










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.11-12 Pretoria ene. 2011
RESEARCH LETTER
3D techniques and fossil identification: an elephant shrew hemi-mandible from the Malapa site
Aurore ValI; Kristian J. CarlsonII; Christine SteiningerII; Job M. KibiiII; Cecil ChurmsIII; Brian F. KuhnII; Lee R. BergerII
IBernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Johannesburg, South Africa
IIInstitute for Human Evolution, University of the Witwatersrand, Johannesburg, South Africa
IIIDeb Tech, De Beers Group Services, Johannesburg, South Africa
ABSTRACT
Conventional methods for extracting fossilised bones from calcified clastic sediments, using air drills or chemical preparations, can damage specimens to the point of rendering them unidentifiable. As an alternative, we tested an in silico approach that extended preparation and identification possibilities beyond those realisable using physical methods, ultimately proving to be crucial in identifying a fragile fossil. Image data from a matrix-encased hemi-mandible of a micromammal that was collected from the Plio-Pleistocene site of Malapa, Cradle of Humankind, South Africa, were acquired using microtomography. From the resultant images, a 3D rendering of the fossil was digitally segmented. Diagnostic morphologies were evaluated on the rendering for comparison with extant comparative specimens, positively identifying the specimen as an elephant shrew (Elephantulus sp.). This specimen is the first positively identified micromammal in the Malapa faunal assemblage. Cutting-edge in silico preparation technology provides a novel tool for identifying fossils without endangering bone integrity, as is commonly risked with physical preparation.
Introduction
The recently discovered site of Malapa has yielded a new hominin, Australopithecus sediba, in association with other faunal remains.1,2 The site is situated in the Cradle of Humankind, South Africa, an area that hosts more than a dozen Plio-Pleistocene hominin-bearing cave deposits.3,4,5,6 Age estimates have been obtained using different techniques that coalesce at 1.977 Ma.7 The majority of the faunal remains in the Malapa assemblage, as with other sites from the Sterkfontein Valley, are embedded in dense matrix (e.g. calcified clastic sediments).2,8 Amidst the more than 300 non-hominin remains that have been recovered so far,2 there is a well-preserved left hemi-mandible of a micromammal (specimen UW88-1100) with the third and fourth premolars, as well as the first and second molars (Figure 1). At present, no micromammals have been positively identified in the Malapa faunal assemblage.2 The calcified clastic sediment block (UW88-B738) containing the hemi-mandible was displaced by miners prior to the discovery of the site, making it difficult to re-establish its precise facies of origin. However, according to dates obtained from the presently known depositional sequence at Malapa, the hemi-mandible is likely to have been deposited in a single debris flow around 1.977 Ma.7
Following standard protocols for the preparation of Malapa fossils, the lingual side of the specimen was exposed initially using an air scribe, revealing a hemi-mandible that is missing the distal part of the angular process and the entire mandibular body anterior to the P3 (Figure 1).
Qualitative and quantitative observations of mandibular morphology and dental morphology were sufficient to identify the specimen as insectivorous but taxonomic identification below the level of Order was not possible without verifying the number and morphology of tooth cusps.9,10 However, unobstructed observation of mandibular morphology and tooth occlusal surfaces was problematic because of the matrix still adhering to the specimen. Visible cracks riddling the surface of the specimen effectively ruled out further physical or chemical preparation, leaving positive taxonomic identification unobtainable. Therefore, we applied virtual extraction and reconstruction techniques in order to advance the taxonomic diagnosis beyond a state reached through traditional preparation techniques.
Methods
Initial physical preparation of the block was performed by a preparator (Roseberry Languza) using a small diamond-tipped electrical drill. This initial preparation revealed the extent of the specimen within the block. After exposing the lingual surface of the hemi-mandible by removing a few cubic centimetres of matrix, it became obvious that further preparation (e.g. exposing tooth crowns) would irreparably damage the specimen, meaning that physical extraction from the matrix was too risky. Hence, in order to continue with taxonomic identification, we decided to scan the entire block containing the specimen at high resolution (approximately 10 mm) using an X-Tek 225 HMX microfocus CT scanner (Nikon Metrology NV, Leuven, Belgium). We used the resulting serial .tif stack to segment the fossil specimen and produce a 3D rendering (Figure 2). The 3D rendering of the specimen was done by importing the stack of images into Avizo® 6.211 using the Label Fields module to partition bone or enamel from the surrounding matrix, and then render a surface from the labels. Thus, in completing preparation of the specimen, we adopted a digital extraction method rather than a physical extraction method, with only the former providing an opportunity for a classic comparative palaeontological study.

Results
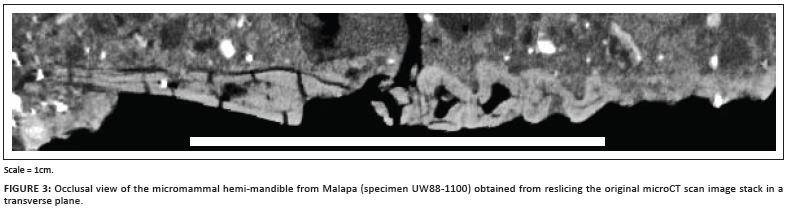
The 3D rendering provided an opportunity to characterise features of the dentition and the mandibular shape (Figure 3) that would not have been possible using traditional preparation techniques. The most diagnostic of these features are the sharpness of the cusps, teeth with higher lingual sides than buccal sides and the orientation of molars and premolars along the same axis. According to the qualitative characteristics of the hemi-mandible and tooth morphology, as well as the dimensions visible on the 3D model (Table 1), we identified the hemi-mandible as belonging to the Family Macroscelidae (the elephant shrew).10
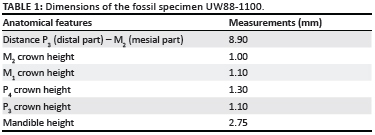
Six extant species of elephant shrew inhabit areas from the west to the east coast of southern Africa (Namibia, Botswana, South Africa and Mozambique).10 Each of these species also is known from Plio-Pleistocene fossil assemblages of South Africa.12,13,14 Two additional extinct species, Elephantulus antiquus and Elephantulus broomi, are present in Plio-Pleistocene fossil assemblages of South Africa.12,13
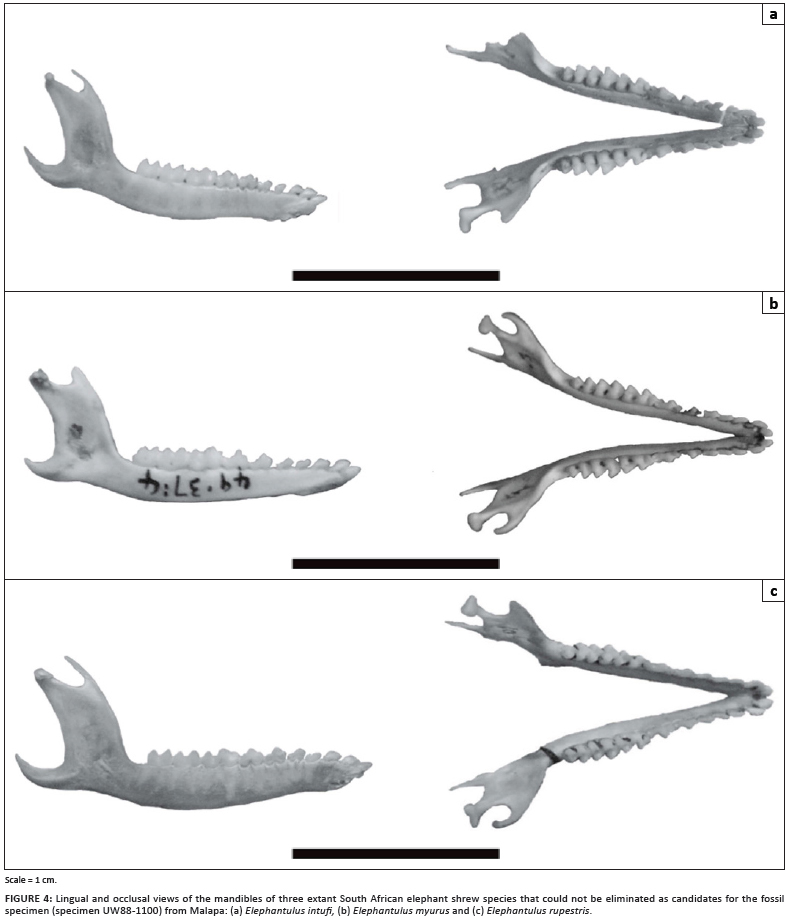
Two of the extant species found in Plio-Pleistocene South African cave sites (i.e. Elephantulus fuscus and Elephantulus brachyrhynchus) were ruled out for the Malapa specimen because they possess a third lower molar, whereas the specimen from Malapa lacks this tooth. Several dental features identify Elephantulus to species, notably the number of cusps on the incisors and on the upper premolars.9,10 Although cuspal patterns can be adequately visualised on the rendering, the hemi-mandible from Malapa (UW88-1100) does not retain the more taxonomically diagnostic lower incisors, upper dentition and maxilla. It is therefore imprudent to rule out any of the four extant taxa (i.e. Elephantulus edwardii, Elephantulus myurus, Elephantulus rupestris and Elephantulus intufi; Figure 4), or the two South African extinct species (E. antiquus and E. broomi) on the basis of the hemi-mandible alone, leading us to provisionally assign this hemi-mandible to Elephantulus sp.
Discussion
Advantages of the new method
Incorporation of microCT into fossil preparation techniques, together with the opportunity to acquire digital measurements using the in silico approach, provided several unique advantages over traditional techniques in this case. Firstly, continuing traditional preparation of the occlusal surfaces in the tiny fossil likely would have destroyed informative morphology because of the delicate condition of the specimen. Secondly, physical preparation, whether using an air drill or applying chemicals, can be quite time-consuming. MicroCT scanning of the fossil required only about an hour; acquiring a stack of .tif or DICOM images was accomplished in less time. Thirdly, once the fossil had been scanned, potentially risky handling of the damaged specimen was drastically reduced. Moreover, the importance of reduced handling was not fully appreciated until the condition of the specimen (i.e. the presence of abundant microcracks) was fully illuminated through microCT scanning. Macrocracks on the external surface (on the lingual side previously exposed) were visible with the naked eye, but only after virtual images were taken and the rendering was obtained, was it documented that microcracks were actually running throughout the whole hemi-mandible.
From the 3D rendering, suitable images for conducting comparative studies were generated. These studies included examination of internal dimensions via virtual sectioning, as well as the ability to manipulate the rendering of the specimen freely in all three dimensions. It also was possible to take repeatable measurements on the rendering (to an accuracy of sub-100 micron), which could be used in comparisons with measurements on other free specimens. None of these data would have been obtainable on the actual specimen, unless destructive methods were employed (i.e. the continuation of physical preparation, sectioning, etc.).
Lastly, commercial visualisation software (e.g. Avizo 6.2, VGStudio Max) provides the ability to zoom in, which in this case allowed observation of finer details on the specimen through magnification. Such software also permits the digital extraction of a specific structure (e.g. individual or multiple teeth; Figure 5) from which a 3D printout can be produced. The 3D printout can be transported and distributed for use, or can even serve as the basis for producing a mould of the fossil in order to distribute casts.

Research perspectives
Diagenesis in the caves of the Sterkfontein Valley often results in bone remains being preserved within dense matrix such as calcified clastic sediments.3,8 Image data acquisition using microtomography, or even medical computed tomography, coupled with 3D reconstruction and extraction of specimens, facilitates exploration inside matrix blocks without requiring substantial manual preparation, which itself is associated with financial and time costs.
In this particular case, the specimen was revealed on the surface of the block, after initial physical preparation. The in silico method allowed us to explore the extent of the fossil partially hidden inside the block. This method also permitted virtual identification, measurements and 3D extraction of the specimen whilst it was still inside the block, and without risking further damage to the specimen. As demonstrated in the case of the Malapa fossil (UW88-1100), this approach permits the prescreening of matrix blocks in order to apply cost-benefit analyses for deciding whether to pursue further physical preparation by taking into consideration the value of the informational content of fossils imprisoned within matrix blocks.
Some microfauna species are excellent indicators of the localised environmental conditions such as climate and landscape.14 Therefore, identification of microfaunal remains is a useful tool during the assembly of palaeoclimatic and palaeoenvironmental reconstructions.14 The elephant shrew in southern Africa typically occupies an arid environment, from desert to dry savanna or dry shrubland, but resides in close proximity to a water source.10 Three species, E. rupestris, E. myurus and E. edwardii are associated with rocky ridges and outcrops, or koppies, whereas E. intufi occupies burrows under bushes and is indicative of regions characterised by scrub bush, thin grass cover and sandy substrate.10 Further additions to the Malapa micromammal assemblage will be informative to the nature of the Malapa palaeoenvironment, perhaps narrowing the present characterisation even more.
Because of the geological context of cave sites in the Cradle of Humankind, it is a long and laborious process to identify the presence of small-bodied fauna, to extract them from the encasing matrix and to identify them to the level of species. The application of microCT scanning and in silico preparation techniques will more easily facilitate each of these steps, particularly for microfauna, by minimising (1) the amount of preparation damage to specimens and (2) the misallocation of valuable manual preparation time and effort on specimens yielding comparatively little new information.
Conclusions
Incorporating microCT scanning and segmentation of renderings into palaeontological preparation methods offers an alternative that can limit damage to valuable fossils and reduce time-consuming manual preparation and physical extraction of fossils from matrix. The use of 3D renderings of fossils enhances an opportunity to identify and describe fossil remains that otherwise would be limited when using traditional physical preparation techniques. This method can be particularly advantageous when studying small, damaged or delicate specimens. It also can be useful in other scenarios, including in the analysis of larger taxa from Plio-Pleistocene sites from the Cradle of Humankind where the majority of fossils are often encased within matrix.
Acknowledgements
We are very grateful to Teresa Kearney and the Ditsong National Museum for providing access to an invaluable reference collection and for valuable discussions over the identification of the specimen. We thank DebTech of De Beers Group Services for permission to use their imaging facilities. We are very grateful to Tea Jashashvili for sharing her comments and advice during the digital extraction and reconstruction of the specimen. We thank Roseberry Languza for his manual preparation work on the specimen. We wish to acknowledge the National Research Foundation and the Department of Science and Technology in South Africa, particularly the African Origins Platform programme, for funding the creation and the development of facilities that contributed to this research.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
Aurore Val performed the 3D rendering and palaeontological identification of the specimen, and wrote the article. Kristian Carlson assisted with the microCT scan of the specimen and assisted with the 3D rendering of the specimen and the writing of the article. Christine Steininger assisted in the palaeontological identification of the specimen, participated in discussions about the palaeoenvironmental implications and made very useful comments on the article. Job Kibii assisted in the palaeontological identification and provided field data as well as useful comments on the article. Cecil Churms allowed access to the microCT scanner and conducted the microCT scan of the specimen. Brian Kuhn photographed the specimen, assisted in the palaeontological identification and commented on the article. Lee Berger is a joint permit holder for the site of Malapa.
References
1. Berger L, De Ruiter DJ, Churchill SE, et al. Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science. 2010;328:195-204. http://dx.doi.org/10.1126/science.1184944 [ Links ]
2. Dirks P, Kibii JM, Kuhn BF, et al. Geological setting and age of Australopithecus sediba from southern Africa. Science. 2010;328:205-208. http://dx.doi.org/10.1126/science.1184950 [ Links ]
3. Brain CK. The hunters or the hunted? An introduction to African cave taphonomy. Chicago: University of Chicago Press; 1981. [ Links ]
4. Brain CK, editor. Swartkrans: A cave's chronicle of early man. Pretoria: Transvaal Museum; 1993. [ Links ]
5. Clarke R, Kuman K. The Sterkfontein caves palaeontological and archaeological site [pamphlet]. Johannesburg: University of the Witwatersrand; 2000. [ Links ]
6. Herries AIR, Curnoe D, Adams JW. A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa. Quat Int. 2009;202:14-28. http://dx.doi.org/10.1016/j.quaint.2008.05.017 [ Links ]
7. Pickering R, Dirks PHGM, Jinnah Z, et al. Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo. Science. 2011;333:1421-1423. http://dx.doi.org/10.1126/science.1203697 [ Links ]
8. Wilkinson MJ. Geomorphic perspectives on the Sterkfontein australopithecine breccias. J Archaeol Sci. 1983;10:512-529. http://dx.doi.org/10.1016/0305-4403(83)90034-1 [ Links ]
9. Hillson S. Teeth. 2nd ed. Cambridge: Cambridge University Press; 2005. [ Links ]
10. Skinner JD, Chimimba CT. The mammals of the southern Africa subregion. 3rd ed. Cambridge: Cambridge University Press; 2005. [ Links ]
11. Avizo®. Version 6.2. Burlington, MA: Visualization Sciences Group SAS; 2010. [ Links ]
12. Avery DM. Notes on the systematics of micromammals from Sterkfontein, Gauteng, South Africa. Palaeontologia Afr. 2000;36:83-90. [ Links ]
13. Avery DM. An assessment of the lower pleistocene micromammalian fauna from Swartkrans members 1-3, Gauteng, South Africa. Geobios. 1998;31(3):393-414. http://dx.doi.org/10.1016/S0016-6995(98)80022-3 [ Links ]
14. Avery DM. The Plio-Pleistocene vegetation and climate of Sterkfontein and Swartkrans, South Africa, based on micromammals. J Hum Evol. 2001;41:113-132. http://dx.doi.org/10.1006/jhev.2001.0483 [ Links ]
 Correspondence to:
Correspondence to:
Aurore Val
Postal address:
Private Bag 3
Wits 2050, South Africa
Email: auroreval696@gmail.com
Received: 10 Jan. 2011
Accepted: 30 June 2011
Published: 07 Nov. 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}