










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.9-10 Pretoria Out. 2011
RESEARCH ARTICLE
Understanding pathogen transmission dynamics in waterbird communities: at what scale should interactions be studied?
Lindy H. MacGregor; Graeme S. Cumming; Philip A.R. Hockey
DST/NRF Centre of Excellence, Percy FitzPatrick Institute of African Ornithology, University of Cape Town, Cape Town, South Africa
ABSTRACT
Pathogen transmission in animal populations is contingent on interactions between and within species. Often standard ornithological data (e.g. total counts at a wetland) are the only data available for assessing the risks of avian pathogen transmission. In this paper we ask whether these data can be used to infer fine-scale transmission patterns. We tested for non-randomness in waterbird assemblages and explored waterbird interactions using social network analysis. Certain network parameter values were then compared to a data set on avian influenza prevalence in southern Africa. Our results showed that species associations were strongly non-random, implying that most standard ornithological data sets would not provide adequate information on which to base models of pathogen spread. In both aquatic and terrestrial networks, all species regularly associated closely with other network members. The spread of pathogens through the community could thus be rapid. Network analysis together with detailed, fine-scale observations offers a promising avenue for further research and management-oriented applications.
Introduction
Pathogens that cross species boundaries, especially between wild and domestic animals and between non-human animals and humans, have particular socio-economic and conservation importance.1 In many instances, transmission of broad-spectrum pathogens within and between different animal communities depends on finer details of the behaviour of the host species, something that has not been widely quantified for most potential wild reservoirs. In this paper we explore whether such quantification is necessary for waterbirds in the context of avian influenza transmission dynamics. Although there are many available data sets2 that describe fluctuations in the total numbers of birds using a particular wetland, and a smaller number of point count data sets that focus at scales of about 150 m, it is unclear whether these data offer a suitable reflection of species associations (i.e. host-host associations) from which to assess the potential for pathogen transmission.
In wild bird communities, avian influenza virus (AIV) is spread principally by the indirect faecal-oral route.3,4,5 Although higher concentrations of highly pathogenic AIV may be shed via the respiratory tract than via the intestinal tract,6,7 aerosol transmission of the virus is still considered to play a lesser role in the spread of the disease than direct contact between birds, or faecal-oral transmission.6 For aquatic birds, water is the most likely medium in which the virus is transferred between birds,8,9,10 and accumulation of birds in large numbers at wetlands, particularly before migration or during moult, presents a substantial opportunity for efficient viral transmission.9,11,12
The null hypothesis in this instance is that species associations are simply a function of the total diversity of birds present at the study site. If this hypothesis were validated, then waterbird counts at a wetland would be a good indication of fine-scale species associations and hence of the probability of pathogen transmission. Alternatively, if species associations are a function of social or ecological choice on a finer spatial scale than a total count or point count, pathogen transmission is likely to be non-random and epidemiological models of direct pathogen transmission would need to incorporate finer-scale behavioural data. We first evaluated the importance of fine-scale interactions and then used social network analysis to explore the differences between species in their potential as disease vectors in the community. Having obtained estimates of vector potential in this way, we then compared our network-based estimates to a data set on avian influenza prevalence in southern Africa.
Methods
Study area
The study was conducted at Strandfontein wastewater treatment works (34º05'S, 18º31'E) in the Western Cape Province, South Africa. The site consists of an agglomeration of 28 ponds (originally constructed over a network of ephemeral wetlands) separated by service roads. Of the 28 ponds, 20 ponds with surface areas ranging between 5000 m2 and 170 000 m2 were chosen for data collection based on ease of observation and the regular occurrence of the chosen study species. Pond types varied in water depth, extent of open water areas, and the presence of reed beds, sand banks and mud flats along the water's edge.
Data were collected between 07:00 and 18:00 over a period of 20 days between 17 October and 09 November 2006. Except for 2 days when ponds were sampled both in the early morning and late afternoon, each pond was sampled only once a day. The time of the day at which a particular pond was sampled varied throughout the study period.
Species assemblages
Field sampling focused mainly on species within the family Anatidae because of their known high sensitivity to AIV infection.11,13 Within this group, we excluded uncommon and rare species, further limiting sampling to Egyptian geese (Alopochen aegyptiaca), Maccoa ducks (Oxyura maccoa), southern pochards (Netta erythrophthalma), Cape teal (Anas capensis), red-billed teal (Anas erythrorhyncha), Cape shovelers (Anas smithii) and yellow-billed ducks (Anas undulata). Because of their strong spatial association with anatids in open water, three additional aquatic bird species were included in the analyses: the black-necked grebes (Podiceps nigricollis), little grebes (Tachybaptus ruficollis) (both Order Ciconiiformes) and red-knobbed coots (Fulica cristata, Order Gruiformes).
Data collection
We quantified the strength of waterbird species associations for both aquatic and terrestrial social networks in the wetland. The strength of association between two individuals was assessed by the frequency of their co-occurrence within 3 m of one another (from now on referred to as close or fine-scale association). Species co-occurrence at fine scales is relevant because more immediate exposure to a higher viral load greatly increases the likelihood that the threshold for infection is exceeded.6
Species associations were recorded by the same observer throughout the study period. A focal individual was randomly selected as a starting point (Figure 1), and the identities of its nearest two neighbours (if present) within a 3-m surrounding radius were determined. Conspecifics were included as potential neighbours, and thus a single species (but not individual) could occupy both neighbouring positions of association. An individual could be recorded as an associate to more than one focal individual on the pond, but was only recorded once as a focal individual itself. One by one each individual on the pond (together with its two nearest neighbours) was recorded, moving systematically across the pond so as not to miss any individuals. Data recording was considered complete for a pond for that day when each individual on the pond had been selected as a focal individual. For each observation, we noted the habitat type (in water but more than 3 m from reeds, in water but within 3 m of reeds, or on land), as well as the water depth category where applicable (< 0.5 m, 0.5 m - 1 m, or > 1 m deep) within which each association took place. These data were used to investigate correlations between species associations and the habitat type or water depth in which associations occurred. Although the individuals' actions during associations were not recorded, the study species mainly utilise the pond water as a feeding ground, whereas associations on land were during either roosting or preening.

Data analyses
Calculation of species association indices
For analysis, associations on land (sand or mud banks) were separated from associations in the water: pooling the data would require accounting for time spent in and out of the water by each species. We analysed the recorded associations within each pond for each observation day independently. For instance, the data collected at pond P1 on 17 October represented an individual pond day.
To compute a value that would serve as an association index between two species in a pond, the following formula was applied to each pond day:

where λ1-2 is the association index between species 1 and 2, n1 and n2 are the respective abundances of species 1 and 2, A is the number of associations recorded between individuals of species 1 and 2, T is the potential number of associations which could have been recorded (equal to twice the total number of focal individuals present in the pond at the time of sampling), and x is the total number of associations (between individuals of all species) recorded for the pond day.
For example, a pond might contain 20 Egyptian geese, 10 Cape teal and 15 little grebes. Therefore, the total number of focal individuals present is 45. Given a potential maximum of 2 associations per focal bird, a maximum of 90 associations (T = 90) could have been recorded. However, it is possible that only 60 associations were observed (x = 60), because some focal individuals may not have had two neighbours within a 3-m surrounding radius. Thus, when calculating the association index (λ) between little grebes and Egyptian geese on this pond day, the expressions 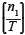 and
and  represent the abundance of little grebes and Egyptian geese, respectively, as a proportion of twice the number of focal birds present. Computing the square root of each of these proportions before multiplication adjusts for the difference in species frequencies by decreasing the size difference between the two proportions, thereby reducing the potential bias that could arise from a large variation in species frequencies on the pond. The value of x is constant for each pond day, and division by x corrects for the difference in actual pairings recorded between ponds. This standardisation is necessary because in an instance where two ponds contain the same number of focal individuals of each species, the measure of association should reflect the fact that the pond with the greater total number of close associations is also likely to include a greater number of random associations.
represent the abundance of little grebes and Egyptian geese, respectively, as a proportion of twice the number of focal birds present. Computing the square root of each of these proportions before multiplication adjusts for the difference in species frequencies by decreasing the size difference between the two proportions, thereby reducing the potential bias that could arise from a large variation in species frequencies on the pond. The value of x is constant for each pond day, and division by x corrects for the difference in actual pairings recorded between ponds. This standardisation is necessary because in an instance where two ponds contain the same number of focal individuals of each species, the measure of association should reflect the fact that the pond with the greater total number of close associations is also likely to include a greater number of random associations.
We calculated the association index for each potential species pair, including intraspecific associations, on each pond day and then grouped the results for reciprocal species pairs. For example, results for λ1-2 were grouped with those for λ2-1. Species associations were therefore viewed as being bidirectional. Each species pair's group of association indices (which comprises indices from each of the pond days) is termed its index group. A Kruskal-Wallis analysis of variance by ranks was used to test the significance of differences between index groups. Statistical analysis was performed using Statistica version 7.14 Because the association indices in the majority of index groups were not normally distributed, we used the median value of each as a representative association strength (both conspecific and heterospecific).
For many of the pond days, the individuals of a species were never recorded close enough to the individuals of another species to form recognised interspecific associations, resulting in many association indices of zero. To prevent a large number of zeros within index groups from skewing the data, we screened all indices equalling zero by removing the association index (= 0) from the group if there were fewer than four individuals of a species present in the particular pond, or if the number of individuals of a species amounted to less than 30% of all individuals present in the pond. This strategy meant that species that seldom came into contact with other species, because of their relative scarcity in a pond, were not discriminated against by according them low median values as their representative association strengths.
Hypothesis testing: Matrix correlations
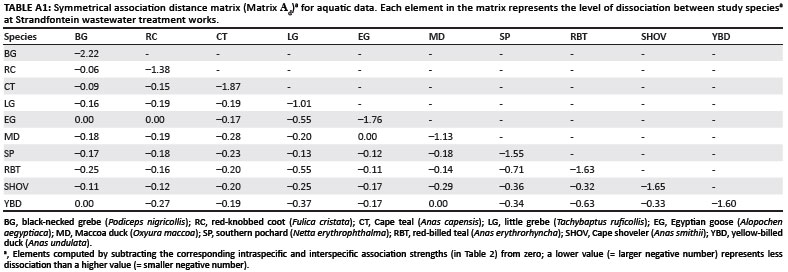
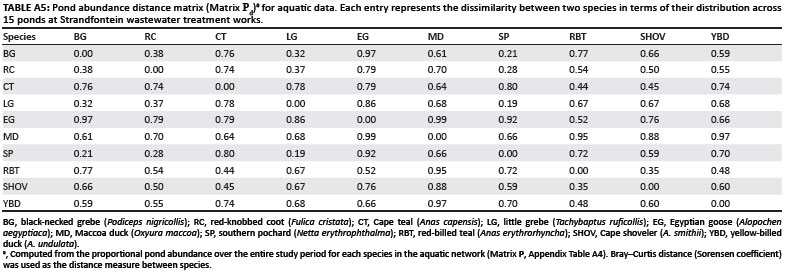
Performing matrix correlations for aquatic data required a distance matrix of species association strengths. The median values of the index groups for the aquatic data were converted to dissociation strengths by subtracting each value from zero. These values represent the level of dissociation between (or within) species, and they were used as elements in a distance matrix (Matrix Ad, Appendix Table A1). A low value such as -2.0 (i.e. a large negative number) therefore indicates less dissociation between (or within) species than a higher value such as -0.5.
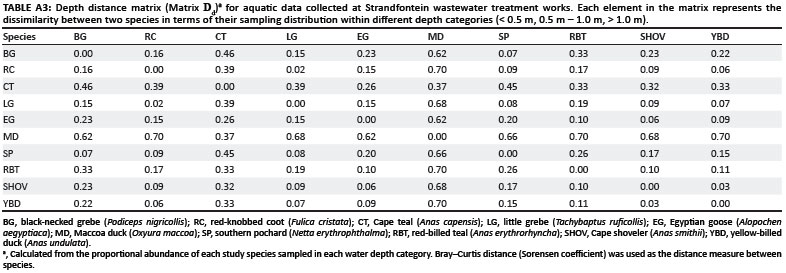
Besides Matrix Ad, two additional distance matrices were developed using the aquatic data; the first described species distributions within basic habitat types (within 3 m of reeds versus in open water) (= Matrix H), and the second described species distributions within water depth categories (= Matrix D). The elements in these matrices represent the proportion of individuals of a species that were recorded in each habitat type and depth category respectively. To produce a habitat distance matrix (Matrix Hd, Appendix Table A2) and a depth distance matrix (Matrix Dd, Appendix Table A3), a Bray-Curtis distance (Sorensen coefficient) was used to measure species dissimilarity.15 Therefore, each element in Matrix Hd and Matrix Dd represents the difference between (or within) species in their overall habitat or water depth preference.
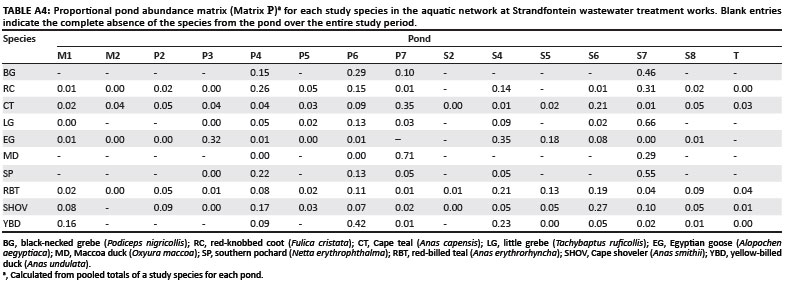
For each species, we calculated the proportional abundance per pond observed across the entire study period, and entered the values into a pond abundance matrix (Matrix P, Appendix Table A4). Data from ponds smaller than 0.6 ha were excluded from this matrix because of low bird abundances, decreasing the pond number to 15. A pond abundance distance matrix was then computed using the Bray-Curtis distance measure (Matrix Pd, Appendix Table A5). We used simple Mantel tests16 to assess the degree of correlation between the dissimilarity of elements in two distance matrices. Thus the Mantel test between Matrix Ad (the primary matrix) and Matrix Dd (the secondary matrix) examined the degree to which species association strengths reflected species' depth preferences. By replacing Matrix Dd with Matrix Hd as the secondary matrix, we tested the correlation between species association strengths and habitat preferences. In the same way, we assessed the degree of concordance between species association strengths and pond abundance, by using Matrix Pd as the secondary matrix. The software package PC-ORD17 was used to compute distance matrices and to perform the Mantel tests, using 1000 Monte Carlo randomisations to evaluate the test statistic.
Network evaluation
We used UCINET 6 for Windows18 to quantify both aquatic and terrestrial association network parameters. The associated NetDraw package19 allowed graphic visualisation of networks. All network diagrams used ordinal data corresponding to the representative median association strength between two species. Because species associations were bidirectional, a single link could be used as a representation of the association strength between two species (referred to as nodes or actors). Network parameters and their definitions (adapted from Hanneman and Riddle20) are summarised in Table 1. When calculating degree centrality, intraspecific associations were ignored and we used adjacency matrices (merely showing the presence or absence of an association) as input parameters, and not the similarity matrices that show actual association strengths between species.
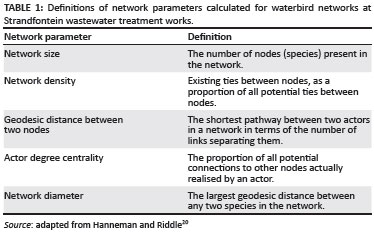
As an overall measure of the connection strength of each species to the rest of the network, we considered the group of median interspecific association strengths belonging to a species and calculated the mean and standard deviation of this group. Median association strengths of zero were omitted from the calculations, and so the resulting mean represented the overall strength of interspecific associations actually realised by the species.
Comparison with independent prevalence data
The final step of the analysis was to test whether empirical data on the prevalence of avian influenza in our study species offered any support to the hypothesis that social interactions may influence influenza prevalence. We used a recently published independent data set on avian influenza prevalence in southern Africa.21 The majority of these data come from three sites: (1) Lakes Manyame and Chivero in Zimbabwe, (2) Barberspan (in the North West Province of South Africa), and (3) Strandfontein (the study site for the analysis presented here). The prevalence of avian influenza across all sampled species in the region is about 2.5%,21 and some of the species considered in this study (particularly the grebes and the diving ducks) are virtually impossible to catch. Recorded prevalence from Strandfontein was not feasible to use on its own because the total numbers of sampled birds and viruses were too low for several of the study species. However, by merging data from all three sites, we were able to obtain workable regional prevalence estimates for six of our study species (numbers following species names indicate the number of sampled birds and the avian influenza prevalence, as a ratio of virus detected to birds sampled): red-knobbed coot (498, 0.014), Cape teal (115, 0.009), Egyptian goose (738, 0.009), red-billed teal (762, 0.046), Cape shoveler (39, 0) and yellow-billed duck (310, 0.006).
We compared these prevalence data to our network-derived measures of mean geodesic distance and actor degree centrality. Because the influence of sample size dominates prevalence estimates, we used partial correlations to compare prevalence to network measures with sample size corrected for. The analysis was run separately for terrestrial and aquatic interaction networks.
Results
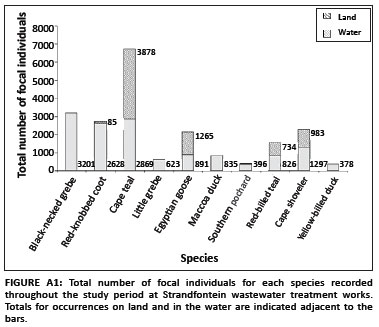
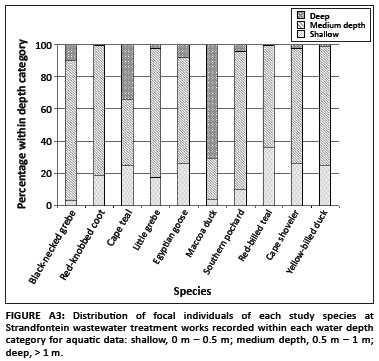
In total, 20 907 focal bird observations were made, resulting in 35 587 associations. Of these associations, 67% occurred in the water and the remaining 33% on land (Appendix Figure A1). The proportion of each focal species observed within different habitat types and water depth categories varied between species (Appendix Figures A2 and A3).
Hypothesis testing: Matrix correlations
For aquatic data, there was no significant correlation between pond abundances of individuals of a species and association strengths in ponds (Mantel test r = 0.041, p = 0.326). Therefore, species association strengths in a pond were not random. Similarly, there were no significant correlations between species association strengths in the water and species' habitat (distance from reeds) or depth preferences (Mantel tests r = 0.076, p = 0.199 and r = 0.110, p = 0.194, respectively). Therefore, neither water depth nor the presence or absence of reeds were significant drivers of species associations.
Network analysis
General network properties
Black-necked grebes, little grebes and Maccoa ducks were never involved in associations on land, resulting in a terrestrial network size of 7. The aquatic network included these three species, resulting in a network size of 10. The aquatic network had an overall density of 0.89, compared with the density of 0.67 for the terrestrial network, indicating that there were relatively more 'missing' associations between species in the terrestrial network than in the aquatic network (Figures 2 and 3).

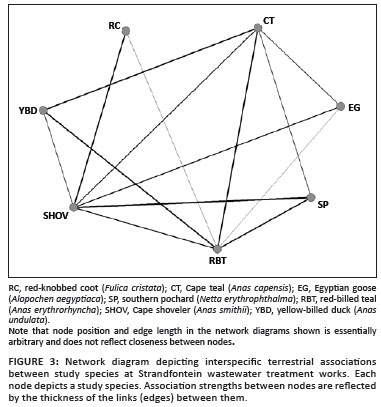
In both networks analysed, all species were either directly or indirectly reachable by others, meaning that no species was completely isolated from other species in the network. The network diameter of both networks was 2.00, further highlighting the cohesive (well-connected) structure of both networks.
Network properties relating to intra-specific and inter-specific associations
Differences between index groups were significant for both aquatic data (Kruskal-Wallis H = 1266.70; χ2 = 1190.16; df = 50; p < 0.001) and terrestrial data (H = 284.29; χ2 = 274.71; df = 20; p < 0.001). The medians representing the interspecific and intraspecific association strengths ranged between 0 and 2.22 in the aquatic network, and between 0 and 2.00 in the terrestrial network (Table 2). The thicknesses of lines linking nodes in the network diagrams (Figures 2 and 3) are proportional to the association strengths between species.
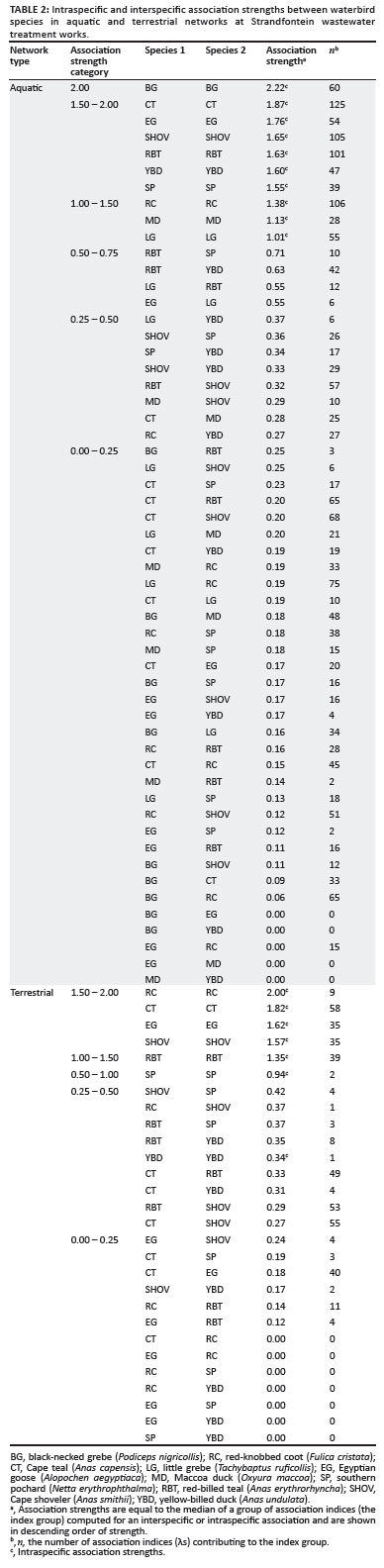
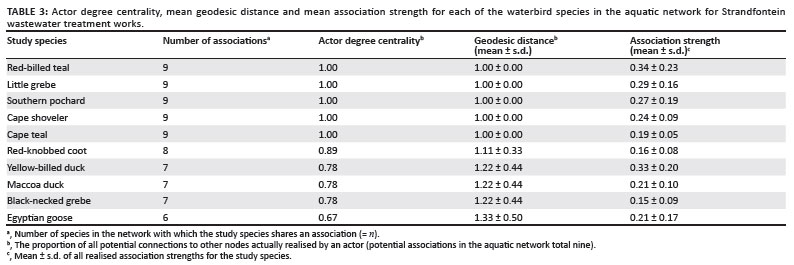
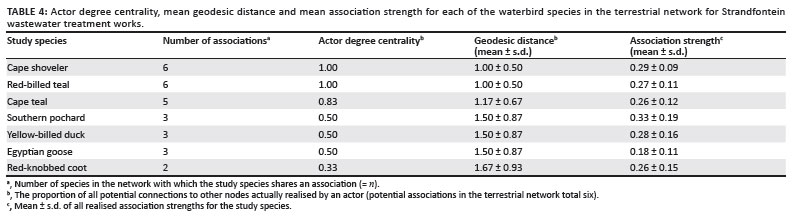
All birds on the water associated more with conspecifics than with heterospecifics (Table 2). Mean association strength, actor degree centrality and mean geodesic distance (Tables 3 and 4) provide an indication of a species' gregariousness and its level of influence in the community in its potential to transmit AIV or contract the virus itself.
Empirical comparison
Correlations of prevalence data with actor degree centrality and mean geodesic distance yielded similar results, although the signs of the correlations were different. For the aquatic network, both metrics were significantly correlated with prevalence (partial correlation coefficient = -0.91 for mean geodesic distance and 0.91 for actor degree centrality; p < 0.02). For the terrestrial network the relationships were not significant at the 0.05 level (partial correlation coefficient magnitude = 0.58; p < 0.3).
Discussion
Waterbird associations both in the water and on land at Strandfontein were clearly not random, supporting the hypothesis that community composition at a point-count or pond scale does not provide an adequate indicator of interaction strengths and hence of transmission capability. Typical waterbird count data alone are thus inadequate to infer the potential for influenza transmission.
Water depth and the presence or absence of reeds could not be used to predict patterns of species associations. It is possible that other habitat variables (such as water pH, salinity and nutrient status) may be influencing patterns of association, but there is little spatial variation in water quality in the ponds at Strandfontein; fine-scale species associations appear to be driven largely by social choice. In other words, a bird may choose to associate with another bird belonging to a different species because it benefits in some way by doing so.22,23,24 These 'choice' associations appear to override fine-scale ecological drivers of association.
This conclusion must be assessed within the context of our network analysis. The small diameters of both networks (2.00) indicate that within each network, all species are easily 'reachable' by others. An indirect route of infection between two species would at most involve one extra species as a link in a chain. Therefore, should viral transmission simply depend on the presence or absence of associations between species and not be a function of association strength, AIV would have the potential to spread rapidly to all species present in the network. The higher network density of the aquatic network indicates that it is more cohesive than the terrestrial network, suggesting that there should be a faster rate of disease spread amongst birds on the water than on land. The greater size of the aquatic network further implies that a greater proportion of species in the wetland community we examined would be more immediately exposed to the virus, were it to be introduced. The time spent by a species in each habitat and the physical parameters affecting viral survival in water or in faeces on land would also play a role in determining the rate of AIV transmission in water and on land, respectively.
The high intraspecific association strengths in both networks (between 1.01 and 2.20 for the aquatic network, and between 0.34 and 2.00 for the terrestrial network) suggest that AIV would spread faster within a species group than between species. Little grebes, red-knobbed coots and Maccoa ducks had the lowest intraspecific association strengths in the aquatic network, although this probably (for the first two species) reflects the fact that they are territorial when breeding and the study occurred during the breeding season.25 Red-knobbed coots showed the highest intraspecific association strength on land, suggesting that intraspecific viral transmission in coots may nonetheless be a potentially important route of infection throughout the year.
Red-billed teal had the highest mean association strength in the aquatic network, portraying this species as potentially the most influential species (for disease transmission) in the network. This result is independently supported by the higher avian influenza prevalence in teal found by Cumming et al.21 If southern pochards are discounted because of their low abundance during the study, then Cape shovelers exhibit the highest mean association strength in the terrestrial network. Based on actor degree centrality, Egyptian geese and red-knobbed coots occupied the least influential positions in the aquatic and terrestrial networks, respectively.
The relatively strong correlation that we found between avian influenza prevalence and measures of network membership is intriguing. We are hesitant to regard it as rigorous proof of the value of a network approach because of the relatively small number of bird species involved in the comparison, the generally low prevalence of avian influenza in the subregion, and the resulting small numbers of positive samples on which the analysis is based. However, the fact that there is a strong and significant correlation between actor location within the aquatic network and documented prevalence lends credence to the approach and assumptions presented here and suggests that this may be a fertile area for further research that links network analysis and epidemiology. It is also interesting that results were significant for the aquatic network but not the terrestrial network, as might be expected for a waterborne pathogen in a waterbird community; we speculate that in these relatively warm habitats, viral survival on the banks may be low and co-feeding rather than co-roosting may thus become the dominant mechanism driving transmission.
The number and strength of associations in a species network (and the ways in which they change through space and time) are not the only factors that could influence the pattern of pathogen transmission. For instance, juveniles may show lower immunological competence,8 making breeding colonies more vulnerable. Differences in AIV susceptibility between species could also affect interspecies transmission, even in a fairly cohesive association network. The nature of interspecific interactions may further influence the potential for interspecies pathogen transmission, and the problem extends beyond the species that we considered in this study. Charadriiformes (wading birds), for example, having also been identified as potential reservoirs for AIV,11,13 and by virtue of feeding in association with anatids,23,26 warrant further investigation for their role in the broader avian influenza disease network.
For diseases which cross the wildlife-domestic interface, further expansion of network analysis should attempt to identify the potential critical links in pathogen transmission between wild and domestic animals. For instance, in South Africa, outbreaks of both high-pathogenic and low-pathogenic AIV in semi-intensively farmed common ostriches (Struthio camelus) have been associated with large numbers of wild waterfowl frequenting ostrich camps on the affected farms.27,28 As a result of the bidirectional nature of interspecific associations, wild birds are placed at risk by ostriches just as much as domestic poultry are placed at risk by wild birds.
Our findings demonstrate that the scale at which interactions are recorded is of prime importance for epidemiological analysis. For health-care professionals who are seeking to evaluate wild bird risks, a combination of information from multiple scales may be required for the accurate depiction of pathogen transmission dynamics. Fine-scale species association data collected at the level of individual ponds revealed strongly non-random species interactions that differed from those interactions that might be inferred from broader-scale abundance data. Although broad-scale patterns are also relevant to understanding which species are present in a given area, transmission potential must be studied at a finer scale if we are to understand the dynamics of AIV in wild populations.
Acknowledgements
We thank Éva Plagányi-Lloyd who developed the association index formula. We also thank Morné du Plessis, Marna Sinclair, Birgit Erni and Morné Carstens and his team at Strandfontein for advice and assistance. This study was funded by the DST/NRF Centre of Excellence at the Percy FitzPatrick Institute of African Ornithology and the USAID-supported Global Avian Influenza Network for Surveillance (GAINS) initiative of the Wildlife Conservation Society's Field Veterinary Program. Further financial support was provided by the Gordon Sprigg Scholarship Fund.
References
1. Cleaveland S, Laurenson MK, Taylor LH. Diseases of humans and their domestic mammals: Pathogen characteristics, host range and the risk of emergence. Philos Trans R Soc London Biol. 2001;356:991-999. doi:10.1098/rstb.2001.0889, PMid:11516377, PMCid:1088494 [ Links ]
2. Taylor PB, Navarro RA, Wren-Sargent M, Harrison JA, Kieswetter SL. TOTAL CWAC report: Coordinated water bird counts in South Africa, 1992-97. Cape Town: Avian Demography Unit; 1999. [ Links ]
3. Slemons RD, Easterday BC. Virus replication in the digestive tract of ducks exposed by aerosol to type-A influenza. Avian Dis. 1978;22:367-377. doi:10.2307/1589291, PMid:358960 [ Links ]
4. Hinshaw VS, Webster RG, Turner, B. Water-borne transmission of influenza A viruses? Intervirology. 1979;11:66-68. doi:10.1159/000149014, PMid:429143 [ Links ]
5. Kida H, Yanagawa R, Matsuoka Y. Duck influenza lacking evidence of disease signs and immune response. Infect Immun. 1980;30:547-553. PMid:7439994, PMCid:551346 [ Links ]
6. Scientific report on migratory birds and their possible role in the spread of highly pathogenic avian influenza. Annex to The EFSA Journal. 2006;357:1-46. [ Links ]
7. Brown JD, Stallknecht DE, Beck JR, Suarez DL, Swayne DE. Susceptibility of North American ducks and gulls to H5N1 highly pathogenic avian influenza viruses. Emerg Infect Diseases. 2006;12:1663-1670. PMid:17283615 [ Links ]
8. Webster RG, Bean WJ, Gorman OT, Chamber TM, Kawaoka Y. Evolution and ecology of influenza A viruses. Microbiol Rev. 1992; 56:152-179. PMid:1579108, PMCid:372859 [ Links ]
9. Tracey JP, Woods R, Roshier D, West P, Saunders GR. The role of wild birds in the transmission of avian influenza for Australia: An ecological perspective. Emu. 2004;104:109-124. doi:10.1071/MU04017 [ Links ]
10. Gilchrist P. Involvement of free-flying wild birds in the spread of the viruses of avian influenza, Newcastle disease and infectious bursal disease from poultry products to commercial poultry. World's Poult Sci J. 2005;61:198-214. doi:10.1079/WPS200451 [ Links ]
11. Alexander DJ. A review of avian influenza in different bird species. Vet Microbiol. 2000;74:3-13. doi:10.1016/S0378-1135(00)00160-7 [ Links ]
12. Delogu M, De Marco MA, Donatelli I, Campitelli L, Catelli E. Ecological aspects of influenza A virus circulation in wild birds of the western Palearctic. Vet Res Comm. 2003;27:101-106. doi:10.1023/B:VERC.0000014125.49371.14, PMid:14535376 [ Links ]
13. Stallknecht DE. Ecology and epidemiology of avian influenza viruses in wild bird populations: Waterfowl, shorebirds, pelicans, cormorants, etc. Avian Dis. 1997;47:61-69. [ Links ]
14. STATISTICA. Version 7. Tulsa: StatSoft Inc.; 2004. [ Links ]
15. McCune B, Grace JB. Analysis of ecological communities. Gleneden Beach: MjM Software Design; 2002. [ Links ]
16. Legendre P. Comparison of permutation methods for the partial correlation and partial Mantel tests. J Statist Comput Simul. 2000;67:37-73. doi:10.1080/00949650008812035 [ Links ]
17. McCune B, Mefford MJ. PC-ORD multivariate analysis of ecological data. Version 4.34. Gleneden Beach: MjM Software Design; 1999. [ Links ]
18. Borgatti SP, Everett MG, Freeman LC. UCINET 6 for Windows: Software for social network analysis. Version 6.138. Harvard: Analytic Technologies; 2002. [ Links ]
19. Borgatti SP. NetDraw: Graph visualisation software. Version 2.055. Harvard: Analytic Technologies; 2002. [ Links ]
20. Hanneman RA, Riddle M. Introduction to social network methods [homepage on the Internet]. c2005 [cited 2007 Jan 10]. Available from: http://www.faculty.ucr.edu/~hanneman/nettext/ [ Links ]
21. Cumming G, Caron A, Abolnik C, et al. The ecology of influenza A viruses in wild birds in southern Africa. EcoHealth. In press 2010. doi: 10.1007/s10393-011-0684-z [ Links ]
22. Dawson R. Marsh sandpipers (Tringa stagnatilis) associating with feeding avocets (Recurvirostra avosetta) and other species. Brit Birds. 1975;68:294-295. [ Links ]
23. Pettet A. Marsh sandpipers (Tringa stagnatilis) associating with feeding teal (Anas crecca). Brit Birds. 1975;68:295. [ Links ]
24. Lewis AD. Two commensal feeding associations observed in Kenya. Scopus. 1989;12:102-103. [ Links ]
25. Hockey PAR, Dean WRJ, Ryan PG, editors. Roberts - Birds of southern Africa. 7th ed. Cape Town: The Trustees of the John Voelcker Bird Book Fund; 2005. [ Links ]
26. Reynolds JF. Feeding association between marsh sandpiper and Hottentot teal. East Afr Nat Hist Soc Bull. 1972:189. [ Links ]
27. Pfitzer S, Verwoerd DJ, Gerdes GH, et al. Newcastle disease and avian influenza A virus in wild waterfowl in South Africa. Avian Dis. 2000;44:655-660. doi:10.2307/1593107, PMid:11007015 [ Links ]
28. Sinclair M, Brückner GK, Kotze JJ. Avian influenza in ostriches: Epidemiological investigation in the Western Cape Province of South Africa. Veterinaria Italiana. 2006;42:5-12. [ Links ]
 Correspondence to:
Correspondence to:
Lindy MacGregor
Postal address: Percy FitzPatrick Institute of African Ornithology, University of Cape Town
Private Bag X3
Rondebosch 7701, South Africa
Email: lindy.macgregor@gmail.com
Received: 19 July 2010
Accepted: 08 May 2011
Published: 16 Sept. 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.
Appendix 1
Sampling distribution of species within habitat types (figures) and matrices used in data analyses (tables)




{kind=link}
{kind=link}