










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.11-12 Pretoria Jan. 2011
RESEARCH LETTER
Springtail diversity in South Africa
Charlene JanionI; Anne BedosII; Jan BengtssonIII; Louis DeharvengII; Bettine Jansen van VuurenI,IV; Hans Petter LeinaasV; Amy LiuI; Anna MalmströmIII; David PorcoVI; Steven L. ChownI
ICentre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University, Stellenbosch, South Africa
IIMuseum National d'Histoire Naturelle, Origine, Structure et Evolution de la Biodiversite, Paris, France
IIIDepartment of Ecology, Swedish University of Agricultural Sciences, Uppsala, Sweden
IVMolecular Genomics Group, Department of Botany and Zoology, Stellenbosch University, Stellenbosch, South Africa
VIntegrative Biology Group, Department of Biology, University of Oslo, Oslo, Norway
VILaboratoire ECODIV, UFR Sciences, Université de Rouen, Rouen, France
ABSTRACT
Despite their significance in soil ecosystems and their use for investigations of soil ecosystem functioning and in bioindication elsewhere, springtails (Collembola) have not been well investigated in South Africa. Early recognition of their role in soil systems and sporadic systematic work has essentially characterised knowledge of the southern African fauna for some time. The situation is now changing as a consequence of systematic and ecological work on springtails. To date this research has focused mostly on the Cape Floristic Region and has revealed a much more diverse springtail fauna than previously known (136 identifiable species and an estimated 300 species for the Cape Floristic Region in total), including radiations in genera such as the isotomid Cryptopygus. Quantitative ecological work has shown that alpha diversity can be estimated readily and that the group may be useful for demonstrating land use impacts on soil biodiversity. Moreover, this ecological work has revealed that some disturbed sites, such as those dominated by Galenia africana, may be dominated by invasive springtail species. Investigation of the soil fauna involved in decomposition in Renosterveld and Fynbos has also revealed that biological decomposition has likely been underestimated in these vegetation types, and that the role of fire as the presumed predominant source of nutrient return to the soil may have to be re-examined. Ongoing research on the springtails will provide the information necessary for understanding and conserving soils: one of southern Africa's major natural assets.
Introduction
The significance of soil organisms for ecosystem functioning and ecosystem service delivery is widely appreciated.1,2 Amongst the many arthropod taxa that contribute to soil ecosystem functioning, the springtails (Collembola) have been identified as an important group. These small arthropods occur in most ecosystems and may reach densities of several hundred thousand individuals per square metre.3 They form the prey of a wide variety of soil organisms and ground-living arthropods, actively contribute to soil formation and structure,4 and have major effects on both plants and plant consumers; and thus link the above-ground and below-ground components of terrestrial systems.2,5 The ecological roles and abundance of springtails in terrestrial systems have also led to their recognition as important organisms for bioindication,6,7 and as model organisms in ecotoxicology.8 Consequently, springtails have been the subject of substantial and long-standing interest, especially in Europe and North America.3,9,10
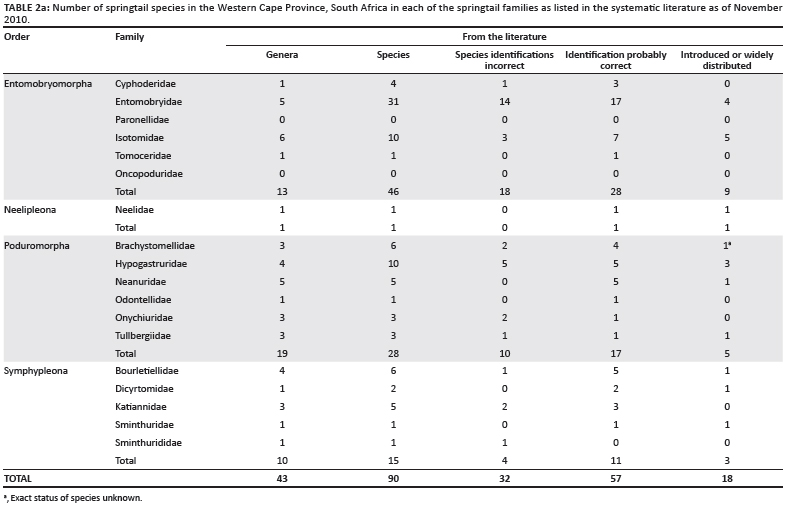
The significance of springtails in soil systems in South Africa is unlikely to be different from that elsewhere, recognising that springtails reach their highest species richness and abundance in moist habitats rather than in arid areas.11,12 However, in contrast with many other regions of the world,10 and with other components of the soil fauna in South Africa, such as the ants,13,14,15,16 springtails have, until recently, been the subject of little attention. Early research recognised that springtails are likely to play an important role in various South African biomes,17 and since the initial work of Womersley18, ongoing, but sporadic and typically restricted taxonomic investigations have been made.19,20,21,22,23,24,25,26,27,28 Few investigations have sought to understand comprehensively the diversity of the group (at present the only available, but unpublished list by P. Greenslade, places the fauna at about 43 genera and 90 species for the Western Cape), their contribution to ecosystem functioning, and their utility for bioindication,29 although occasional attempts at doing so have been made.30 The exception is research on the group undertaken in the sub-Antarctic Tundra and Polar Desert biomes of the Prince Edward Islands, which, at least geopolitically, form part of South Africa.31 For these island biomes, the limited fauna of 16 species is relatively well understood.32,33,34,35,36 Perhaps more significantly, it is this island work that has precipitated a recent and substantial change in current understanding of springtail ecology and systematics in continental South Africa.
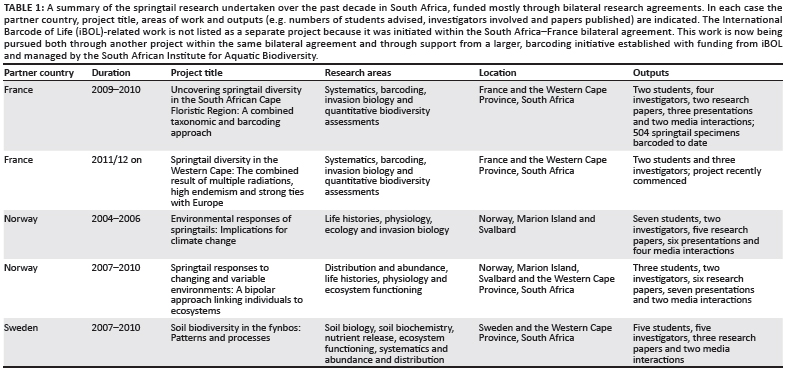
On the basis of a comprehensive comparison of the likely effects of climate change on indigenous versus invasive springtails on the Prince Edward Islands, funded by a bilateral grant under the South Africa-Norway agreement (Table 1),34 a second project was developed to compare life histories of species in polar systems (namely, the Prince Edward Islands and Svalbard, Norway) and more temperate systems (southern Norway and the Western Cape Province, South Africa). This work sought also to investigate the implications thereof for ecosystem functioning and drew in further expertise from Sweden and support from a South Africa- Sweden bilateral (Table 1). In both cases, it was recognised from the outset that the springtails in South Africa are poorly known systematically, and that this taxonomic impediment37 would therefore constrain work substantially. Thus, additional collaborations were established with systematic experts under a third bilateral agreement (South Africa- France) and with the support of the International Barcode of Life project (iBOL - http://ibol.org) (Table 1). Here we report on research so far undertaken across these major projects, how systematic and ecological understanding of the group is progressing for South Africa, and the prospects for ongoing research and future collaborations amongst researchers with an interest in soil diversity and ecosystem functioning.
Systematics and barcoding
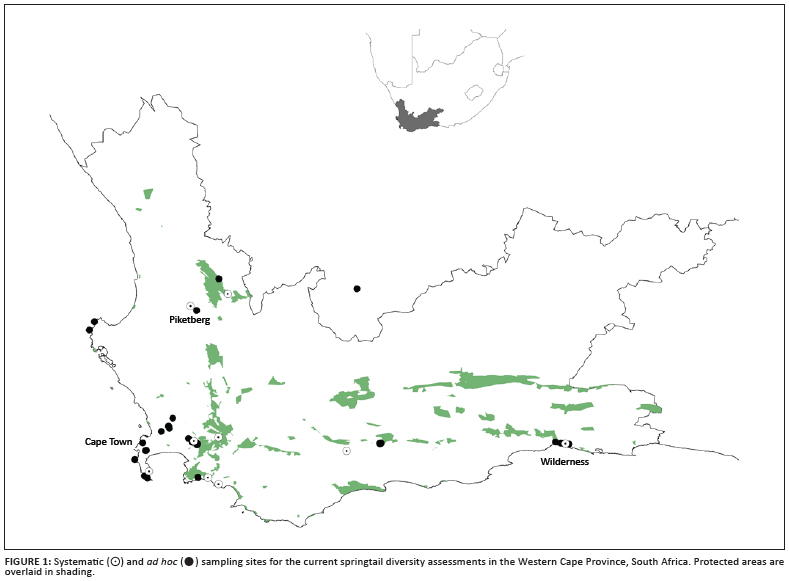
Owing to estimates of the likely large size of the South African springtail fauna, the collaborative research was established to investigate the diversity of the Cape Floristic Region, focusing mostly on fynbos, renosterveld and forest vegetation. From the outset, the research approach emphasised collection across a range of sites in a systematic fashion (Figure 1) for estimates of richness and abundance, supplemented by ad hoc collections from as wide a range of habitats as possible for the systematic and barcoding studies. Importantly, our investigations combined traditional taxonomy and barcoding to ensure an integrated, modern approach to the systematics of the group.
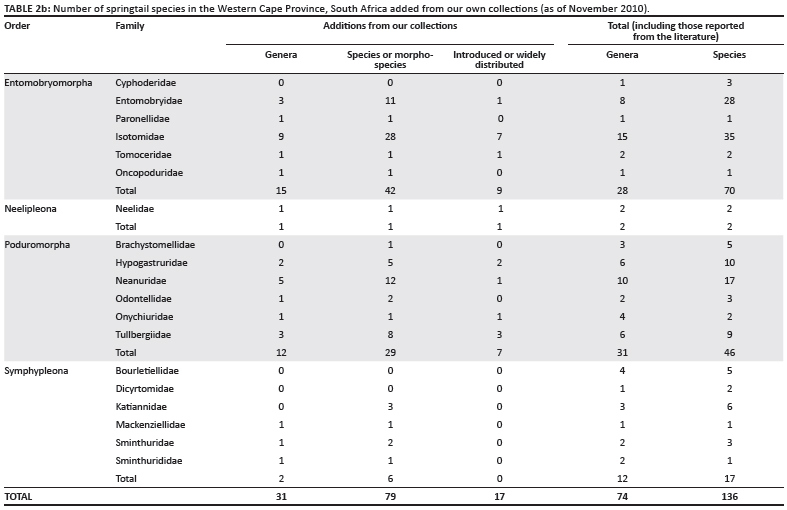
Initial assessments of the published taxonomic research indicated a faunal assortment of 90 species in 16 families, but, excluding incorrect identifications, only 57 valid and recognisable species (Table 2a). Now, based on a geographically extensive collection of more than 450 samples from the Western Cape (Figure 1), it is clear that the faunal component is much larger. The collection methods included high gradient extraction of litter bags; Berlese-Tullgren extraction of leaf litter, moss and rotten wood; vegetation beating and collection by hand (especially in caves and streams); pitfall trapping; litter sifting; and soil washing (of beach and deep soil sand). The current faunal assemblage stands at 136 morphospecies in 19 families (Table 2b), but only half of the samples have been sorted to species so far and not all microhabitats have been surveyed to completion. Moreover, assessments of previous lists (in particular Paclt19) for the country, the examination of museum specimens and barcoding work have indicated that many species have been misidentified and that several groups have either cryptic species or have shown substantial radiations. For example, more than 11 Cryptopygus and 7 Parisotoma species were found during our survey, in contrast to only 2 species that have been previously recorded - Cryptopygus caecus and Parisotoma notabilis.19 In addition, 23 described species of Seira are found in South Africa,22 whilst only 1 species is described from Australia.38 These groups that have radiated in the Cape Floristic Region are now the subject of closer investigation from the classical taxonomic39,40 and barcoding perspectives. Thus, although research to date has more than doubled the number of known species, based on our experience from elsewhere it seems likely that at least 300 species will be characteristic of the regional (i.e. Cape Floristic Region) fauna. For South Africa as a whole the richness is certainly much higher. Even so, the Collembola likely has fewer species
than are found in the highly diverse insect orders, such as the Coleoptera or Lepidoptera.41 Many sequences obtained from barcoding have been connected to traditional taxonomic species, although certain species with poor morphological descriptions have been more difficult (as in the case of several Seira species). The barcoding results have also led to careful re-examination of the individuals traditionally assigned to the widespread species Parisotoma notabilis, which actually represent a group of several similar species of endemic Parisotoma. In addition, the barcoding has proven not only to be of particular importance to detect introduced species of complex groups (e.g. Isotomurus maculatus), but also to show that some forms expected to be European actually do not match any European cluster (such as one cluster of Isotomurus and several of Ceratophysella).
Invasions and diversity estimation
Amongst the 136 species-level taxa identified for the Western Cape, at least 34 represent species either with wide distributions or known or suspected to have been introduced from elsewhere. Based on our preliminary molecular analysis, two of these species – Neanura muscorum and Isotomurus cf. maculatus – have already been shown to be invasive, occurring locally in fynbos or forest habitat. Similar comparisons between South African and European populations of other suspected invasive species are in progress. For example, in an investigation of diversity and decomposition rates in renosterveld fragments in the Piketberg area,42 the springtail assemblages were dominated (60% by abundance) by a single species, Hypogastrura manubrialis, which is widely distributed in Europe,43 and clearly is an invasive species in the Western Cape. Similarly, two Isotomurus species, thought to be introduced to the region, were also found in high abundance in a preliminary assessment of the diversity of springtails in Cape Flats Sand Fynbos and an adjacent pine plantation in the Tokai Forest Reserve, which forms part of Table Mountain National Park; these species were restricted to the pine plantation. This study also showed that springtail abundance and species richness (but not species identity) differ significantly amongst these two major habitat types, and, that for complete estimation of the fauna using a litter sampling technique (for each sample: 1 L of litter collected over a standardised 1 m2 and then extracted by a Berlese– Tullgren funnel in the laboratory), the extent of sampling is not so onerous that it precludes reasonably straightforward estimation of local (alpha) diversity (Figure 2). In consequence, the springtails could be used for assessments of soil health, as they are elsewhere.3,9
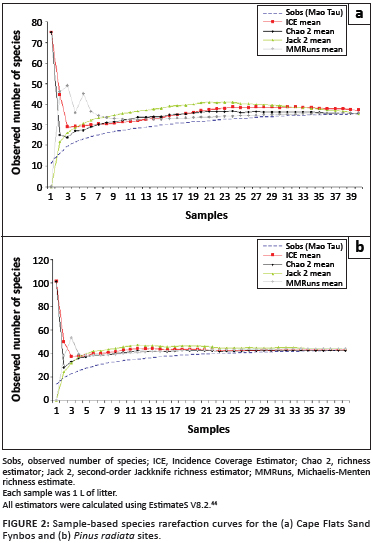
Ecosystem functioning
In the context of soil health, the influence of springtails on litter decomposition rates and nutrient cycling, including the ways in which litter type, spatial position and home-field advantage influence decomposition, has long been of interest to soil ecologists.45,46,47,48,49 As a consequence of the research priorities identified within our South Africa- Sweden and South Africa-Norway bilateral projects (Table 1), we have also been concerned with these questions. The dominant paradigm for the Fynbos biome has been that loss of organic matter and nutrient cycling take place largely as a consequence of fires, which, with an average fire frequency of 11 years,50 return nutrients accumulated in litter to the soil. Biological decomposition was largely relegated to a less significant role.51,52,53
During the investigation of springtail diversity in renosterveld fragments, decomposition of the litter of three representative plant species was also investigated. Decomposition of these species varied between 0.00674/day for Galenia africana, a shrub favoured by disturbances such as overgrazing, to 0.00222/day for renosterbos Elytropappus rhinocerotis and 0.00029/day for the sturdy geophyte Watsonia borbonica, corresponding to litter half-lives of 0.3, 0.8 and 6 years, respectively.42 The rates for Galenia and renosterbos are much faster than those previously found for litter of the fynbos species Leucadendron parile and Protea repens.51,52 However, they are not unusual compared with other fynbos species we have studied (Figure 3; personal observation).
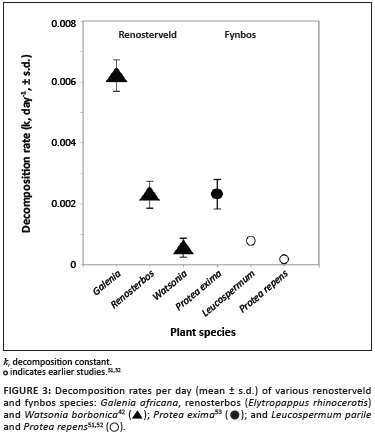
If the mean fire return time for fynbos systems is about 11 years,50 and if decomposition of the less hardy species proceeds such that many litters have half-lives that are less than half this time, then biological decomposition as a nutrient recycling process is much more significant than previously estimated.53 Ultimately, the significance of this decomposition will depend on the relative contributions of species with more readily or less readily decomposed litter to the litter pool of any given system. To date, such estimates have not been made. Species with high decomposition rates of the kinds we investigated clearly make a significant contribution to the fynbos flora, and it is obvious that litter of many species do not accumulate on the ground in the way that some Protea species do (personal observations). Thus, the current research on ecosystem functioning has demonstrated that important as fire is in the Fynbos biome, it may be complemented and sometimes surpassed by other processes.
Conclusions
Our research on springtails has thus far revealed not only a hitherto undocumented diversity in South Africa, but also that the group may be much more significant for ecosystem functioning than previously thought. Given a growing human population and its impacts on the environment,54 national requirements for sustainable development and conservation, and the need to provide measures of conservation and sustainable development success internationally, ongoing work on the Collembola will prove both useful and valuable. The current collaborations described here will continue throughout the duration of the International Barcode of Life initiative (iBOL - http://ibol.org), with support from several institutions, both in South Africa and abroad, and growing interest from various sectors in demonstrating soil health and conservation success. Moreover, as this knowledge develops so more information will be made available through the project home page ( www.sun.ac.za/cib/collembola ). Our long-term aims are to encourage additional work on the group in the southern African region, and to foster collaborations that can enhance understanding of this significant group of organisms. Underlying these aims is the realisation that sustainable development and conservation must continue to focus on one of southern Africa's major natural assets: the soil.
Acknowledgements
We are grateful for support, through the National Research Foundation, from the South Africa-France, South Africa- Norway and South Africa-Sweden bilateral grants, and the International Barcode of Life project. We thank Melodie McGeoch, Brigitte Brashler and two anonymous reviewers for comments on a previous version of the manuscript. The map was kindly produced by Dian Spear. Cape Nature and SANParks provided collection permits.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
S.L.C. was the project leader; A.B., L.D., H.P.L. and C.J. provided taxonomic data; H.P.L., J.B., A.M. and C.J. provided decomposition data; A.L. provided rarefaction curves; and D.P. and B.J.V.V. provided data on barcoding.
References
1. Wardle DA. Communities and ecosystems: Linking the aboveground and belowground components. Princeton, NJ: Princeton University Press; 2002. [ Links ]
2. Wardle DA, Bardgett RD, Klironomos JN, Setälä H, Van der Putten WH, Wall DH. Ecological linkages between aboveground and belowground biota. Science. 2004;304:1629-1633. http://dx.doi.org/10.1126/science.1094875, PMid:15192218 [ Links ]
3. Hopkin S. Biology of the springtails. Insecta: Collembola. Oxford: Oxford University Press; 1997. [ Links ]
4. Rusek J. Soil microstructures-contributions on specific soil organisms. Quest Ent. 1985;21:497-514. [ Links ]
5. Scheu S, Treenhaus A, Jones TH. Links between the detritivore and the herbivore system: Effects of earthworms and Collembola on plant growth and aphid development. Oecologia. 1999;119:541-551. http://dx.doi. org/10.1007/s004420050817 [ Links ]
6. Deharveng L. Soil Collembola diversity, endemism, and reforestation: A case study in the Pyrenees (France). Conserv Biol. 1996;10:74-84. http://dx.doi.org/10.1046/j.1523-1739.1996.10010074.x [ Links ]
7. Cassagne N, Gauquelin T, Bal-Serin M-C, Gers C. Endemic Collembola, privileged bioindicators of forest management. Pedobiologia. 2006;50:127-134. http://dx.doi.org/10.1016/j.pedobi.2005.10.002 [ Links ]
8. Van Straalen NM, Van Gestel CAM. Soil invertebrates and microorganisms. In: Calow P, editor. Handbook of ecotoxicology. Oxford: Blackwell; 2009. [ Links ]
9.Rusek J. Biodiversity of Collembola and their functional role in the ecosystem. Biodiv Conserv. 1998;7:1207-1219. http://dx.doi.org/10.1023/A:1008887817883 [ Links ]
10. Deharveng L. Recent advances in Collembola systematics. Pedobiologia. 2004;48:415-433. http://dx.doi.org/10.1016/j.pedobi.2004.08.001 [ Links ]
11. Petersen H, Luxton M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos. 1982;39:287-388. http://dx.doi.org/10.2307/3544689 [ Links ]
12. Ulrich W, Fiera C. Environmental correlates of species richness of European springtails (Hexapoda: Collembola). Acta Oecol. 2009;35:45-52. http://dx.doi.org/10.1016/j.actao.2008.07.007 [ Links ]
13. Robertson HG. Afrotropical ants (Hymenoptera: Formicidae): Taxonomic progress and estimation of species richness. J Hymen Res. 2000;9:74-84. [ Links ]
14. Parr CL, Robertson HG, Biggs HC, Chown SL. Response of African savanna ants to long-term fire regimes. J Appl Ecol. 2004;41:630-642. http://dx.doi. org/10.1111/j.0021-8901.2004.00920.x [ Links ]
15. Botes A, McGeoch MA, Robertson HG, Van Niekerk A, Davids HP, Chown SL. Ants, altitude and change in the northern Cape Floristic Region. J Biogeogr. 2006;33:71-90. http://dx.doi.org/10.1111/j.13652699.2005.01336.x [ Links ]
16. Braschler B, Kirsten M, Karenyi N, Gaston KJ, Chown SL. Realizing a synergy between research and education: How participation in ant monitoring helps raise biodiversity awareness in a resource-poor country. J Insect Cons. 2010;14:19-30. http://dx.doi.org/10.1007/s10841-009-9221-6 [ Links ]
17. Lawrence RF. The biology of the cryptic fauna of forests, with special reference to the indigenous forests of South Africa. Cape Town: Balkema; 1953. [ Links ]
18. Womersley H. On some Collembola Arthropleona from South Africa and southern Rhodesia. Ann S Afr Mus. 1934;30:441-475. [ Links ]
19. Paclt J. Collembola. In: Hanstrom B, Brinck P, Rudebeck G, editors. South African animal life. Results of the Lund University Expedition in 1950- 1951. Vol. VI. Uppsala: Almqvist & Wiksells Boktryckeri AB, 1959; p. 24-78. [ Links ]
20. Yosii R. Collembola fauna of the Cape Province, with special reference to the genus Seira Lubbock. In: Biological results of the Japanese Antarctic Research Expedition 6. Kyoto: Siraham, Wakayam-Ken, 1959; p. 1-23. [ Links ]
21. Coates TJ. The Collembola of South Africa - 1: The genus Neanura. J Ent Soc Sth Afr. 1968;31:185-195. [ Links ]
22. Coates TJ. The Collembola of South Africa - 2: The genus Seira. J Ent Soc Sth Afr. 1968;31:435-462. [ Links ]
23. Coates TJ. The Collembola of South Africa - 3: The genus Lepidokrugeria. J Ent Soc Sth Afr. 1969;32:87-89. [ Links ]
24. Coates TJ. Check-list of the Collembola of South African Parks (Part 1). Koedoe. 1970;13:181-184. [ Links ]
25. Weiner WM, Najt J. Collembola Poduromorpha of South Africa. Bonn Zool Beitr. 1991;42:369-387. [ Links ]
26. Barra J-A. Nouveaux Collemboles Poduromorphes de la Province du Natal (Rép. Sud Africaine) (Insecta: Collembola). [New Poduromorpha Collembola from Natal Province (Republic of South Africa) (Insecta: Collembola)]. J Afr Zool. 1994;108:181-189. French. [ Links ]
27. Barra J-A. Nouveaux Collemboles Poduromorphes des sables littoraux (partie terrestre) de la Province du Natal (Rép. Sud Africaine) (Insecta : Collembola). [New Poduromorpha Collembola from the sand coasts (terrestrial part) from Natal Province (Republic of South Africa) (Insecta: Collembola)]. J Afr Zool. 1995;109:125-139. French. [ Links ]
28. Barra J-A. Nouveaux Collemboles Entomobryomorphes des sables littoraux (partie terrestre) de la Province du Natal (Rép. Sud Africaine) (Insecta : Collembola). [New Entomobryomorpha Collembola from the sand coasts (terrestrial part) of the Natal Province (Republic of South Africa) (Insecta : Collembola)]. J Afr Zool. 1997;111:465-480. French. [ Links ]
29. McGeoch MA. The selection, testing and application of terrestrial insects as bioindicators. Biol Rev. 1998;73:181-201. http://dx.doi.org/10.1017/S000632319700515X [ Links ]
30. Reinecke AJ, Albertus RMC, Reinecke SA, Larink O. The effects of organic and conventional management practices on feeding activity of soil organisms in vineyards. Afr Zool. 2008;43:66-74. http://dx.doi. org/10.3377/1562-7020(2008)43[66:TEOOAC]2.0.CO;2 [ Links ]
31. Chown SL, Froneman PW. The Prince Edward Islands. Land-sea interactions in a changing climate. Stellenbosch: African Sun Media; 2008. [ Links ]
32. Deharveng L. Collemboles des iles subantarctiques de l'Océan Indien Mission J. Travé 1972-1973. [Collembola of the sub-Antarctic islands of the Indian Ocean Mission J. Travé 1972-1973]. Com Nat Fr Rech Antarct. 1981;48:33-108. French. [ Links ]
33. Gabriel AGA, Chown SL, Barendse J, et al. Biological invasions on Southern Ocean islands: The Collembola of Marion Island as a test of generalities. Ecography. 2001;24:421-430. http://dx.doi.org/10.1034/j.1600-0587.2001.d01-198.x [ Links ]
34. Chown SL, Slabber S, McGeoch MA, Janion C, Leinaas HP. Phenotypic plasticity mediates climate change responses among invasive and indigenous arthropods. Proc R Soc Lond B. 2007;274:2661-2667. http://dx.doi.org/10.1098/rspb.2007.0772, PMid:17686728, PMCid:2275886 [ Links ]
35. Myburgh M, Chown SL, Daniels SR, Jansen van Vuuren B. Population structure, propagule pressure, and conservation biogeography in the sub-Antarctic: Lessons from indigenous and invasive springtails. Divers Distrib. 2007;13:143-154. [ Links ]
36. Janion C, Leinaas H-P, Terblanche JS, Chown SL. Trait means and reaction norms: The consequences of climate change/invasion interactions at the organism level. Evol Ecol. 2010;24:1365-1380. http://dx.doi.org/10.1007/s10682-010-9405-2 [ Links ]
37. Samways MJ. Insect conservation biology. London: Chapman & Hall; 1994. [ Links ]
38. Greenslade P. Collembola. In: Houston VWK, editor. Zoological catalogue of Australia. Volume 22. Protura, Collembola, Diplura. Melbourne: CSIRO, 1994; p. 19-138. [ Links ]
39. Potapov M, Janion C, Deharveng L. Two new species of Parisotoma (Collembola: Isotomidae) from the Western Cape, South Africa. Zootaxa. 2011;2771:17-24. PMid:21547000, PMCid:3086760 [ Links ]
40. Janion C, Bedos A, Deharveng L. The genus Ectonura, Cassagnau 1980 in South Africa (Collembola: Neanuridae: Neanurinae), with a key to South African Neanurinae. ZooKeys. 2011;136:31-45. [ Links ]
41. Scholtz CH, Chown SL. Insects in southern Africa: How many species are there? S Afr J Sci. 1995;91:124-126. [ Links ]
42. Bengtsson J, Janion C, Chown SL, Leinaas HP. Variation in decomposition rates in the fynbos biome, South Africa: The role of plant species and plant stoichiometry. Oecologia. 2011;165:225-235. http://dx.doi.org/10.1007/s00442-010-1753-7, PMid:20827492, PMCid:3015188 [ Links ]
43. Fjellberg A. The Collembola of Fennoscandia and Denmark. Part I: Poduromorpha. Fauna Entomologica Scandinavica 35. Leiden: Brill; 1998. [ Links ]
44. Colwell RK. EstimateS: Statistical estimation of species richness and shared species from samples (software and user's guide). Version 8.2.0. Storrs, CT: University of Connecticut; 2009. [ Links ]
45. Seastedt TR. The role of microarthropods in decomposition and mineralization processes. Annu Rev Entomol. 1984;29:25-46. [ Links ]
46. Verhoef H, Brussaard L. Decomposition and nitrogen mineralization in natural and agro-ecosystems: The contribution of soil animals. Biogeochemistry. 1990;11:175-211. http://dx.doi.org/10.1007/BF00004496 [ Links ]
47. Barrios E. Soil biota, ecosystem services and land productivity. Ecol Econ. 2007;64:269-285. [ Links ]
48. Ettema CH, Wardle DA. Spatial soil ecology. Trends Ecol Evol. 2002;17:177-183. http://dx.doi.org/10.1016/S0169-5347(02)02496-5 [ Links ]
49. Ayres E, Steltzer H, Simmons BL, et al. Home-field advantage accelerates leaf litter decomposition in forests. Soil Biol Biochem. 2009;41:606-610. [ Links ]
50. Van Wilgen BW. The evolution of fire and invasive alien plant management practices in fynbos. S Afr J Sci. 2009;105:335-342. [ Links ]
51. Mitchell DT, Coley PGF, Webb S, Allsopp N. Litter fall and decomposition processes in the coastal fynbos vegetation, south-western Cape, South Africa. J Ecol. 1986;74:977-993. [ Links ]
52. Mitchell DT, Coley PGF. Litter production and decomposition from shrubs of Protea repens growing in sand plain lowland and mountain fynbos, south-western Cape. S Afr J Bot. 1987;53:25-31. [ Links ]
53. Stock WD, Allsopp N. Functional perspective of ecosystems. In: Cowling RM, editor. The ecology of fynbos: Nutrients, fire and diversity. Cape Town: Oxford University Press, 1992; p. 241-259. [ Links ]
54. Chown SL. Temporal biodiversity change in transformed landscapes: A southern African perspective. Phil Trans R Soc B. 2010;365:3729-3742. [ Links ]
 Correspondence to:
Correspondence to:
Charlene Janion
Postal address:
Private Bag X1
Matieland 7602, South Africa
Email: cjanion@sun.ac.za
Received: 10 Jan. 2011
Accepted: 09 Sept. 2011
Published: 07 Nov. 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}