










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.11-12 Pretoria Jan. 2011
RESEARCH ARTICLE
Single nucleotide and copy number polymorphisms of the SULT1A1 gene in a South African Tswana population group
Hlengiwe P. MbongwaI,II; Petrus J. PretoriusI,II Annemarie KrugerIII; Gerhard KoekemoerIV; Carolus J. ReineckeI,II
ICentre for Human Metabonomics, North-West University, Potchefstroom, South Africa
IISchool of Physical and Chemical Sciences, North-West University, Potchefstroom, South Africa
IIIAfrica Unit for Transdisciplinary Health Research, North-West University, Potchefstroom, South Africa
IVStatistical Consultation Service, North-West University, Potchefstroom, South Africa
ABSTRACT
Previous studies on gene mapping have firmly established variation in segments of the DNA structure amongst different population groups. SULT1A1 is one of four SULT1A genes that maps to the short arm of chromosome 16, and this area has been shown to contain many repetitive sequences and to be highly duplicated. Using the polymerase chain reaction (PCR)-based restriction fragment length polymorphism method, we set out to determine the SULT1A1 genotype and allele frequency distributions in the largest sample studied to date: a homogeneous South African Tswana population of 1867 individuals from the Prospective Urban and Rural Epidemiological (PURE) study, and found the SULT1A1*1 and SULT1A1*2 alleles present at a frequency of 0.68 and 0.32, respectively. This finding corresponded with those obtained for the Black, Caucasian, and mixed-race South African groups reported in previous studies. Next, using a quantitative multiplex PCR method to estimate the SULT1A1 gene number of copies in 459 subjects of our population, we discovered between one and five copies: 0.65% of the subjects had a single copy (allele deletion) and 60.14% of the subjects had three or more copies. Our findings correspond with an earlier study on a small African- American group, but differ from those based on two Caucasian groups. Whereas the genotype distribution was comparable to the Caucasian groups, there was a significant difference in the number of copies, which indicated a genetic link between Tswana and African-American populations despite differences in cultural lifestyle associated with their geographical location.
Introduction
Biochemical sulphation is part of the phase II detoxification metabolism in humans. Human cytosolic sulphotransferase (SULT) enzymes catalyse the sulphate conjugation of many exogenous substrates, such as drugs and some xenobiotics, but also endogenous substrates, such as thyroid hormones1 and neurotransmitters.2 This biotransformation process uses cosubstrate 3'-phosphoadenosine-5'-phosphosulphate to transform the substrates to water-soluble compounds through catalysis by sulphotransferase enzymes,3,4 giving rise to the sulphonated product and the 3'-phosphoadenosine 5'-phosphate.5 This reaction usually is effective in rendering the xenobiotic compound less biologically active and stable enough to be excreted from the body.2 However, the reaction sometimes plays a role in the bioactivation of xenobiotics.6 Population studies on SULT1A1 polymorphisms have been extensive, as it has been shown that compromised sulphonation of xenobiotics by the less active SULT1A1*2 enzyme may predispose an individual to cancer, especially in combination with other risk factors such as advanced age, smoking and a poor diet.2,6,7,8,9,10,11,12,13
The genotype distribution covers only a partial aspect of genetic diversity; the copy number variation should also be considered. The study of human genetic variation at the DNA level constitutes a major challenge,14,15 ranging from large, microscopically visible chromosome anomalies to single nucleotide changes.16 Deletions, insertions, duplications and complex multisite variants,16,17,18 collectively termed copy number variations (CNVs) or copy number polymorphisms (CNPs), are found in all humans.19 Often, CNPs and CNVs are associated with disease, and also with the evolution of the genome itself and have been observed for a number of genes encoding drug metabolising enzymes.20 The extent to which large duplications and deletions contribute to human genetic variation and diversity is largely unknown.19 The dominating type of variation explored so far in the genome has been single nucleotide polymorphisms (SNPs), overshadowing the issue of CNPs (gains and deletions).15 The importance of genotype distribution of the SULT1A1 gene is well documented. However, studies carried out previously on South African subjects contained individuals from different population groups (Black and mixed-race groups)21 and only a limited number of cases were included in a study on Nigerian and African-American groups.7,11 We set out to observe the SULT1A1 genotypes and CNPs in a South African Tswana population group. In this study, we aimed to provide complementary details to the recently published comprehensive study on the genome of a single individual, Archbishop Emeritus Desmond Tutu, from the Sotho-Tswana population group.22
Materials and methods
Subjects
This investigation forms part of an international study, the Prospective Urban and Rural Epidemiological (PURE) programme, which is a prospective cohort study that tracks changing lifestyles, risk factors and chronic disease using periodic standardised data collected in urban and rural areas of 16 countries in transition (developing countries). Full details of the PURE study were published on behalf of all participating scientific groups.23 Ethical approval to participate in the South African component of the PURE (PURE-SA) study was obtained from the North-West University Ethics Committee (reference approval number 04M10). A total of 2010 apparently healthy African volunteers of Tswana ancestry gave informed consent to participate in the baseline study of PURE-SA in 2005 (1002 were from an urban setting around Potchefstroom and 1008 were from three rural villages ~450 km from Vryburg en route to Botswana). The South African population is quite heterogeneous in terms of ethnic and societal characteristics. The experimental group may, however, be regarded as relatively homogeneous, as all participants were of the same ethnic group and complied with the PURE definition of a community as 'a group of people who have common characteristics and reside in a defined geographical area'23.
DNA isolation from blood
Blood was collected intravenously into vacutainer tubes containing ethylenediaminetetraacetic acid (EDTA). Sufficient blood was available for the successful DNA analysis of 1867 of the original 2010 experimental subjects of the PURE study. The FlexiGene DNA kit (Qiagene, Hilden, Germany) was used to isolate DNA from blood. DNA was quantified using a Nano-Drop ND 1000 UV/Vis Spectrophotometer (Nano-DropTechnologies, Wilmington, DE, USA) and was kept at -20 ºC until further use.
SULT1A1 genotyping
Genotypes for the SULT1A1*2 polymorphism (Arg213His, rs9282861) were carried out by using a set of primers, 5'-GTT GGC TCT GCA GGG TTT CTA GGA-3' (forward) and 5'-CCC AAA CCC CCT GCT GGC CAG CAC CC-3' (reverse), which were designed to amplify a region in an intron flanking exon VII as described by Coughtrie et al.7 The primers did not contain a restriction enzyme recognition site. Amplification of DNA was performed in a 50-µL incubation mixture comprising 10 pmol of each primer, 200 µM dNTPs, 2 mM MgCl2, 1 unit GoTaq (Promega, Madison, WI, USA), DNA polymerase, ~100 ng of genomic DNA and 1X Flexi buffer (Promega, Madison, WI, USA). Initial denaturation was at 94 ºC for 5 min. This initial denaturation was followed by 39 cycles of denaturation (at 94 ºC for 30 s), annealing (at 63.4 ºC for 30 s) and extension (at 72 ºC for 30 s). The final extension was at 72 ºC for 10 min. PCR performed under these conditions results in the specific amplification of a 333-bp fragment of DNA which includes exon VII of the SULT1A1 gene. PCR reaction products (5 µL) were digested with 2 units of HaeII (New England BioLabs, Ipswich, MA, USA) restriction enzyme for 90 min at 37 ºC in a 20 µL reaction mixture containing 1X NEBuffer 4 (pH 7.9; New England BioLabs, Ipswich, MA, USA) and 100 µg/mL bovine serum albumin. After digestion, 9 µL of digestion product was resolved on 3% (w/v) agarose gel for 1 h in a 1X TAE buffer (made in-house) (pH 8.5). The 333-bp fragment yielded two fragments of 168 bp and 165 bp, after digestion with HaeII.
The wildtype allele frequency distribution was calculated according to Snyder et al.24 Allele frequency of AA = [(2 x AA) + AH]/2 x N and HH = 1 - AA. Where AA (Arg/Arg) is the total cases of SULT1A1*1 form, AH (Arg/His) is the total cases of the SULT1A1*1/*2 form, HH (His/His) is the total cases of SULT1A1*2 form and N is the total number of cases.
SULT1A1 gene copy number
Because of the high costs of copy number analysis, not all 1867 PURE samples could be analysed for this part of the study. Hence, a proportional stratified sample of 459 was selected to investigate SULT1A1 gene CNP. The stratification was based on gender, area and genotype. The SULT1A1 copy number was estimated by calculating the height and area ratio of the 212 bp amplicon of SULT1A1 to the reference 208 bp amplicon of SULT1A2 as described by Hebbring et al.20 The set of PCR primers used were designed, as per Hebbring et al.20, to co-amplify a 212-bp fragment within exons 2 and 3 of SULT1A1 and a 208-bp fragment within exons 3 and 4 of SULT1A2. In this case, SULT1A2 was used as an internal control for copy number. Amplification of the required DNA fragments from ~25 ng genomic DNA was performed in a 25-µL incubation mixture comprising 0.4 µM each of SULT1/2 forward (5'-6FAM/TCA CCG AGC TCC CAT CTT-3'), SULT1/2 reverse (5'-GTTT GGG GCA GGT GTG TCT TCAG-3'), Factor V forward (5'-6FAM/ATG GAC TTC CAC ATT AGG GAC-3') and Factor V reverse (5'-GTTT GAA GGT AGT GGA TTC TCC ATCA-3') primers; 200 µM dNTPs; 2 mM MgCl2; 1 unit GoTaq DNA polymerase and 1X Flexi buffer (pH 9.0). Initial denaturation was at 95 ºC for 10 min. This was followed by 29 cycles of denaturation (at 95 ºC for 30 s), annealing (at 58 ºC for 30 s), and extension (at 72 ºC for 30 s). The final extension was at 72 ºC for 10 min.
PCR product (1 µL) was diluted a 100X with high-performance liquid chromatography-grade purified water, and 1 µL of this dilution was added to a 9-µL mixture of Hi-Di formamide and GeneScan 500 LIZ size standard (15 µL LIZ size standard for every 1 mL of Hi-Di formamide; both Applied Biosystems, Foster City, CA, USA) in a PCR microtitre well plate. After loading samples, the plate was incubated at 94 ºC for 5 min (denaturation). Thereafter the microtitre well plate was centrifuged at 1500 rpm for 30 s at room temperature, and the samples were analysed further in a Hitachi 3130xl genetic analyser (Applied Biosystems, Foster City, CA, USA) according to the conditions recommended by the manufacturer. The peak heights between 100 and 7500 (arbitrary units) were accepted as the minimum and maximum (samples whose peak heights were higher than 7500 were re-analysed but diluted 150X, and samples whose peak heights were lower than 100 were re-analysed but diluted 75X). The samples were analysed in duplicate.
Statistical analyses
If Χ1, Χ2 and Χ3 indicate the number of times each of these genotypes are observed over the n trials, then Χ = (Χ1, Χ2, Χ3) follows a multinomial distribution with parameters n and π = (π1, π2, π3), where π1, π2 and π3 are the true (population) occurrence probability of the three genotypes and π1 + π2 + π3 = 1. The estimates of these probabilities were calculated and the method proposed by Goodman25 was used to construct the 95% simultaneous confidence intervals for π = (π1, π2, π3). Chi-square dependency tests were also performed to determine if the distribution of the copy number differed between Caucasian and African populations. Histograms and bar charts are used for graphical presentation. SPSS version 18,26 Statistica version 9,27 R28 and Excel29 statistical software programs were used for the statistical analyses.
Results
SULT1A1 genotyping
The SULT1A1 gene is one of the three closely related sulphotransferase genes located on chromosome 16p11.2- 12.1.8,30 Digestion does not take place with DNA fragments amplified from SULT1A1 alleles harbouring the CGC→CAC polymorphism at codon 213, as this alters the restriction site recognition sequence for HaeII (Figure 1). The wildtype, heterozygous and homozygous genotype groups were found to be 43.76%, 47.13% and 9.11% with 95% simultaneous confidence intervals given as [41.03; 46.52], [44.38; 49.90] and [7.63; 10.83], respectively. The wildtype allele frequency distribution was found to be 0.68 for the Tswana group. Copy number estimation The distribution of the calculated SULT1A1/1A2 ratio values, which was used to estimate the number of sult 1A1 gene copies, is shown in Figure 2. The copy number estimation in 459 subjects was as follows: 0.65% demonstrated a deletion within the SULT1A1 gene (which had one copy), and 60.14% had three or more copies, with 95% simultaneous confidence intervals given as [0.17; 2.55] and [48.83; 74.00], respectively (Table 1). It has been shown that CNV is highly replicable and that biallelic CNVs may present in various classes, ranging from simple to complex profiles.



Discussion
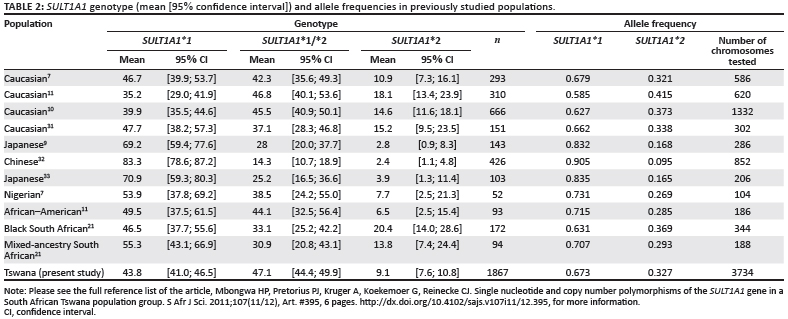
The SULT1A1*2 allele contains a non-synonymous polymorphism and has been extensively reported to occur in individuals in various populations (Table 2).7,9,10,11,21,31,32,33 We used PCR-based restriction fragment length polymorphism, a method known to observe the SULT1A1 genotype distribution in a much bigger (1867), homogeneous South African population group. Availability of nearly 2000 samples from a well-defined profile for each participating individual brought with it uniqueness to this study. From the results presented in Table 2 there seems to be a distinct pattern that populations follow, depending on ancestry. Asian populations show a much higher wildtype genotype frequency when compared to the rest, therefore showing a much lower mutant allele frequency compared to Caucasian populations and African populations. A previous study by Cann et al.34 on mitochondrial DNA concluded that genetic lineage for human origins is in Africa, although the argument is complex, as noted in the study on the genome of the southern African Khoisan and Bantu.22
A study by Dandara et al.21 focused on mixed-race and Black South African populations. Their Black subjects were mainly Xhosa-speaking, and came originally from either the Transkei area of the Eastern Cape or from the Western Cape. The mixed-ancestry individuals are a mix of nationalities resulting from gene flow amongst Black South Africans, Western Europeans, the San, the Khoikhoi, Indonesians and Malaysians who settled in the Cape from the 17th century onwards. The homozygous genotype (Table 2) in mixed-race populations compares relatively closely to the Black South African population but differs to that obtained for the Tswana population. SULT1A1*2 polymorphism has consequences for the sulphonation of drugs and carcinogens.
We calculated 95% simultaneous confidence intervals for this study, as well as for the findings from previous studies (Table 2). This calculation gives lower and upper limits that can be used as a guide to how close the findings are to the true distribution of the population. In general, larger sample sizes produce lower and upper limits that are closer together.
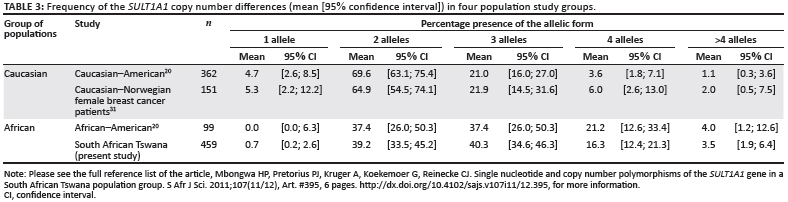
Copy number polymorphisms are found in all humans19; results from previous studies are summarised in Table 3. From the results shown in Table 3, we can conclude that the two Caucasian groups and the two African groups do not differ in distribution. This lack of difference was verified by using a Chi-square dependency test, which produced p-values of 0.651 and 0.72 for the Caucasian and African groups, respectively. We subsequently pooled the Caucasian groups and the African groups to test for a difference in distribution between the Caucasian and African groups, which revealed a p <0.001. This pooled distribution is presented graphically in Figure 3.

These copy number findings confirm results from a much smaller African-American study, and suggests a possible genetic link between the African Tswana and the heritage of the African-Americans. The number of copies demonstrated by each subject does not seem to be conditioned by geography, but could be hereditary.
Copy number variations influence gene expression, phenotypic variation and adaptation by disrupting genes and altering gene dosage, and can cause disease, as in microdeletion or microduplication disorders, or confer risk to complex disease traits. CNVs often represent an appreciable minority of causative alleles at genes at which other types of mutation are strongly associated with specific diseases. Furthermore, CNVs can influence gene expression indirectly through position effects, predispose to deleterious genetic changes, or provide substrates for chromosomal change in evolution.16,35 Because many CNVs include genes that result in differential levels of gene expression, CNVs may account for a significant proportion of normal phenotypic variation.36,37
Information on SNPs and CNPs clearly has a clinical significance. Several studies have established that interindividual and interethnic polymorphic differences are important in responses to factors like chemical exposure and drug treatment, and often relate to the detoxification ability of individuals.38,39 In this regard, it has been shown that the SULT1A1 polymorphism has been associated with an increased risk of breast and oesophageal cancers.21,31,40 Hebbring et al.20 have documented the presence of a common gene duplication or deletion event in SULT1A1. It was shown that individuals carrying additional copies of SULT1A1 represented 'rapid sulphators,' whilst those with fewer copies were considered to be 'slow sulphators' when the SULT1A1 enzyme activity was tested on liver and platelet homogenates. We are currently investigating the activity of SULT1A1 enzyme activity on platelet homogenates. Preliminary results suggest that an increase in copy number benefits individuals of all SULT1A1 genotypes in terms of sulphation abilities. The pharmacogenetic implications of differences in SULT1A1 gene dosage might help explain individual differences in drug toxicity and/or efficacy in the clinical setting (Hebbring et al.20).
Conclusion
Our results on the SULT1A1 allele frequency distribution and copy number estimation per individual were from the largest and most well-defined population group studied thus far and provide data for a comparative view on the SULT1A1 SNPs and CNPs between different population groups (i.e. Tswana, Black and mixed-race). Our findings, together with those by Dandara et al.21 on Black and mixed-race South Africans, bring new information on the SULT1A1 allele frequency distribution regarding South African populations. Identification of susceptible genes facilitates our understanding of the disease process and helps in the discovery of improved medicines and therapeutic regimens to match the target population.20,41 Also, our findings further correspond with those of an earlier study on a small African- American population in the USA, but differ from those on two Caucasian populations. Whereas the allele frequency distributions were comparable in the Caucasian population and our population, there was a significant difference in the number of copies, indicating a genetic link, despite differences in cultural lifestyle associated with geographical location.
Acknowledgements
We are grateful for financial assistance for this study from the National Research Foundation (Scarce Skills PhD Scholarship awarded to HPM), the North-West University Postgraduate Fund and the Biotechnology Partnership and Development (BioPAD) Project BPP007. Professor Scott Hebbring (Mayo Clinic College of Medicine, Rochester, MN, USA) provided the copy number estimation primer sequences, and his advice on the copy number estimation analysis is greatly appreciated. Finally, we thank the people who participated in the PURE study.
Competing interests
We declare that we have no financial or personal relationships which may have inappropriately influenced us in writing this article.
Authors' contributions
H.P. Mbongwa performed the experiments and wrote the manuscript. C.J. Reinecke and P.J. Pretorius were project leaders and advised on the experimental and project design. G. Koekemoer performed the statistical analyses. A. Kruger approved our participation in the PURE study and gave access to the blood samples. As corresponding author, I confirm that each named author contributed to the final manuscript and approved the manuscript submitted for publication.
References
1. Visser TJ, Kaptein E, Glatt H, Bartsch I, Hagen M, Coughtrie MWH. Characterization of thyroid hormone sulfotransferases. Chem-Biol Interact. 1998;109:279-291. http://dx.doi.org/10.1016/S0009-2797(97)00139-7 [ Links ]
2. Hempel N, Gamage N, Martin JL, McManus ME. Human cytosolic sulfotransferase SULT1A1 . Int J Biochem Cell Biol. 2007;39:685-689. http://dx.doi.org/10.1016/j.biocel.2006.10.002 [ Links ]
3. Negishi M, Pedersen LG, Petrotchenko E, et al. Structure and function of sulfotransferases. Arch Biochem Biophys. 2001;390:149-157. http://dx.doi. org/10.1006/abbi.2001.2368, PMid:11396917 [ Links ]
4. Rath VL, Verdugo D, Hemmerich S. Sulfotransferase structural biology and inhibitor discovery. Drug Discov Today. 2004;9:1003-1011. http://dx.doi.org/10.1016/S1359-6446(04)03273-8 [ Links ]
5. Taskinen J, Ethell B, Lautala P, Hood AM, Burchell B, Coughtrie MWH. Conjugation of catechols by recombinant human sulfotransferases, UDPglucuronosyltransferases, and soluble catechol O-methyltransferase: Structure-conjugation relationships and predictive models. Drug Met Dispos. 2003;31:1187-1197. http://dx.doi.org/10.1124/dmd.31.9.1187, PMid:12920175 [ Links ]
6. Coughtrie MWH. Sulfation through the looking glass - recent advances in sulfotransferase research for the curious. Pharmacogenomics. 2002;2:297-308. http://dx.doi.org/10.1038/sj.tpj.6500117, PMid:12439736 [ Links ]
7. Coughtrie WHC, Gilissen RAHJ, Shek B, et al. Phenol sulphotransferase SULT1A1 polymorphism: Molecular diagnosis and allele frequencies in Caucasian and African populations. Biochem J. 1999;337:45-49. http://dx.doi.org/10.1042/0264-6021:3370045, PMid:9854023, PMCid:1219934 [ Links ]
8. Weinshilboum R. Phenol sulfotransferase inheritance. Cell Mol Neurobiol. 1988;8(1):27-34. http://dx.doi.org/10.1007/BF00712908, PMid:3042142 [ Links ]
9. Ozawa S, Shimizu M, Katoh T, et al. Sulfating-activity and stability of cDNA expressed allozymes of human phenol sulfotransferase, ST1A3*1 ((213)Arg) and ST1A3*2 ((213)His), both of which exist in Japanese as well as Caucasians. Biochemistry. 1999;126:271-277. [ Links ]
10. Sun XF, Ahmadi A, Arbman G, Wallin A, Asklid D, Zhang H. Polymorphisms in sulfotransferase 1A1 and glutathione S-transferase P1 genes in relation to colorectal cancer risk and patients' survival. World J Gastroenterol. 2005;11:6875-6879. PMid:16425401 [ Links ]
11. Nowell S, Ratnasinghe DL, Ambrosone CB, et al. Association of SULT1A1 phenotype and genotype with prostate cancer risk in African-Americans and Caucasians. Cancer Epidem Biomar. 2004:13:270-276. http://dx.doi. org/10.1158/1055-9965.EPI-03-0047 [ Links ]
12. Glatt HR. Bioactivation of mutagens via sulfation. FASEB J. 1997;11:314-321. PMid:9141497 [ Links ]
13. Nowell S, Sweeney C, Winters M, et al. Association between sulfotransferase 1A1 genotype and survival of breast cancer patients receiving tamoxifen therapy. Nat Cancer Instit. 2002;94:1635-1640. [ Links ]
14. Buckley PG, Mantripragada KK, Piotrowski A, De Stahl TD, Dumanski JP. Copy-number polymorphisms: Mining the tip of an iceberg. Trends Genet. 2005;21:315-317. http://dx.doi.org/10.1016/j.tig.2005.04.007, PMid:15922827 [ Links ]
15. McCarroll SA, Hadnott TN, Perry GH, et al. Common deletion polymorphism in human genome. Nature Genet. 2006;38:86-92. http://dx.doi.org/10.1038/ng1696 [ Links ]
16. Redon R, Ishikawa S, Fitch KR, et al. Global variation in copy number in the human genome. Nature. 2006;444:444-454. http://dx.doi.org/10.1038/ nature05329, PMid:17122850, PMCid:2669898 [ Links ]
17. Fredman D, White SJ, Potter S, Eichler EE, Den Dunnen JT, Brookes AJ. Complex SNP-related sequence variation in segmental genome duplications. Nature Genet. 2004;36(8):861-866. http://dx.doi. org/10.1038/ng1401, PMid:15247918 [ Links ]
18. Feuk L, Marshall CR, Wintle RF, Scherer SW. Structural variants: changing the landscape of chromosomes and design of disease studies. Hum Mol Gen. 2006;15(1):R57-R66. http://dx.doi.org/10.1093/hmg/ddl057, PMid:16651370 [ Links ]
19. Sebat J, Lakshmi B, Troge J, et al. Large-scale copy number polymorphism in the human genome. Science. 2004;305:525-527. http://dx.doi.org/10.1126/science.1098918, PMid:15273396 [ Links ]
20. Hebbring SJ, Adjei AA, Baer JL, et al. Human SULT1A1 gene: Copy number differences and functional implications. Hum Mol Gen. 2007;16:463-470. http://dx.doi.org/10.1093/hmg/ddl468, PMid:17189289 [ Links ]
21. Dandara C, Li DP, Walther G, Parker MI. Gene-environment interaction: The role of SULT1A1 and CYP3A5 polymorphisms as risk modifiers for squamous cell carcinoma of the oesophagus. Carcinogenesis. 2006;27(4):791-797. http://dx.doi.org/10.1093/carcin/bgi257, PMid:16272171 [ Links ]
22. Schuster SC, Miller W, Ratan A, et al. Complete Khoisan and Bantu genomes from southern Africa. Nature Lett. 2010;463:943-947. http://dx.doi.org/10.1038/nature08795, PMid:20164927 [ Links ]
23. Teo K, Chow CK, Vaz M, Rangarajan S, Yusuf S. The Prospective Urban Rural Epidemiology (PURE) study: Examining the impact of societal influences on chronic noncommunicable diseases in low-, middle-, and high-income countries. Am Heart J. 2009;158:1-8. http://dx.doi.org/10.1016/j.ahj.2009.04.019, PMid:19540385 [ Links ]
24. Population genetics. In: Snyder LA, Freifelder D, Hartl DL, editors. General genetics. Boston, MA: Jones and Bartlett, 1985; 494-528. [ Links ]
25. Goodman LA. On simultaneous confidence intervals for multinomial proportions. Technometrics. 1965;7:247-254. http://dx.doiorg/10.2307/1266673 [ Links ]
26. Statistical Package for the Social Sciences (SPSS). Version 18. Armonk, NY: IBM; 2009. [ Links ]
27. Statistica. Version 9. Tulsa, OK: StatSoft; 2009. [ Links ]
28. R. Version 2.12.0. Vienna: R Development Core Team; 2010. [ Links ]
29. Windows Excel 7. Redmond, WA: Microsoft Corporation; 2009. [ Links ]
30. Hempel N, Masahiko N, McManus ME. Human SULT1A genes: Cloning and activity assays of the SULT1A promoters. Methods Enzymol. 2005;400:147-165. http://dx.doi.org/10.1016/S0076-6879(05)00009-1 [ Links ]
31. Gjerde J, Hauglid M, Breilid H, et al. Effects of CYP2D6 and SULT1A1 genotypes including SULT1A1 gene copy number on tamoxifen metabolism. Ann Oncol. 2007;19(1):56-61. http://dx.doi.org/10.1093/ annonc/mdm434, PMid:17947222 [ Links ]
32. Han DF, Zhou X, Hu MB, et al. Sulfotransferase 1A1 (SULT1A1) polymorphism and breast cancer risk in Chinese women. Toxicol Lett. 2004;150:167-177. http://dx.doi.org/10.1016/j.toxlet.2004.01.012, PMid:15093672 [ Links ]
33. Ohtake E, Kakihara F, Matsumoto N, et al. Frequency distribution of phenol sulfotransferase 1A1 activity in platelet cells from healthy Japanese subjects. Eur J Pharm Sci. 2006;28:272-277. http://dx.doi.org/10.1016/j. ejps.2006.02.008, PMid:16621480 [ Links ]
34. Cann RL, Stoneking M, Wilson AC. Mitochondrial DNA and human evolution. Nature. 1987;325:31-36. http://dx.doi.org/10.1038/325031a0, PMid:3025745 [ Links ]
35. Korbel JO, Kim PM, Chen X, et al. The current excitement about copy-number variation: How it relates to gene duplications and protein families. Curr Opin Struct Biol. 2008;18:1-9. http://dx.doi.org/10.1016/j. sbi.2008.02.005, PMid:18511261, PMCid:2577873 [ Links ]
36. McCarroll SA, Altshuler DM. Copy-number variation and association studies of human disease. Nat Genet Suppl. 2007;39:S37-S42. [ Links ]
37. Freeman JL, Perry GH, Feuk L, et al. Copy number variation: New insights in genome diversity. Genome Res. 2006;16:949-961. [ Links ]
38. Desta Z, Zhao X, Shin JG, Flockhart DA. Clinical significance of the cytochrome P450 2C19 genetic polymorphism. Clin Pharmacokinet. 2002;41:913-958. [ Links ]
39. Devi SS, Vinayagamoorthy N, Agrawal M, et al. Distribution of detoxifying genes polymorphism in Maharastrian population of central India. Chemosphere. 2008;70:1835-1839. [ Links ]
40. Dalhoff K, Buus JK, Enghusen PH. Cancer and molecular biomarkers of phase 2. Methods Enzymol. 2005;400:618-627. http://dx.doi.org/10.1016/ S0076-6879(05)00035-2 [ Links ]
41. Raftogianis RB, Wood TC, Otterness DM, et al. Phenol sulfotransferase pharmacogenetics in humans: Association of common SULT1A1 alleles with TS PST phenotype. Biochem Biophys Res Commun. 1997;239:298-304. http://dx.doi.org/10.1006/bbrc.1997.7466, PMid:9345314 [ Links ]
 Correspondence to:
Correspondence to:
Hlengiwe Mbongwa
Postal address:
Private Bag X6001
Potchefstroom 2520, South Africa
Email: 21046379@nwu.ac.za, hpmbongwa@live.co.za
Received: 07 Aug. 2010
Accepted: 28 June 2011
Published: 04 Nov. 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}