










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.107 n.11-12 Pretoria Jan. 2011
RESEARCH LETTER
Molecular phylogeny of Duvenhage virus
Charmaine van Eeden; Wanda Markotter; Louis H. Nel
Department of Microbiology and Plant Pathology, Faculty of Natural and Agricultural Sciences, University of Pretoria, South Africa
ABSTRACT
The Duvenhage virus (DUVV) constitutes one of the 11 species in the Lyssavirus genus and causes fatal rabies encephalitis. The virus is associated with insectivorous bat species and three human cases have been reported, all of which were linked to contact with bats. Few of these isolates have been studied and thus little is known about the phylogeny and epidemiology of this lyssavirus. Until 2007, when an isolate was made from the East African country of Kenya, all isolations of this virus had been from southern Africa. This discovery led to many questions regarding the spread and diversity of this lyssavirus. Phylogenetic analysis indicated that the DUVV isolates constitute two different lineages, in which the southern African isolates group together to form one lineage and the more recent isolate from Kenya constitutes a new, second lineage. We found that the new isolate has a genetic variation that has not yet been seen for DUVV. Not only is our lack of knowledge regarding the geographical distribution of this uniquely African virus emphasised, but we have also demonstrated the potential diversity within this genotype.
Introduction
The Duvenhage virus (DUVV) belongs to the Lyssavirus genus, a group of single-stranded, negative-sense RNA viruses adapted to replicate in the mammalian central nervous system. All lyssaviruses cause fatal rabies encephalitis and previous classification recognised seven different genotypes (gt) based on the genetic diversity observed between isolates. This classification recently changed and the International Committee for Taxonomy of Viruses now recognises 11 species of lyssaviruses.1 Lyssavirus species are distinguished not only by their genetic diversity but also by other characteristics, such as serological profile, pathogenicity, host species and geographical distribution. The rabies virus (gt 1) (RABV) is the prototype lyssavirus and the other species are known as the rabies-related lyssaviruses (or non-rabies lyssaviruses), namely the Lagos bat virus (LBV) (gt 2), the Mokola virus (MOKV) (gt 3), the DUVV (gt 4), the European bat lyssavirus type 1 (EBLV1) (gt 5), the European bat lyssavirus type 2 (EBLV2) (gt 6), the Australian bat lyssavirus (ABLV) (gt 7)1 and the four most recently recognised species, Aravan (ARAV), Khujand (KHUV), Irkut (IRKV) and West Caucasian (WCBV) bat viruses.2,3 Of these viruses, only RABV, LBV, MOKV and DUVV have been isolated on the African continent, with LBV, MOKV and DUVV being exclusive to Africa. Antibodies that could neutralise WCBV have been identified in bats collected in Kenya,4 and, in 2009, a new lyssavirus species, Shimoni bat lyssavirus, was also isolated from Kenya.5
Based on phylogeny, pathogenicity and serological cross-reactivity, the lyssaviruses have been split into three phylogroups.2,3,6 Phylogroup I consists of RABV, DUVV, EBLV1, EBLV2, ABLV, Aravan, Khujand and Irkut. Phylogroup II consists of MOKV and LBV and Phylogroup III is WCBV.4 All experimental evidence to date suggests that commercial vaccines protect against only the Phylogroup I lyssaviruses and not members of Phylogroups II and III.3,7,8 Nevertheless, cross protection between some lyssavirus species is possible as a result of a similarity in glycoprotein antigenic sites towards which neutralising antibodies are directed.9
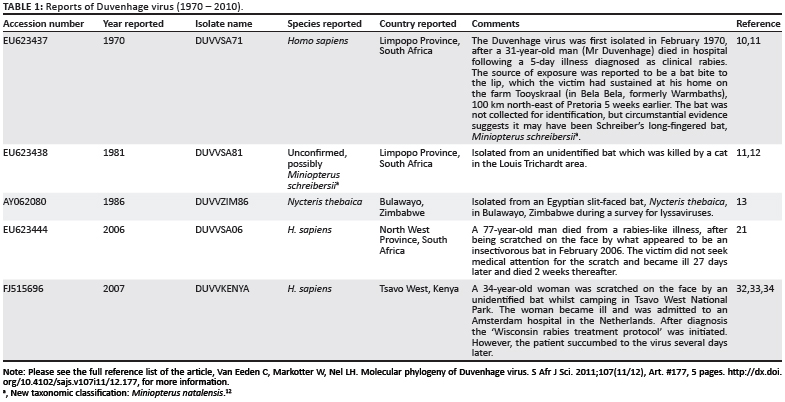
DUVV, for which there have been only five reported cases to date (Table 1), was first encountered in South Africa in 1970 following a human fatality that was associated with a contact exposure involving an unidentified bat.10 DUVV was later isolated from two insectivorous bats. One was suggested to be a member of the Miniopteridae (Miniopterus schreibersii) because of the wide distribution of this genus in the area in South Africa where the exposure occurred.11 New classification now indicates that the bat species previously identified as M. schreibersii in Africa is now considered to be Miniopterus natalensis12; the other isolation was from Nycteris thebaica in Zimbabwe in 1986.13 DUVV was again identified 36 years later in two human fatalities: in South Africa in 2006 and a few months later in Kenya in 2007. In both of these recent cases, there was an exposure to a small bat (probably insectivorous) but the bats were never found or positively identified. These isolations support the notion that the incidence of African rabiesrelated lyssaviruses is underestimated as a result of poor surveillance influenced by the limited diagnostic capacities in African laboratories. Very few laboratories are capable of making a rabies diagnosis with the recommended fluorescent antibody test - which is, in any event, non-differentiating because of the broad spectrum lyssavirus conjugate used. Of these laboratories, only one national African laboratory regularly types positive specimens by monoclonal antibody typing or nucleotide sequencing procedures.
Various studies have shown that DUVV is most closely related to EBLV1, which, although exclusive to Europe, is also associated with insectivorous bat species. A study by Bourhy et al.14 showed that EBLV1 shares more epitopes with DUVV than with EBLV2. Also notable was the fact that EBLV1 and EBLV2 did not form a monophyletic group,15 as EBLV1 and DUVV were more closely related to each other than EBLV1 was to EBLV2. In an investigation to determine the evolution of EBLV,16 it was found that EBLV1 isolates have evolved into at least two genetically distinguishable groups, following geographical drift.15,16 When investigating the host species involved with DUVV and EBLV1 infection, significant overlaps occurred with regard to geographic distribution and co-colonisation in roosts. Myotis myotis and M. schreibersii, for example, have been found to move between colonies and are also known to have direct contact with each other in these mixed colonies.17 The large roost sizes and high population densities of many of these bat species makes them well suited to the sustained transmission and exchange of RNA viruses,18 most likely through the transfer of infectious saliva during licking and biting.19
Several molecular epidemiological studies of RABV have been performed but only a few have been conducted on ABLV,20 EBLV1 and EBLV2,15,16 whilst there have been a limited number of studies focusing on the African lyssavirus.21,22,23 With so little known about the molecular epidemiology of DUVV, and with the addition of two relatively recent viruses to the small pool of known DUVVs, the objective of this study was to investigate the relationship between these DUVV isolates relative to the diversity found in the Lyssavirus genus.
Materials and methods
Viral isolates
All three DUVV isolates from South Africa (DUVVSA71, DUVVSA81 and DUVVSA06) (Table 1) were amplified in the brains of suckling mice. The procedures were approved by the Agricultural Research Council-Onderstepoort Veterinary Institute (ethics approval reference number 15/4P001). Lyophilised passaged brain material was reconstituted in sterile phosphate buffered saline (PBS) (13.7 mmol NaCl, 0.27 mM KCl, 0.43 mM Na2HPO4.2H2O, 0.14 mM KH4PO4, pH 7.3). Two-day-old or three-day-old suckling mice received 30 µ;L of the reconstituted material intracranially.24 Animals were monitored and upon their deaths their brain material was removed aseptically. The direct fluorescent antibody test was used for post-mortem diagnosis of lyssavirus infection according to the standard operational procedure,25 with a polyclonal fluorescein isothiocyanate conjugated immunoglobulin (Onderstepoort Veterinary Institute, Rabies Unit, Pretoria, South Africa) used at a 1:500 dilution. Brain material that tested positive for lyssavirus was pooled and used for RNA extraction.
RNA extraction, reverse transcription polymerase chain reaction and direct sequencing
Total RNA was extracted using a Trizol® reagent (Invitrogen, Cape Town, South Africa) according to the manufacturer's instructions. The reverse transcription polymerase chain reaction (RT-PCR) was performed with the primer set described by Markotter et al.26 Complementary DNA was obtained during reverse transcription with Lys001 (25 ºC for 10 min, 42 ºC for 60 min and 85 ºC for 5 min) in the presence of deoxyribonucleotide triphosphate (10 mM) and avian myeloblastosis virus reverse transcriptase (20 U/µ;L, Roche Diagnostics, Indianapolis, Indiana, USA) and subjected to 30 PCR cycles (94 ºC, 30 s; 37 ºC, 30 s, 72 ºC for 90 s supplemented by a final extension for 7 min at 72 ºC) in the presence of both Lys001 and Lys304 primers using BiolineTaq polymerase (5 U/µ;L, Celtic Molecular Diagnostics, Cape Town, South Africa). After gel electrophoresis, amplicons were purified using the Wizard® SV Gel and PCR Clean-Up System (Promega, Fitchburg, Wisconsin, USA) according to the manufacturer's instructions. Purified amplicons were then subjected to 25 cycles of sequencing (94 ºC for 10 s and 50 ºC for 5 s) and a final cycle at 60 ºC for 4 min using a BigDye® Terminator v3.1 Kit (Applied Biosystems, Foster City, CA, USA) and either a sense or antisense primer. Reactions were stored at -20 ºC before precipitation using the EDTA/ NaOAc/EtOH method according to the BigDye® Terminator v3.1 cycle sequencing protocol (Applied Biosystems). The precipitated reactions were submitted to the sequencing facility of the Faculty of Natural and Agricultural Sciences, University of Pretoria and analysed on an ABI 3100 automated capillary sequencer analyser (Applied Biosystems).
Phylogenetic analysis
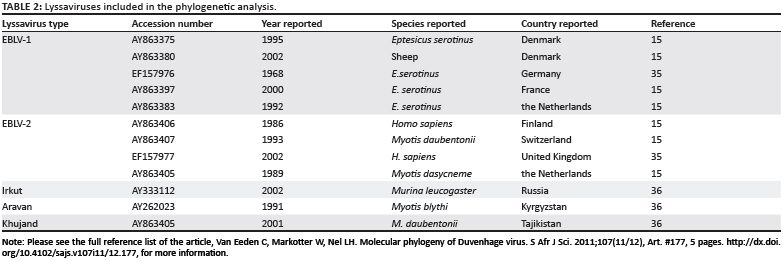
Nucleotide sequencing data were assembled using the VectorNTI 9.1.0 software package (Invitrogen) and trimmed using the BioEdit software package.27 Alignments including the DUVVKenya and DUVVZIM86 (Table 1), as well as other representative lyssavirus sequences (Table 2) available in the public domain, were then carried out using ClustalW.28 Only nt 8-406 of the nucleoprotein gene was used in this analysis for comparison with the DUVVKENYA isolate for which only a partial sequence was available in the public domain. The calculation of genetic distances and the construction of phylogenetic trees based on nucleotide sequence were performed using MEGA 3.1 software.29 Genetic distances were calculated between pairs of sequences using Kimura's 2-parameter method,30 and based on these distances neighbour-joining trees were constructed using the method of Saitou and Nei31. The branching order of the trees was evaluated by using bootstrap analysis of 1000 pseudoreplicate data sets. Results were validated by maximum parsimony as implemented in MEGA 3.1.29
Results
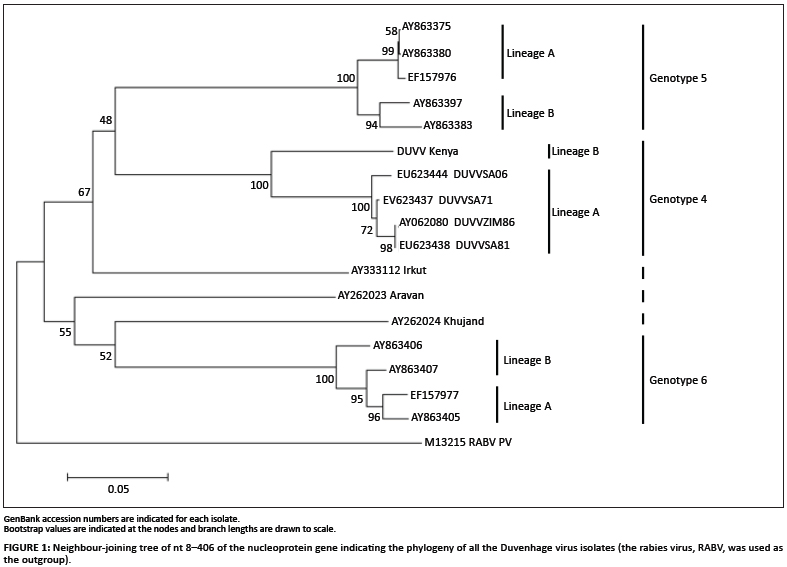
Phylogenetic trees including all five DUVV isolates as well as representative isolates of other lyssavirus species (Table 2) were constructed using a 398-bp fragment of the nucleoprotein gene (nt 8-406). The neighbour-joining method indicated low bootstrap support (71% and lower) for all major clusters representing DUVV, EBLV1 and EBLV2, as well as the newly recognised species ARAV, KHUV and IRKV (Figure 1). DUVV, EBLV1 and EBLV2 all split into two lineages: EBLV1a and EBLV1b; EBLV2a and EBLV2b; and DUVV Lineage A (isolates from southern Africa - South Africa and Zimbabwe) and DUVV Lineage B (the isolate from Kenya). These groupings were supported by high bootstrap values (95% and higher). Maximum parsimony phylogenetic analysis also indicated the major clusters representing the different lyssavirus species as well as the distinct DUVV lineages (results not shown).
Analysis of genetic distances between all five DUVV isolates was carried out using a well-conserved 398-nt sequence from the nucleoprotein gene (nt 8-406). The intrinsic variation between DUVV isolates from southern Africa was low, with a 97.7% - 100% nt identity, even though these isolates were isolated several years apart (ranging from 1971 to 2006). Isolate DUVVZIM86 was found to be 100% identical to DUVVSA81 for this region of the genome, although they were isolated 5 years apart in different countries. The East African isolate, DUVVKenya, had a much lower sequence identity (88.9% - 89.7%) than the other DUVV isolates, which supports the phylogenetic analysis that suggested this isolate forms part of a different lineage. DUVVKenya was shown to be most similar to DUVVSA71 (89.7%) - the first DUVV to have been isolated.
Discussion
Our investigation considered the diversity amongst all known DUVV isolates relative to other members of the Lyssavirus genus. Based on partial nucleoprotein sequences, we confirmed a clear separation of the DUVV isolates from those of EBLV1, although these two groups are most closely related within the lyssaviruses. The DUVV isolates examined also constitute two separate lineages with the longer branch lengths suggesting that these two lineages evolved from an earlier separation than did the lineages of EBLV1. Intrinsic heterogeneity between the DUVV isolates also allows for a clear differentiation between these two lineages. Lineage A isolates, which are from southern Africa, were found to have less than 2% nucleotide variation, even though the isolates were obtained up to 36 years apart. Lineage B, at present consisting solely of the DUVV isolate from Kenya, was found to vary from the lineage A isolates, with respect to this region of the viral RNA, by 11%.
As a result of a number of surveillance programmes for EBLV1, many isolates have been made over the past few decades, demonstrating the significant activity of EBLV1 in bat populations in Eurasia.16 EBLV1a shows phylogenetic homogeneity between isolates across geographical regions, possibly as a result of viral traffic amongst bat populations.18 For EBLV1b, however, geographic origin plays a significant role in phylogenetic clusters, as there is less contact between bat populations18; these observations may also hold true for DUVV. Despite the lack of widespread surveillance, our data suggest that the DUVV cycle is likely to be well established and stable in insectivorous bat species. The relationship between genomic evolution in lyssaviruses and the host's response to infection could explain the rarity of human or other terrestrial mammal encounters with this virus.
As DUVV is present in bat populations, persons interacting with these animals should follow appropriate precautions, including vaccination. Considering the global mobility of humans and animals – and the presence of potential vector species across the globe – African lyssaviruses can easily pose a threat to any continent. It is thus of importance that laboratory, veterinary and medical personnel understand and recognise the resultant public health implications. The demonstrated separation between the Kenyan and the southern African DUVV isolates is indicative of significant spatial evolution that is clearly underestimated by current data, as it may be taken for granted that DUVV cycles are not restricted to South Africa, Zimbabwe and Kenya. It is imperative that more, active surveillance programmes are initiated in order to establish more information about the distribution, prevalence, genetic diversity and host species associated with DUVV.
Acknowledgements
We would like to thank Claude T. Sabeta (Onderstepoort Veterinary Institute, Pretoria, South Africa) and Janusz Paweska (National Institute for Communicable Diseases, National Health Laboratory Services, Johannesburg, South Africa) for providing the South African DUVV isolates.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
W.M. and L.H.N. conceived the experiments and W.M., L.H.N. and C.v.E. designed the experiments. C.v.E. performed the experiments and analysed the data. C.v.E., W.M. and L.H.N. wrote the article.
References
1. International Committee for Taxonomy of Viruses. Official taxonomy: Updates since the 8th report [document on the Internet]. c2009 [cited 2008 Sept. 12]. Available from: http://talk.ictvonline.org/media/p/1208.aspx [ Links ]
2. Kuzmin IV, Orciari LA, Yohko TA, et al. Bat lyssaviruses (Aravan and Khujand) from central Asia: Phylogenetic relationships according to the N, P and G gene sequences. Virus Res. 2003;97:65-79. http://dx.doi. org/10.1016/S0168-1702(03)00217-X [ Links ]
3. Kuzmin IV, Hughes GJ, Botvinkin AD, Orciari LA, Rupprecht CE. Phylogenetic relationships of Irkut and West Caucasian bat viruses within the Lyssavirus genus and suggested quantitative criteria based on the N gene sequence for lyssaviruses genotype definition. Virus Res. 2005;111:28-43. http://dx.doi.org/10.1016/j.virusres.2005.03.008, PMid:15896400 [ Links ]
4. Kuzmin IV, Neizgoda M, Franka R, et al. Possible emergence of West Caucasian bat virus in Africa. Emerg Infect Dis. 2008;14:1887-1889. http://dx.doi.org/10.3201/eid1412.080750, PMid:19046512, PMCid:2634633 [ Links ]
5. Kuzmin IV, Mayer AE, Niezgoda M, et al. Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res. 2010;149:197-210. http://dx.doi.org/10.1016/j.virusres.2010.01.018, PMid:20138934 [ Links ]
6. Badrane H, Bahloul C, Perrin P, Tordo N. Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J Virol. 2001;75:3268-3276. http://dx.doi.org/10.1128/JVI.75.7.3268-3276.2001, PMid:11238853, PMCid:114120 [ Links ]
7. Fekadu M, Shaddock JH, Sanderlin DW, Smith JS. Efficacy of rabies vaccines against Duvenhage virus isolated from European house bats (Eptesicusserotinus), classic rabies and rabies-related viruses. Vaccine. 1988;6:533-539. http://dx.doi.org/10.1016/0264-410X(88)90107-7 [ Links ]
8. Bahloul C, Jacob Y, Tordo N, Perrin P. DNA-based immunization for exploring the enlargement of immunological cross-reactivity against the lyssaviruses. Vaccine. 1998;16:417-425. http://dx.doi.org/10.1016/S0264410X(97)00204-1 [ Links ]
9. Benmansour A, Leblois H, Coulon P. Antigenicity of rabies virus glycoprotein. J Virol. 1991;65:4198-4203. PMid:1712859, PMCid:248855 [ Links ]
10. Meredith CD, Rossouw AP, Van Praag Koch H. An unusual case of human rabies thought to be of chiropteran origin. S Afr Med J. 1971;45:767-769. PMid:5106834 [ Links ]
11. Van der Merwe M. Bats as vectors of rabies. SAfr J Sci. 1982;78:421-422. [ Links ]
12. Miller-Butterworth CM, Eick G, Jacobs DS, Schoeman MC, Harley EH. Genetic and phenotypic differences between South African long-fingered bats, with global Miniopterine phylogeny. J Mammol. 2005;86:1121-1135. http://dx.doi.org/10.1644/05-MAMM-A-021R1.1 [ Links ]
13. Foggin CM. Rabies and rabies-related viruses in Zimbabwe: Historical, virological and ecological aspects. PhD thesis, Harare, University of Zimbabwe, 1988. [ Links ]
14. Bourhy H, Kissi B, Lafon M, Sacramento D, Tordo N. Antigenic and molecular characterization of bat rabies virus in Europe. J Clin Microbiol. 1992;30:2419-2426. PMid:1401009, PMCid:265516 [ Links ]
15. Davis PL, Holmes EC, Larrous F, et al. Phylogeography, population dynamics and molecular evolution of European bat lyssaviruses. J Virol. 2005;79:10487-10497. http://dx.doi.org/10.1128/JVI.79.16.1048710497.2005, PMid:16051841, PMCid:1182613 [ Links ]
16. Amengual B, Whitby JE, King A, Serra Cobo J, Bourhy H. Evolution of European bat lyssaviruses. J Gen Virol. 1997;78:2319-2328. PMid:9292021 [ Links ]
17. Serra-Cobo J, Amengual B, Abellan C, Bourhy H. European bat lyssavirus infection in Spanish bat populations. Emerg Infect Dis. 2002;8:413-420. http://dx.doi.org/10.3201/eid0804.010263, PMid:11971777, PMCid:2730232 [ Links ]
18. Mackensie JS, Field HE, Guyatt KJ. Managing emerging diseases borne by fruit bats (flying foxes) with particular reference to henipaviruses and Australian bat lyssavirus. J Appl Microbiol. 2003;94:59-69. http://dx.doi. org/10.1046/j.1365-2672.94.s1.7.x [ Links ]
19. Ghatak S, Banerjee R, Agarwak RK, Kapoor KN. Zoonoses and bats: A look from human health viewpoint. J Comm Dis. 2000;32:40-48. PMid:11129564 [ Links ]
20. Guyatt KJ, Twin J, Davis P, et al. A molecular epidemiological study of Australian bat lyssavirus. J Gen Virol. 2003;84:485-496. http://dx.doi.org/10.1099/vir.0.18652-0, PMid:12560583 [ Links ]
21. Paweska JT, Blumberg LH, Liebenberg C, et al. Fatal human infection with rabies-related Duvenhage virus, South Africa. Emerg Infect Dis. 2006;12:1965-1966. PMid:17326954 [ Links ]
22. Sabeta CT, Markotter W, Mohale DK, Shumba W, Wandeler AI, Nel LH. Mokola virus in domestic mammals, South Africa. Emerg Infect Dis. 2007;13:1371-1373. PMid:18252112, PMCid:2857310 [ Links ]
23. Markotter W, Kuzmin I, Rupprecht CE, Nel LH. Phylogeny of Lagos bat virus: Challenge for lyssavirus taxonomy. Virus Res. 2008;135:10-21. http://dx.doi.org/10.1016/j.virusres.2008.02.001, PMid:18359532 [ Links ]
24. Koprowski H. The mouse inoculation test. In: Meslin FX, Kaplan MM, Koprowski H, editors. Laboratory techniques in rabies. Geneva: World Health Organization, 1996; p. 80-87. [ Links ]
25. Centers for Disease Control. DFA diagnosis [document on the Internet]. c2008 [cited 2009 July 24]. Available from: http://www.cdc.gov/ncidod/dvrd/Rabies/Professional/Publications/DFA_diagnosis [ Links ]
26. Markotter W, Kuzmin I, Rupprecht CE. Isolation of Lagos bat virus from water mongoose. Emerg Infect Dis. 2006;12:1913-1918. PMid:17326944 [ Links ]
27. Hall TA. BioEdit: A user friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Res. 1999;41:95-98. [ Links ]
28. Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673-4680. http://dx.doi.org/10.1093/nar/22.22.4673, PMid:7984417, PMCid:308517 [ Links ]
29. Kumar S, Tamura K, Nei M. MEGA3: Integrated software for molecular evolutionary genetic analysis and sequence alignment. Brief Bioinform. 2004;5:150-163. http://dx.doi.org/10.1093/bib/5.2.150, PMid:15260895 [ Links ]
30. Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;15:111-120. http://dx.doi.org/10.1007/BF01731581, PMid:7463489 [ Links ]
31. Saitou N, Nei M. The neighbour-joining method: A new method for constructing phylogenetic trees. Mol Biol Evol. 1987;4:406-425. PMid:3447015 [ Links ]
32. Willoughby RE, Tieves KS, Hoffman GM, et al. Survival after treatment of rabies with induction of a coma. New Engl J Med. 2005;352:2508-2514. http://dx.doi.org/10.1056/NEJMoa050382, PMid:15958806 [ Links ]
33. Van Thiel P, Van Den Hoek J, Eftimov F, et al. Fatal case of human rabies (Duvenhage virus) from a bat in Kenya: The Netherlands, December 2007. Eurosurveillance. 2008;13:1. [ Links ]
34. Van Thiel PAM, De Bie RMA, Eftimov F, et al. Fatal human rabies due to Duvenhage virus from a bat in Kenya: Failure of treatment with coma-induction, ketamine and antiviral drugs. PLOS Negl Trop Dis. 2009;3(7):e428. http://dx.doi.org/10.1371/journal.pntd.0000428, PMid:19636367, PMCid:2710506 [ Links ]
35. Marston DA, McElhinney LM, Johnson N, et al. Comparative analysis of the full genome sequence of European bat lyssavirus type 1 and 2 with other lyssaviruses and evidence for a conserved transcription termination and polyadenylation motif in the G-L 3' non translated region. J Gen Virol. 2007;88:1302-1314. http://dx.doi.org/10.1099/vir.0.82692-0, PMid:17374776 [ Links ]
36. Kuzmin IV, Wu X, Tordo N, Rupprecht CE. Complete genomes of Aravan, Khujand, Irkut and West Caucasian bat viruses, with special attention to the polymerase gene and non-coding regions. Virus Res. 2008;136:81-90. http://dx.doi.org/10.1016/j.virusres.2008.04.021, PMid:18514350 [ Links ]
 Correspondence to:
Correspondence to:
Louis Nel
Postal address:
Department of Microbiology and Plant Pathology, University of Pretoria
Pretoria 0002, South Africa
Email: Louis.nel@up.ac.za
Received: 26 Jan. 2010
Accepted: 30 May 2011
Published: 01 Nov. 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}