the dearth of additional fossil specimens since the species was named suggests that the sample assigned to the raccoon dog most likely represents a variant Plio-Pleistocene population of Canis, a genus that is well represented in the Cradle of Humankind sites.
Acknowledgements
This data was collected during the period of my National Research Foundation (NRF) postdoctoral fellowship hosted by the School of Anatomical Sciences, University of the Witwatersrand Medical School. I would like to thank my hosts, Prof. Ron Clarke and Dr Kathleen Kuman, for their support of my scientific endeavours. I thank the Museum curators of the Ditsong Natural History Museum (formerly the Transvaal Museum, Pretoria), the Bernard Price Institute, University of the Witwatersrand (Johannesburg) and the Mammal Department of the Natural History Museum, (London) for access to their comparative collections. This research received support from the 2008 SYNTHESYS Project http://www.synthesys.info/ which is financed by European Community Research Infrastructure Action under the FP6 'Structuring the European Research Area' Programme. I thank Lars Werdelin, Alan Morris and an anonymous reviewer for insightful comments that greatly improved this manuscript.
Competing interests
I declare that I have no financial or personal relationships which may have inappropriately influenced me in writing this article.
References
1. Ward OG, Wuster-Hill DH. Mammalian species no. 358: Nyctereutes procyonoides. Am Soc Mammalogists. 1990;358:1-5. [ Links ]
2. Nyctereutes procyonoides (raccoon dog) IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1 [homepage on the Internet]. c2011 (cited 2011 Jul 14). Available from: www.iucnredlist.org [ Links ]
3. Estes RD. The behaviour guide to African mammals, including hoofed mammals, carnivores, primates. Berkeley: University of California Press; 1991. [ Links ]
4. Bardeleben C, Moore RL, Wayne RK. A molecular phylogeny of the Canidae based on six nuclear loci. Mol Phylogenet Evol. 2005;37:815-831. http://dx.doi.org/10.1016/j.ympev.2005.07.019, PMid:16213754 [ Links ]
5. Reynolds SC. Where the wild things were: Spatial and temporal distribution of carnivores in the Sterkfontein Valley in relation to the accumulation of mammalian assemblages. J Taph. 2010;8(2-3):233-257. [ Links ]
6. Geraads D. Plio-Pleistocene Carnivora of northwestern Africa: A short review. Comptes Rendus Palevol. 2008;7(8):591-599. http://dx.doi.org/10.1016/j.crpv.2008.09.008 [ Links ]
7. Geraads D, Alemseged Z, Bobe R, Reed D. Nyctereutes lockwoodi N. sp.: A new canid (Carnivora: Mammalia) from the Middle Pliocene of Dikika, Lower Awash, Ethiopia. J Vert Pal. 2010;30(3):981–987. http://dx.doi.org/10.1080/02724631003758326 [ Links ]
8. Werdelin L, Dehghani R. Carnivora. In: Harrison T, editor. Paleontology and geology of Laetoli: Human evolution in context. Volume 2: Fossil hominins and the associated fauna. Vertebr Paleobiol Paleoanthropology. 2011;189–232. http://dx.doi.org/10.1007/978-90-481-9962-4_8 [ Links ]
9. Monguillon A, Spassov N, Argant A, Kauhala K, Viranta S. Nyctereutes vulpinus comb. et stat. nov. (Mammalia, Carnivora, Canidae) from the late Pliocene of Saint-Vallier (Drôme, France). Geobios. 2004;37:S183-S188. http://dx.doi.org/10.1016/S0016-6995(04)80014-7 [ Links ]
10. Ewer RF. The carnivores. Ithaca and London: Cornell University Press; 1973. [ Links ]
11. Ficcarelli G, Torre D, Turner A. First evidence for a species of raccoon dog, Nyctereutes Temminck, 1838, in South African Plio-Pleistocene deposits. Estratto dal bollettino della Societa Paleontologica Italiana. 1984;23(1):125-130. [ Links ]
12. Broom R. Some South-African Pliocene and Pleistocene mammals. Ann Transvaal Mus. 1948;21:1-28. [ Links ]
13. Turner A. New fossil carnivore remains from the Sterkfontein hominid site (Mammalia: Carnivora). Ann Transvaal Mus. 1987;34:319-347. [ Links ]
14. Turner A. Further remains of Carnivora (Mammalia) from the Sterkfontein hominid site. Palaeontol Afr. 1997;34:115-126. [ Links ]
15. Lacruz R, Turner A, Berger LR. New Dinofelis (Carnivora: Machairodontinae) remains from Sterkfontein Valley sites and a taxonomic revision of the genus in southern Africa. Ann Transvaal Mus. 2006;43:89-106. [ Links ]
16. O'Regan HJ. A revision of the Carnivora from Member 5, Sterkfontein, South Africa, based on a reassessment of published material and site stratigraphy. Ann Transvaal Mus. 2007;44:209-214. [ Links ]
17. O'Regan HJ, Reynolds SC. An ecological reassessment of the southern African carnivore guild: A case study from Member 4, Sterkfontein, South Africa. J Hum Evol. 2009;57:212–222. http://dx.doi.org/10.1016/j.jhevol.2009.04.002, PMid:19647856 [ Links ]
18. Thackeray JF, Kirschvink JL, Raub TD. Palaeomagnetic analysis of calcified deposits from the Plio-Pleistocene hominid site of Kromdraai, South Africa. S Afr J Sci. 2002;98:537-540. [ Links ]
19. Pickering R, Kramers JD. Re-appraisal of the stratigraphy and determination of new U-Pb dates for the Sterkfontein hominin site, South Africa. J Hum Evol. 2010;59:70-86. http://dx.doi.org/10.1016/j.jhevol.2010.03.014, PMid:20605190 [ Links ]
20. Herries AIR, Shaw J. Palaeomagnetic analysis of the Sterkfontein palaeocave deposits: Implications for the age of the hominin fossils and stone tool industries. J Hum Evol. 2011;60(5):523–539. http://dx.doi.org/10.1016/j.jhevol.2010.09.001, PMid:21392817 [ Links ]
21. Kuman K, Clarke RJ. Stratigraphy, artefact industries and hominid associations for Sterkfontein Member 5. J Hum Evol. 2000;38:827-847. http://dx.doi.org/10.1006/jhev.1999.0392, PMid:10835264 [ Links ]
22. Reynolds SC, Vogel JC, Clarke RJ, Kuman KA. Preliminary results of investigations at Lincoln Cave, Sterkfontein, South Africa. S Afr J Sci. 2003;99:286-288. [ Links ]
23. Vrba ES. Ecological and adaptive changes associated with early hominid evolution. In: Delson E, editor. Ancestors, the hard evidence. New York: Alan R. Liss, 1985; p. 63-71. [ Links ]
24. Brain CK. The hunters or the hunted? An introduction to African cave taphonomy. Chicago: University of Chicago; 1981. [ Links ]
25. Ewer RF. The fossil carnivores of the Transvaal Caves: Canidae. Proc Zool Soc London. 1956;126(1):97–120. http://dx.doi.org/10.1111/j.1096-3642.1956.tb00427.x [ Links ]
26. Reynolds SC. Mammalian body size changes and Plio-Pleistocene environmental shifts: Implications for understanding hominin evolution in eastern and southern Africa. J Hum Evol. 2007;53(5):528–548. http://dx.doi.org/10.1016/j.jhevol.2007.01.014, PMid:17915290 [ Links ]
27. Van Valkenburgh B, Wayne RK. Shape divergence associated with size convergence in sympatric East African jackals. Ecology. 1994;5(6):1567- 1581. http://dx.doi.org/10.2307/1939618 [ Links ]
28. Girman DJ, Vilà C, Geffen E, et al. Patterns of population subdivision, gene flow and genetic variability in the African wild dog (Lycaon pictus). Mol Ecol. 2001;10:1703–1723. http://dx.doi.org/10.1046/j.0962-1083.2001.01302.x, PMid:11472538 [ Links ]
 Correspondence to:
Correspondence to:
Sally Reynolds
Postal address: 17 Rue Docteur Fighièra
Nice 06300, France
Email: reynolds@ipgp.fr
Received: 15 Jan. 2011
Accepted: 26 July 2011
Published: 13 Jan. 2012
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}