










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.108 no.1-2 Pretoria Jan. 2012
REVIEW ARTICLE
Why did schistosomiasis disappear from the southern part of the Eastern Cape?
Chris C. AppletonI; Inbarani NaidooII
ISchool of Biological and Conservation Sciences, University of KwaZulu-Natal, Durban, South Africa
IIMalaria Research Unit, Medical Research Council, Durban, South Africa
ABSTRACT
We reviewed the early literature and maps of the occurrence of urogenital schistosomiasis (bilharzia) in the Eastern Cape, South Africa from the 1860s until its decline from about 1900 and reappearance in 2002. Although this decline in transmission has received little attention to date, clinical descriptions of the disease over this period indicate that infection was common, probably patchy, although sometimes with severe morbidity. The long period of quiescence between 1900 and 2002 is thought to be as a result of several factors, but primarily because of the impact of the area's cold winters and drought-prone climate on the survival and reproduction of both the snail intermediate host Bulinus africanus and the intramolluscan stages of the parasite. The concept of an outbreak area is invoked to describe the occurrence of intense urogenital schistosomiasis transmission in localised areas for relatively short periods of up to 35 years in this the southernmost part of its range in Africa, a suboptimal environment for transmission.
Introduction
Monitoring changes in the distribution of urogenital schistosomiasis (bilharzia)1 in South Africa is important with respect to public health, especially the proposed national helminthiasis control programme.2 However, the fact that schistosomiasis is not, and never has been, a notifiable disease in South Africa makes monitoring and surveillance difficult. Whilst there is no indication that the urogenital form of the disease is spreading within the country, there is clear evidence in the literature for a historical contraction, albeit temporary, in its geographic range in what is now the Eastern Cape Province. In this paper, we review the historical literature on urogenital schistosomiasis in the Eastern Cape, south of the Buffalo River, to reconstruct the clinical epidemiology of the disease from its discovery in 1863 to circa 1900 and to examine reasons for its subsequent disappearance from this area until 2002.
Historical account
Urogenital schistosomiasis was diagnosed in boys from Uitenhage in the Eastern Cape, South Africa, by Dr J. Harley, a general practitioner in Port Elizabeth, in 1863.3 These diagnoses were only 11 years after the publication of Dr Theodor Bilharz' report4 on his discovery of the causative agent of the disease, the blood fluke Schistosoma haematobium, in the mesenteric vein during a post-mortem in Kasr el Aini Hospital, Cairo. Harley's 1864 report3 was the first record of the disease in sub-Saharan Africa and led him to believe (correctly) that it occurred throughout the length of the continent and that his finding was at the southern limit of its range. The decline of the disease from this same part of the Eastern Cape from about 1900 has attracted little attention until now.
Epidemiology and transmission of urogenital schistosomiasis
Although no statistics are available, there was consensus amongst case reports between 1864 and 18993,5,6,7,8,9 in respect of several, now widely recognised, characteristics of urogenital schistosomiasis. All age groups were infected but infection was commonest in children, mostly boys, between 3-years and 16-years old with a history of contact with local waterbodies. The most frequent symptom, haematuria, usually disappeared before the age of 20 because recreational swimming had stopped by this time, thus reducing the chances of further infection.9 As for the longevity of the disease, haematuria was observed, often continuously, in individual patients for up to 10 years9 but, following Harley3, the infection probably persisted 'as long as life continued'. Girls and women were seldom affected because they had little contact with natural water.
Transmission was attributed to contact with rivers, usually through swimming, but apparently also occurred through contact with infested water in vleis, farm dams, irrigation furrows and even streetside canals. In terms of seasonality of transmission, Haw8 believed that spring and summer were more 'dangerous' for contracting schistosomiasis than other seasons, whilst Spencer9 claimed that children contracted the disease almost entirely in the dry winter, when rivers were reduced to a series of pools in which they swam on warm days. Presumably, Spencer meant that most cases of haematuria were diagnosed in winter because we now know that haematuria may take several years post-exposure to develop.10
In the 1890s, knowledge of the schistosome life cycle was incomplete and based on that of the liver fluke Fasciola hepatica.8,11,12 Thus the cycle was assumed to include development inside an aquatic intermediate host and this was widely thought to be a mollusc. It was only in 1915 that the intermediate host of S. haematobium in South Africa was identified as the planorbid freshwater snail Bulinus africanus.13,14
Clinical descriptions of urogenital schistosomiasis
Clinical descriptions of urogenital schistosomiasis3,5,6,7,8,9,12,15,16all noted that macroscopic haematuria and dysuria were the most common and recognisable symptoms. Although these early reports from the Eastern Cape did not give prevalence data, phrases like 'the haematuria of the Cape',3 'the remarkable frequency of haematuria among young boys',15 'It's only the redwater. We've all got it.'16 and 'very prevalent, very chronic'9 indicate that S. haematobium infection was common in the area.
Most diagnoses were confirmed microscopically by demonstrating the presence of the fluke's characteristic terminal-spined eggs in a patient's urine. In 1899 Haw17 therefore recommended that in all cases of haematuria in South Africa, urine should be examined microscopically for the disease. The frequency of haematuria and dysuria and the comments by Chute12 and others that the urine of some patients was dark red ('a deep claret colour') or 'pure blood' indicates that some of these were severe and would be classified as high-intensity infections today. Frequent reports of albuminuria, calculi and spicule-shaped or lozenge-shaped deposits of uric acid support this interpretation. However, it was not appreciated by these early authors that as many asymptomatic as symptomatic cases would have been present in the population, that is, that there would have been as many light infections with microhaematuria rather than macrohaematuria or dysuria, which would probably have gone undiagnosed. Indeed, Van der Werf and De Vlas18 recently estimated that the presence of macroscopic haematuria in a community generally corresponds to an infection intensity of >100 eggs/10 mL. At lower intensities haematuria is less commonly seen and is more likely to be as a result of other causes. Dysuria may occur at lower intensities, that is, >10 eggs/10 mL.
Reports of urine microscopy by these early observers reveal that egg output was sometimes high, up to '30 or 40 or more' per microscope slide, hundreds per terminal micturition ('teaspoonful') and several thousand per day. Eggs were often free in the urine but many, up to about 300, could be seen embedded with epithelial cells in clots of blood in strings of flocculant, dirty white mucus. Harley3,6 provided detailed drawings of these eggs and the miracidia that hatched from them.
Complications such as ureteral constrictions would seldom have been seen with the techniques available but may nevertheless have led to hydroureter, hydronephrosis and perhaps secondary infection and renal failure. However, renal and bladder colic, blood clots and calculi resulting from these complications were commonly reported and their passage down the urinary tract was usually accompanied by symptoms such as lassitude, tenderness of the kidneys and pain, sometimes severe, in the lumbar region radiating to the ureters, urethra, bladder and penis, particularly at the end of micturition.5,6,7,8,9 Appendicitis and anaemia were also common and the growth of infected children was often impaired.12
Microhaematuria is today regarded as one of the earliest symptoms of urogenital schistosomiasis, starting as soon as 4 weeks after exposure.10 Macroscopic haematuria, often accompanied by dysuria, usually takes longer to appear, up to 3 months post-exposure, and sometimes up to 2 or 3 years after exposure. Calculi may take 5-7 years to develop. Symptoms such as these thus point to chronic S. haematobium infections but the pre-1900 authors cited previously were seldom able to confirm this by examining autopsy material for urinary tract lesions. Neither were cystoscopy, pyelography nor ultrasound available at the time, although by 1910 it had become recommended practice to examine all cases of haematuria cystoscopically for the characteristic pathology of the disease.19
These observations conform to those of the autopsy-based studies of Bhagwandeen20 and Cooppan et al.21 in neighbouring KwaZulu-Natal, which give a more detailed clinical picture of what the disease in the Eastern Cape may have looked like. In these studies a high proportion of patients had detectable morbidity. Many of these patients complained of haematuria and sometimes dysuria and urography or ultrasound showed abnormalities of the urinary tract. Bladder cystitis was common and involved a variety of macroscopic lesions in the bladder wall and the lower ureters. Polyps and sandy patches formed in the mucosa and submucosa and thickened the tract's walls, causing ureteral obstructions which restricted the flow of urine (hydronephrosis and hydroureter) and led to secondary infection. A smaller proportion of infected subjects was subclinical but urine microscopy would have shown microhaematuria.
Overall the morbidity indicated by the early case reports from the Eastern Cape seems comparable with that described later from KwaZulu-Natal. Some cases were undoubtedly serious and Bhagwandeen's summary of the situation in KwaZulu-Natal20 probably also applies to the Eastern Cape in the late 19th century. Bhagwandeen20 wrote: '... from this study has emerged the startling fact that urinary bilharziasis has a high bladder morbidity and that ureteric damage is not only common but is often severe'. Although Harley6 considered that the disease was rarely fatal, Turner22 in his review of schistosomiasis in South Africa subsequently recorded deaths as a result of hydronephrosis in the former Transvaal. Turner22,23 also recorded pulmonary complications and the presence of S. haematobium eggs in genital lesions. These related complications probably occurred in the study area too. Genital involvement is now known to be common in S. haematobium infections and is reflected in the changed name for the disease, urogenital schistosomiasis.1 Genital lesions are not infrequently reported from the Transkei today.24
Historical and current distribution of urogenital schistosomiasis
The southernmost locality for urogenital schistosomiasis in Africa has long been considered to be the Port Elizabeth-Uitenhage areas of the Eastern Cape, that is, approximately 34ºS. This limit is based on Harley's 1864 report of infection in children from Uitenhage which he described as common in the catchment of the Swartkops River. A survey of the literature3,5,6,7,8,9,12,15,16,25 indicates that, in addition to Port Elizabeth and Uitenhage, transmission occurred at King William's Town (the most frequently cited locality) and East London over a period of about 35 years from the 1860s to at least the late 1890s. In comparison, transmission occurring at Alice, Fort Beaufort and Graaff Reinet was reported only in the late 1860s. Single case reports from numerous other localities testify to transmission, probably at lower intensities, but the dates of these are often not given. These localities, together with others listed by Turner22, indicate that transmission occurred widely in the region and was associated with the drainages of most if not all rivers from the Baakens River in the west to the Buffalo River in the east. The Buffalo River stands out as the most frequent source of infection.
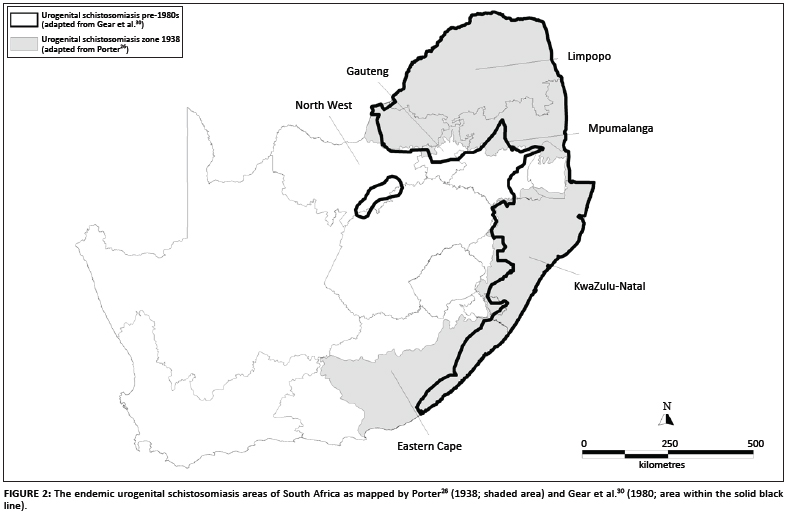
In 1938 Porter26 published the first map of the geographic distribution of S. haematobium in South Africa (Figure 1a). This map was drawn in 1934 from data collected between the 1860s and 1932 and its database combined cases that Porter had personally diagnosed, was told of by 'reliable medical sources' or had gleaned from the literature, as well as records of the intermediate host, the snail B. africanus, carrying schistosome infections. Porter's26 endemic area was bounded by localities identified in this way and so includes intervening areas where it was thought that schistosomiasis could occur. The disease's south-western limit lay along a line running north-west from Port Elizabeth to Graaff Reinet (where Porter rated transmission as 'slight'), that is, bounded by the Baakens, Swartkops and Sundays Rivers. The northern boundary lay along a transect from Graaff Reinet eastwards to Sterkstroom and Elliot at altitudes between 800 m and 1500 m. Porter noted that, whilst infection rates varied greatly over this area, transmission had been well established at Port Elizabeth and King William's Town and their environs. Porter's monograph also describes snail compatibility experiments with S. haematobium carried out in 1920 using eggs from a patient who had contracted the disease in King William's Town - evidence that transmission continued in King William's Town for some years beyond 1900.
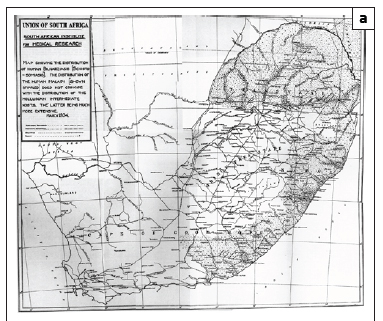
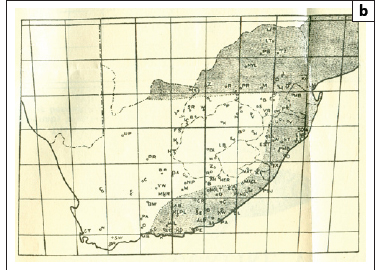
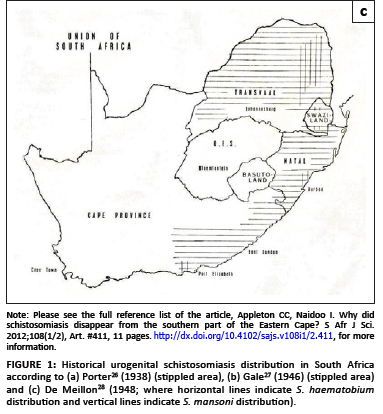
Maps similar to that of Porter's26 were published by Gale27 and De Meillon28 (Figures 1b and 1c) but showed transmission to extend westwards to George and Humansdorp, respectively. De Meillon28 noted, as had Porter26, that in the Eastern Cape (and elsewhere) the distribution of the snail host was wider than that of the disease but predicted that the disease's range might spread to correspond with that of its snail host. Later, in 1959, a survey by Pitchford and Geldenhuys29 recorded a prevalence of 0.9% (2/224) for the Uitenhage district but noted that according to the district surgeon at the time, the prevalence, presumably diagnosed from haematuria and/or urine microscopy, had been ±20% some 20-30 years earlier (i.e. in the 1930s and 1940s). Similar low prevalences were found east of the Port Elizabeth-Uitenhage-Graaff Reinet axis, namely 1.1% at both King William's Town (2/181) and East London (5/444), but, as with Uitenhage, unpublished surveillance by local health authorities had recorded higher levels of 20% - 25% for these towns several decades before; a situation that persisted until at least the late 1940s.
The Atlas of bilharzia in South Africa30 contains maps of disease prevalence data from surveys carried out between the early 1950s and late 1970s (with addenda to 1982), but it is not possible to assign exact dates to the records because the raw data are no longer available. Nevertheless, the atlas shows a much more restricted distribution for the disease in the Eastern Cape (Figure 2). Data were presented in prevalence categories with records in the 6% - 10% and 11% - 25% categories for Mount Gulu and Chalumna, 20 km and 35 km south of East London, respectively. Occasional reports to C.C.A. from local health authorities in the East London area suggest that focal transmission may have occurred there into the 1990s.
In the Eastern Cape today, it is only in the former Transkei that prevalences above 20% are encountered routinely.31,32,33 This observation was confirmed in 2000 through routine surveillance by the Eastern Cape Health Department (Meidany F 1999, personal communication, October 28). Currently the southern limit of the disease is in the southern Transkei but the specific locality is not known with certainty. Health Department surveillance suggests that it lies in the vicinity of Willowvale (±32º15'S), approximately 80 km south-east of Umtata. A prevalence of 9.1% was recorded there in 1994 (Meidany F 1999, personal communication, October 28).
Pitchford and Geldenhuys29 did not find any infections west of the Port Elizabeth-Uitenhage axis, that is, at Humansdorp, Kareedouw, the Tsitsikamma and Groot River valleys and Knysna. Interestingly, however, the two maps of the disease published during the 1940s27,28 showed transmission in the Humansdorp area, as well as beyond Humansdorp to George, but without supporting parasitological information (Figures 1b and 1c). De Meillon28 cited data from Dr Harrington, of the South African Institute for Medical Research in Port Elizabeth, who reported that he had seen 'numbers of undoubted indigenous cases' of S. haematobium from Humansdorp. Despite this report, neither Le Roux34 nor De Meillon28 actually stated that urogenital schistosomiasis had spread to Humansdorp. In fact Le Roux34 conceded that his observations on the ungulate schistosome Schistosoma mattheei did not preclude human transmission occurring at Humansdorp, and De Meillon's28 map showed that it did. It is interesting then that a later map by Pitchford and Visser35 showed three isolated areas of transmission remaining in the study area: Humansdorp, Port Elizabeth and King William's Town (Figure 3).
In summary, urogenital schistosomiasis clearly occurred in people between the Port Elizabeth-Uitenhage area and the Buffalo River, an area of ±46 000 km2, during the last four decades of the 19th century. Pockets of high transmission were located in populated areas. Prevalences at these localities had dropped to 20% - 25% by the 1930s and 1940s, or perhaps earlier - an interpretation supported by the fact that reports of transmission in the area ceased to be published around 1900. Figure 4 tracks the reports on individual foci of transmission as gleaned from reports in the literature. From Figure 4, we suggest that for about 35 years (from 1863 to 1899), the major transmission areas were in Port Elizabeth, Uitenhage, King William's Town and East London, whilst Fort Beaufort was a major transmission area for only about 4 years (1866-1869). Most other sites are represented by single reports only and are therefore thought to have been short-lived. The post-1900 decline in transmission continued such that the disease had all but disappeared from this part of the Eastern Cape by the late 1950s, with residual prevalences of <1%. This situation remains today.
Recent transmission in the Humansdorp area
Humansdorp is the type locality for the ungulate schistosome S. mattheei. This species was described by Veglia and Le Roux36 from specimens collected in 1926 during autopsies of sheep that had died following an outbreak of schistosomiasis on a farm in the area. Named after Mr S.W. Matthee, owner of the farm, S. mattheei caused heavy stock losses at the time but subsequently disappeared from the area37 because, according to Pitchford38, Mr Matthee had followed Le Roux's34 advice on control measures.
In September 2002, schistosomiasis was diagnosed near Jeffrey's Bay 24 km east of Humansdorp by the South African Institute for Medical Research in Port Elizabeth. Fifteen cases of overt haematuria were recorded by staff at Humansdorp Hospital in children aged between 7 and 15 (on average 10 years old) and all were confirmed from urine microscopy to be infected by S. haematobium with egg loads being described as 'abundant'. A count of 1993 eggs/10 mL urine was obtained for one sample processed at the University of KwaZulu-Natal (Archer CE 2002, personal communication, October 02). This count is very high by international standards and is classified as a severe intensity infection.39 Mean measurements ± s.d. for the eggs from these children were 133.6±14.9 mm x 65.5±8.8 mm (n = 25) and lie within the normal range for S. haematobium. Large amounts of mucus were present in the urine. All diagnosed children were treated with praziquantel at the standard dose of 40 mg/kg (Coetzee DO 2002, personal communication, October 08).
A visit by C.C.A. to the area from 08 to 10 October 2002 showed that 13 of the children with haematuria lived on two farms, Sonop (11 children) and Die Bult (2 children), and the remaining 2 children lived in an informal settlement, Smithstown. All localities were situated within a few kilometres of each other, less than 10 km from the coast. The parasite was probably introduced to the area by migrant fishermen from the Transkei and/or KwaZulu-Natal who worked on local squid (chokka) boats and lived in the Smithstown informal settlement.
Bulinus africanus, the snail intermediate host for S. haematobium, is known from the area and was recorded from the Kabeljous River near Jeffrey's Bay.29,30 This snail also occurs in the adjacent two rivers to the west (Seekoei and Kromme Rivers) with Kareedouw on the Kromme (33º56'S, 24º17'E) being its south-westernmost locality in Africa. The absence of the snail from rivers further westwards is probably because these rivers are generally acidic (pH 4.5 - 6.7).40 The record by Connolly41 of B. africanus from Knysna has not been confirmed.
A survey of six waterbodies in the area (three earth farm dams and three sites on the Kabeljous River) showed B. africanus to occur in one of the dams, Tweekloof. According to parents and local farmers, children from the area, including those who contracted S. haematobium, often swam in this dam on their way home from school in the nearby town of Jeffrey's Bay. Tweekloof dam is the presumed site of transmission in this outbreak.
Tweekloof dam
Tweekloof dam is located at 33º58'S, 24º55'E at an altitude of ±71 m above sea level, ±9 km north-west of Jeffrey's Bay, and is fed by a canal from the Kabeljous River. The dam has grassy banks as well as floating and emergent vegetation (Ludwigia sp. and Scirpus sp., respectively). Six adult B. africanus, measuring 13.6 mm - 14.9 mm shell height, were collected in the dam within a 30-min period. None of the adult snails was infected with schistosomes (one carried a prepatent echinostome infection) and no juvenile snails were found. Other snail species found were Lymnaea columella and Bulinus tropicus. The water of Tweekloof dam was well oxygenated, was clear with a conductivity of 713 µS/cm, had a pH close to neutral and was pale brown. Although high for fresh water, the observed conductivity is within the tolerance range of B. africanus.42 The dam is used for irrigation and watering cattle. Examination of faecal samples from the cattle that drank in the dam showed no S. mattheei, although 83.3% (15/18) were infected with the conical fluke Calicophoron microbothrium.
Enough is known of the biology of B. africanus to suggest that the failure to find juveniles in Tweekloof dam, whilst juveniles of B. tropicus were present, is meaningful. This finding suggests that in the south-western parts of the Eastern Cape, the south-westerly end of its geographic range, B. africanus may not exhibit the annual pattern of three overlapping generations identified in the lowveld of the Mpumalanga Province - a region highly endemic for S. haematobium.43 This difference is because this region of the Eastern Cape experiences long, cold winters, as shown by temperature data from Cape St Francis, the nearest meteorological station, 21 km from the study area (Figure 5).
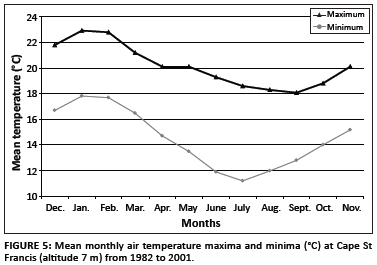
De Kock44 concluded from experimental work that South African B. africanus reproduces over a 10 ºC temperature range (19 ºC - 29 ºC) but that within this range its intrinsic rate of natural increase (rm) is low relative to other snail species. In fact, B. africanus does not survive or breed well in cold water, and at water temperatures below 20 ºC, rmdropped close to zero (0.3). With mean monthly minima below 18 ºC throughout the year in the study area (Figure 5) and mean monthly maxima below 20 ºC for 5 months (June to October), few B. africanus are likely to survive winter and those that do will only start to breed in late spring once the water has warmed to 22 ºC - 23 ºC. It is even less likely that schistosome-infected individuals will overwinter successfully, so transmission will probably depend on each spring-summer generation being infected anew. Following Pitchford and Visser45 this will mean a transmission season of only 3-4 months (late December to March) compared with 10 months in the Mpumalanga lowveld.
Pitchford and Visser35 made similar observations on B. africanus exposed to S. haematobium and S. mattheei and kept in outdoor aquaria at Cradock, one of the towns in the Eastern Cape interior (at an altitude of 885 m) where Porter26 recorded patients infected with S. haematobium. As at Cape St Francis, mean monthly minima fell below 18 ºC throughout the year but mean monthly maxima were below 20 ºC for only 2 months, June and July. Snails reproduced in summer when mean monthly maxima rose above 25 ºC but overall survival was poor with all test snails dying in winter when mean monthly minima dropped below 5 ºC. Cercarial shedding was intermittent for both schistosome species and limited to short periods between January and May. These results led Pitchford and Visser35 to suggest that neither S. haematobium nor S. mattheei was likely to become established in the area although short periods of transmission might occur.
Discussion
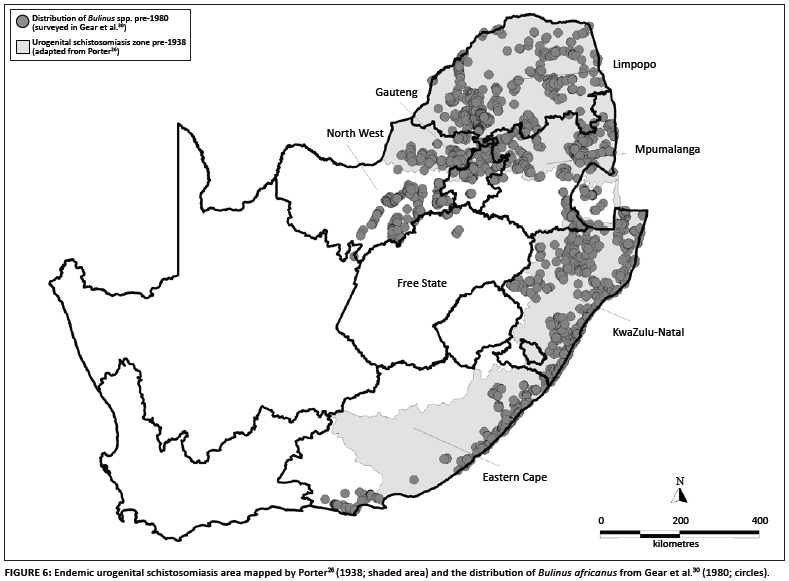
A comment by Le Roux34 that S. haematobium transmission was 'by no means widespread' in the Cape Province but was instead localised to certain well-defined areas conflicts with the later maps26,27,28 (Figures 1 and 2). However, Porter's26 map, as well as the other two which seem to be copies, included areas considered to be of 'potential' as well as areas of proven endemicity. Nevertheless, it is clear that transmission occurred at numerous places in the Eastern Cape between the early 1860s and late 1890s, but declined after about 1900. No new foci were reported for the next 100 years until that near Jeffrey's Bay in 2002. Some localised events may of course have gone unreported and Pitchford's38 remark that 'it is probably somewhat rare today' suggests that he doubted that urogenital schistosomiasis had completely disappeared in the study area. We also do not view the quiescence of the disease in this area as a reduction in the disease's geographic range but rather we classify the area as an outbreak area for urogenital schistosomiasis following the definition given by Green et al.46. A comparison between the maps of Porter26 and Gear et al.30 (Figure 6) shows this outbreak area to be approximately 12% of the total endemic area of South Africa as mapped by Porter26.
The disease's decline and subsequent quiescence in this area during the 20th century has been attributed to various environmental and biological factors. Cawston47 suggested a role for changing climatic conditions by drawing attention to a prolonged drought in the area and the vulnerability of B. africanus populations in both rivers and disjunct pools to droughts. The distribution of B. africanus given by Gear et al.30, and subsequently by Brown48 and De Kock and Wolmarans49, shows B. africanus to be absent from much of the S. haematobium area of the Eastern Cape as mapped earlier26,27,28 (Figure 6). As Gear et al.30, Brown48 and De Kock and Wolmarans49 all used confirmed B. africanus identifications from the National Freshwater Snail Collection, it can be argued that the snail's recorded absence from much of this area is either because it never occurred there or its range has retracted from the area since the 19th-century outbreaks. Alternatively, B. africanus occurs more widely in this area than these maps show, but, as discussed by Appleton and Stiles40, only as difficult-to-find relict or temporary populations in standing waterbodies associated with dykes in the Karoo sediments that dominate the area. Support for the opinion that standing waterbodies within the study area may be colonised by B. africanus, comes from the fact that three large dams have been built in the area and B. africanus has been reported to occur in all of them.50
We believe that the occurrence of B. africanus in the study area, the marginal south-west of its range, is both sporadic and unpredictable and this is key to the recorded decline in schistosomiasis transmission. As De Kock44 noted for such areas, a severe winter followed by a poor rainy season could lead to the extirpation of many of these populations. Immigration and repopulation are also likely to be slow under these conditions. Even though B. aftricanus is considered to be an only 'moderately successful' aestivator,48 adults can survive for prolonged periods in sediment with a surface moisture content as low as 1.8% - 1.9% at temperatures up to 45 ºC.51 The ephemeral nature of these habitats, the snail's tolerance of adverse conditions and its high innate capacity for increase will undoubtedly lead to dramatic fluctuations in its numbers, such that its survival in any given year will depend largely on the rainfall pattern and the length of time for which its habitats are dry.49
In her analysis of the historical climate of Africa, Nicholson52,53 did in fact identify a severe drought in the Eastern Cape from 1895 to 1920. Rainfall in the region during the preceding period 1868-1895, which covers most of the outbreaks discussed earlier, was 'variable' but with evidence for increased precipitation at several sites including King William's Town, one of the outbreak sites, and Graaff Reinet. More recent analysis54 has shown that in the Eastern Cape, a summer rainfall area, the coefficient of variation in annual rainfall is typically high, indicating that year-to-year fluctuations in precipitation are likely to vary by up to 40%. Under such conditions it is to be expected that, following De Kock and Wolmarans49, the habitats of B. africanus will dry out making its long-term presence unpredictable. Shiff55 showed how this may happen by demonstrating the effects of the dual stresses of low winter temperatures and drying of the habitat in reducing, or even stopping, reproduction by the closely related B. globosus in a seasonal pool in Zimbabwe. These stresses would also inhibit or retard asexual reproduction by the intramolluscan stages of S. haematobium resulting, in the present situation, in transmission for 3-4 summer months only. Indeed, periodic drying of waterbodies for 3 months or more has often been recommended as a snail (and parasite) control measure. By way of comparison, the ungulate schistosome Schistosoma margrebowiei is only able to survive in seasonal pools in dry areas of West Africa because the similarly brief opportunities for transmission there are compensated for by its egg production being rapid (more so than S. haematobium) and its snail intermediate hosts, Bulinus forskalii and Bulinus scalaris, being better aestivators than B. africanus.48,56
Complicating the understanding of human urogenital schistosomiasis in the region in the late 19th century is the fact that S. haematobium and S. mattheei are sympatric and both use B. africanus as an intermediate host. Additionally, there are the opinions of several authors, including Le Roux57 and Pitchford58, that the drawings of eggs that were recovered by Harley3 from urine samples from patients in Uitenhage, showed mixed infections. The majority of eggs illustrated were S. haematobium but one figure clearly showed S. mattheei. That these mixed schistosome infections were in fact hybrids was demonstrated by Pitchford59 and Wright and Ross.60 With the benefit of hindsight, this finding is not surprising because the two schistosomes share the same snail host and consequently may be transmitted in the same waterbodies when these are used by both humans and domestic stock. These and subsequent studies have shown that (1) a prior S. haematobium infection is necessary for hybrids with S. mattheei to develop in people, (2) the hybrid combination of male S. haematobium x female S. mattheei is particularly robust and is, like S. haematobium, a parasite of the urinary system (the less robust and less common male S. matheei x female S. haematobium hybrid is an intestinal parasite) and (3) the vigour shown by this hybrid is evident in both its larval and adult stages. Wright and Ross60 and Wright and Southgate61 argued, based on experimental data, that hybrid infection rates in B. africanus would be higher and produce more cercariae than pure S. haematobium or S. mattheei infections. They argued that hybrid cercariae were likely to be more infective to people and so hybrid worm loads would be greater than those of pure infections of either species. Hybrid adult worms grow larger than in the initial S. haematobium infections, mature more rapidly and produce more eggs; they are thus likely to be more pathogenic.
Whether this hybrid could have spread under Eastern Cape conditions to the extent that it increased the pathogenicity of the disease is debatable. Following Van Wyk37 and Southgate et al.62, the combination of male S. haematobium and female S. mattheei would give rise to a sizeable population of hybrids in people, but probably at different rates in different places, perhaps replacing the original pure S. haematobium infections in some situations. However, because the study area experiences long, cold winters and is drought prone, snail populations would probably have needed to be reinfected annually in spring - a situation that allows for transmission to people for only 3-4 months of the year. In addition, adult S. mattheei have a much shorter lifespan in humans than do S. haematobium,63 so frequent reinfection in humans by S. mattheei might also be needed to maintain hybridisation. In other words, persistence of hybrids in these outbreaks required the snail intermediate hosts to be reinfected frequently, perhaps annually, and for those involving intense transmission, reinfection of the final hosts as well.
Calculations by Kruger et al.64 indicated, however, that this was unlikely to happen because far fewer hybrid eggs than S. haematobium eggs would be voided into water. Consequently, hybrid worm burdens in people would be too low to produce the pathogenic strain that would be needed to raise the disease's public health profile. They concluded from experimental and field data in the sub-tropical Mpumalanga lowveld that most human S. mattheei infections arose from infections contracted in early spring. They argued that S. mattheei cercariae were more common at this time when temperatures were rising than were S. haematobium cercariae. However, as temperatures in the Eastern Cape do not rise above 20 ºC until October, this pulse of infections may not have occurred there. On balance it seems that the hybrid was not widely spread and there is no evidence that the morbidity here was any more severe than in, for example, KwaZulu-Natal, as discussed earlier.20,21
Pitchford and Geldenhuys29 considered that anthropogenic factors may have been responsible for the retreat of urinary schistosomiasis from the study area. These factors included control measures such as improved living conditions, relocating of houses away from infested streams as was done at Uitenhage, and a prohibition on bathing in the Buffalo River at King William's Town coupled with a chemotherapy campaign in the town. The drug used for chemotherapy was not named but was probably one of several antimony-based compounds available. Typically these had high cure rates but severe side effects so compliance was often problematic. However, R.J. Pitchford thought that increasing agricultural development and the accompanying pollution of waterbodies were important factors (1975, personal communication to CCA, May 10). Changing livestock farming practices dictated by agricultural development may also have upset transmission of the hybrid by separating waterbodies used by stock from those used by people. This separation would have prevented hybridisation by creating a reproductive barrier between the species and result, once again, in only pure S. haematobium infections in people. This separation would in turn have reduced the pathogenicity of human urogenital schistosomiasis infections and lowered the disease's profile in the community.
The discovery of active S. haematobium transmission associated with the Kabeljous River in 2002 represented a new geographical limit for urogenital schistosomiasis in Africa because the locality lies approximately 65 km south-west of the previously accepted limit, the Port Elizabeth-Uitenhage axis, and a few kilometres further south. This new limit also represented a return of the disease to the Eastern Cape area. Schistosomiasis may in fact have been transmitted within the Kabeljous River area before, because parents of the infected children recalled that haematuria had occurred in the area some 30-35 years earlier (i.e. in the late 1960s). This anecdote might explain the delineation of Humansdorp and its environs as an S. haematobium-endemic area by Pitchford and Visser35 (Figure 3). There are also anecdotal reports of haematuria at Kareedouw on the upper reaches of the Kromme River some 70 km further to the west (Van Niekerk J and Kotze M, Department of Health, Humansdorp 2002, personal communication, October 10). Gear et al.30 recorded B. africanus at several localities on the Kromme River near Kareedouw and S. haematobium transmission in this area should be investigated.
The outbreak near Jeffrey's Bay may be a recent example of what happened previously at the historical Eastern Cape hotspots of Port Elizabeth, Uitenhage, King William's Town and East London. Snails were present at these localities and all that was needed was the introduction of the parasite through human contact. The Jeffrey's Bay outbreak was clearly localised and characterised by intense transmission. Its short duration was probably as a result of chemotherapy.
Although various factors would have affected the urinary schistosomiasis transmission cycle at the time, it is likely that the most influential in terms of the decline of the disease between about 1900 and 2002 were those that acted directly on host snail bionomics and cercarial transmission. The area is characterised by unstable, drought-prone habitats with high B. africanus mortality likely during the long, cold winters, which means that the new generation of snails would have needed to be reinfected annually in late spring by the schistosome parasites and this would in turn have led to a short cercarial transmission season. The short lifespan of S. mattheei in human hosts means that, for transmission of the S. haematobium/S. mattheei hybrid to persist, people too would have needed to be reinfected annually. The impact of public health interventions such as the chemotherapy campaign at King William's Town29 would have been more localised.
Acknowledgements
C.C.A. acknowledges discussions with the late Dr R.J. Pitchford (South African Medical Research Council) in the 1970s on the reduction in schistosomiasis transmission in the Eastern Cape. Thanks go to Dr D.O. Coetzee (Superintendent, Humansdorp Hospital) for drawing attention to the 2002 outbreak near Jeffrey's Bay and for hospitality during his visit and to the hospital staff for help in locating Tweekloof dam. Mrs C.E. Archer (University of KwaZulu-Natal) processed the single urine sample sent to Durban. The temperature data for Cape St Francis were kindly supplied by the South African Weather Service, Pretoria.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
C.C.A. was the principal investigator. I.N. contributed to the epidemiological interpretation of the records, mapped the data and formatted the figures.
References
1. World Health Organization (WHO). Statement - WHO working group on urogenital schistosomiasis and HIV transmission, 1-2 October 2009. Geneva: WHO; 2010. [ Links ]
2. Department of Health. Regular treatment of children for soil-transmitted helminth infections and bilharzia - policy and implementation guidelines. Pretoria: South African National Department of Health; 2007. [ Links ]
3. Harley J. On the endemic haematuria of the Cape of Good Hope. Med Chir Trans. 1864;47:55-72. PMid:20896246, PMCid:2147819 [ Links ]
4. Bilharz T. [Further observations on Distomum haematobium which lives in the portal vein of humans and its association with certain pathological manifestations]. Z Wiss Zool. 1852;4:72-76. German. [ Links ]
5. Harley J. A second communication on the endemic haematuria of the Cape of Good Hope and Natal. Med Chir Trans. 1869;52:379-387. PMid:20896346, PMCid:2150512 [ Links ]
6. Harley J. A third communication on the endemic haematuria of the south-eastern coast of Africa, with remarks on the topical medication of the bladder. Med Chir Trans. 1871;54:47-62. PMid:20896377, PMCid:2150475 [ Links ]
7. Chute HM. Intermittent haematuria. S Afr Med J. 1886;(3):38-39. [ Links ]
8. Haw WH. A paper on African haematuria - endemic haematuria - or bilharzia haematobia. S Afr Med J. 1894;(Jan):171-178. [ Links ]
9. Spencer HA. Bilharzia haematobia. S Afr Med J. 1899;6:269-271. [ Links ]
10. Schneider J. Bilharziasis with particular reference to South Africa. Johannesburg: Cape and Transvaal Printers; 1970. [ Links ]
11. Cobbold TS. On the development of Bilharzia haematobia. Br Med J. 1872;2:89-92. http://dx.doi.org/10.1136/bmj.2.604.89, PMCid:2296977 [ Links ]
12. Chute HM. Bilharzia haematobia, Distomum haematobium, endemic haematuria. S Afr Med J. 1888;(10):85-87. [ Links ]
13. Watkins-Pitchford W. Note on schistosomiasis. S Afr Med J. 1915;13:226. [ Links ]
14. Cawston FG. Schistosomiasis in Natal. S Afr Med J. 1916;14:53-54. [ Links ]
15. Hartler EB. Bilharzia haematobia. S Afr Med J. 1887;(5):200-201. [ Links ]
16. Anonymous. Parasitic diseases in South Africa. S Afr Med J. 1898;6:1-5. [ Links ]
17. Haw WH. Review of volume II of Allbutt's system of medicine. S Afr Med J. 1899;7:65. [ Links ]
18. Van der Werf MJ, De Vlas S. Morbidity and infection with schistosomes or soil-transmitted helminths. Report to Parasitic Diseases and Vector Control Division. Geneva: World Health Organization; 2001. [ Links ]
19. Temple Mursell H. Remarks of fulguration for bladder growths. S Afr Med J. 1921;19:25-27. [ Links ]
20. Bhagwandeen SB. The clinico-pathological manifestations of schistosomiasis in the African and the Indian in Durban. Pietermaritzburg: University of Natal Press; 1968. [ Links ]
21. Cooppan RM, Schutte CHJ, Mayet FM, Dingle CE, Van Deventer JMG, Mosese PG. Morbidity from urinary schistosomiasis in relation to intensity of infection in the Natal Province of South Africa. Am J Trop Med Hyg. 1986;35:765-776. PMid:3089036 [ Links ]
22. Turner GA. Bilharziosis in South Africa. Parasitology. 1908;1:195-217. http://dx.doi.org/10.1017/S0031182000003437 [ Links ]
23. Turner GA. Pulmonary bilharziosis in South Africa. Transvaal Med J. 1908;4:107-110. [ Links ]
24. O'Mahoney D, Banach L, La Grange LL. A case of vulval schistosomiasis. S Afr Fam Pract. 2007;49:42-43. [ Links ]
25. Batho E. On endemic haematuria at the Cape of Good Hope and Natal. Br Army Med Rep. 1870;12:502-504. [ Links ]
26. Porter A. Larval Trematoda found in certain South African Mollusca with special reference to schistosomiasis (bilharzia). Publ S Afr Inst Med Res. 1938;62:1-492. [ Links ]
27. Gale GW. Bilharzia (human redwater) disease. Information Brochure 339. Pretoria: Department of Health; 1946. [ Links ]
28. De Meillon B. Aspects of the natural history of bilharzia in South Africa. S Afr Med J. 1948;22:253-260. PMid:18862892 [ Links ]
29. Pitchford R, Geldenhuys PJ. Bilharziasis in the Transkei and Eastern Cape Province. S Afr J Lab Clin Med. 1960;6:237-246. PMid:13736089 [ Links ]
30. Gear JHS, Pitchford RJ, Van Eeden JA. Atlas of bilharzia in South Africa. Johannesburg: South African Institute for Medical Research, South African Medical Research Council and Department of Health; 1980. [ Links ]
31. Pitchford RJ. Report on visit to Transkei, 3-11 September 1981. Unpublished report to the Medical Services, Transkei. Cape Town: South African Medical Research Council; 1981. [ Links ]
32. Mqoqi NP, Dye AH. Schistosoma haematobium in Transkei: A preliminary survey conducted in the Ngqaleni district. S Afr J Sci. 1992;88:445-447. [ Links ]
33. Mqoqi NP, Appleton CC, Dye AH. Prevalence and intensity of Schistosoma haematobium (urinary schistosomiasis) in the Port St Johns district of Transkei. S Afr Med J. 1996;86:76-80. PMid:8685788 [ Links ]
34. Le Roux PL. Notes on the life-cycle of Schistosoma mattheei and observations on the control and eradication of schistosomiasis in man and animals. 15th Ann Rep Dir Vet Ser Union of South Africa. 1929:407-438. [ Links ]
35. Pitchford RJ, Visser PS. The use of behaviour patterns of larval schistosomes in assessing the bilharzia potential of non-endemic areas. S Afr Med J. 1969;43:983-995. PMid:5821211 [ Links ]
36. Veglia F, Le Roux PL. On the morphology of a schistosome (Schistosoma mattheei sp. nov.) from the sheep in the Cape Province. 15th Ann Rep Dir Vet Ser Union of South Africa. 1929:335-346. [ Links ]
37. Van Wyk JA. Transmission of Schistosoma mattheei from animals to man. In: Gear JHS, editor. Medicine in a tropical environment. Proceedings of an international symposium. Pretoria: A.A. Balkema, 1976; p. 705-717. [ Links ]
38. Pitchford RJ. Some aspects of bilharzia in southern Africa. S Afr Med J. 1986;70(suppl):80-82. PMid:3726706 [ Links ]
39. World Health Organization (WHO). Guidelines for the evaluation of soil-transmitted helminthiasis and schistosomiasis at community level. WHO/CTD/SIP/98.1. Geneva: WHO; 1998. [ Links ]
40. Appleton CC, Stiles G. Geology and geomorphology in relation to the distribution of snail intermediate hosts of bilharzia in South Africa. Ann Trop Med Parasitol. 1976;70:189-198. PMid:938124 [ Links ]
41. Connolly M. A monographic survey of the South African non-marine Mollusca. Ann S Afr Mus. 1939;33:1-660. [ Links ]
42. Heeg J. A note on the drastic effects in total dissolved solids on the aquatic pulmonate snail Bulinus (Physopsis) africanus (Krauss). J Limnol Soc S Afr. 1975;1:29-32. [ Links ]
43. Appleton CC. The population fluctuations of five freshwater snail species in the eastern Transvaal lowveld, and their relation to known bilharzia transmission patterns. S Afr J Sci. 1974;70:145-150. [ Links ]
44. De Kock KN. [The population dynamics of five freshwater snail species of medical and veterinary importance under controlled temperature conditions]. DSc thesis, Potchefstroom, Potchefstroom University, 1973. Afrikaans. [ Links ]
45. Pitchford RJ, Visser PS. Some further observations on schistosome transmission in the eastern Transvaal. Bull WHO. 1965;32:83-104. PMid:14292065, PMCid:2555205 [ Links ]
46. Green MS, Swartz T, Mayshar E, et al. When is an epidemic an epidemic? Isr Med Assoc J. 2002;4:3-6. PMid:11802306 [ Links ]
47. Cawston FG. Some effects of a prolonged drought on disease-carriers. J Trop Med Hyg. 1934;37:195-196. [ Links ]
48. Brown DS. Freshwater snails of Africa and their medical importance. Revised 2nd ed. London: Taylor & Francis; 1994. [ Links ]
49. De Kock KN, Wolmarans CT. Distribution and habitats of the Bulinus africanus species group, snail intermediate hosts of Schistosoma haematobium and S. mattheei in South Africa. Water SA. 2005;31:117-125. [ Links ]
50. Pretorius SJ, Joubert PH, De Kock KN. A review of the schistosomiasis risk in South African dams. Water SA. 1989;15:133-136. [ Links ]
51. Cridland CC. Resistance of Bulinus (Physopsis) globosus, Bulinus (Ph.) africanus, Biomphalaria pfeifferi and Lymnaea natalensis to experimental desiccation. Bull WHO. 1967;36:507-513. PMid:5299680, PMCid:2476301 [ Links ]
52. Nicholson SE. The historical climatology of Africa. In: Wigley TML, Ingram MJ, Farmer B, editors. Climate and history: Studies on past climates and their impact on man. Cambridge: Cambridge University Press, 1981; p. 249-270. [ Links ]
53. Nicholson SE. Climate and environmental change in Africa during the last two centuries. Climate Res. 2001;17:123 –144. http://dx.doi.org/10.3354/cr017123 [ Links ]
54. Schulze RE. South African atlas of agrohydrology and climatology. Report TT82/96. Pretoria: Water Research Commission; 1997. [ Links ]
55. Shiff CJ. Studies on Bulinus (Physopsis) globosus in Rhodesia. III-Bionomics of a natural population existing in a temporary habitat. Ann Trop Med Parasitol. 1964;58:240-255. PMid:14183114 [ Links ]
56. Loker ES. A comparative study of the life-histories of mammalian schistosomes. Parasitology. 1983;87:343-369. http://dx.doi.org/10.1017/S0031182000052689 [ Links ]
57. Le Roux PL. The validity of Schistosoma capense (Harley, 1864) amended as a species. Trans R Soc Trop Med Hyg. 1958;52:12-13. [ Links ]
58. Pitchford RJ. Cattle schistosomiasis in man in the eastern Transvaal. Trans R Soc Trop Med Hyg. 1959;53:285-290. http://dx.doi.org/10.1016/0035-9203(59)90010-0 [ Links ]
59. Pitchford RJ. Observations on a possible hybrid between the two schistosomes S. haematobium and S. mattheei. Trans R Soc Trop Med Hyg. 1961;55:44-51. http://dx.doi.org/10.1016/0035-9203(61)90038-4 [ Links ]
60. Wright CA, Ross GC. Hybrids between Schistosoma haematobium and S. mattheei and their identification by isoelectric focusing. Trans R Soc Trop Med Hyg. 1980;74:326-332. http://dx.doi.org/10.1016/0035-9203(80)90091-7 [ Links ]
61. Wright CA, Southgate VR. Hydridisation of schistosomes and some of its implications. In: Taylor AER, Muller R, editors. Genetics aspects of host-parasite relationships. Symposiums of the British Society for Parasitology. 1976;11:91-109. [ Links ]
62. Southgate VR, Tchuem-Tchuente LA, Vercruysse J, Jourdane J. Mating behaviour in mixed infections of Schistosoma haematobium and S. mattheei. Parasitol Res. 1995;81:651-656. http://dx.doi.org/10.1007/BF00931841, PMid:8570579 [ Links ]
63. Pitchford RJ, Visser PS. Excretion of Schistosoma mattheei eggs from man, baboons and cattle living in their natural environment. J Helminthol. 1975;49:137-142. http://dx.doi.org/10.1017/S0022149X00023269 [ Links ]
64. Kruger FJ, Joubert PH, Pretorius SJ. Ratio of Schistosoma haematobium to S. mattheei infections in Bulinus africanus snails from rural areas in the eastern Transvaal lowveld in South Africa. Trans R Soc Trop Med Hyg. 1990;84:556. http://dx.doi.org/10.1016/0035-9203(90)90038-G [ Links ]
 Correspondence to:
Correspondence to:
Chris Appleton
Postal address: School of Biological and Conservation Sciences, University of KwaZulu-Natal, Westville campus
Durban 4000, South Africa
Email: Appletonc@ukzn.ac.za
Received: 20 Aug. 2010
Accepted: 08 July 2011
Published: 09 Jan. 2012
© 2012. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}