










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.107 n.7-8 Pretoria Aug. 2011
http://dx.doi.org/10.4102/sajs.v107i7/8.613
RESEARCH LETTER
Examination of the Florisbad microvertebrates
Patrick J. LewisI; James S. BrinkII,III; Alicia M. KennedyIV; Timothy L. CampbellV
IDepartment of Biological Sciences, Sam Houston State University, Huntsville, Texas, USA
IIFlorisbad Quaternary Research Department, National Museum, Bloemfontein, South Africa
IIICentre for Environmental Management, University of the Free State, Bloemfontein, South Africa
IVDepartment of Biology, Villanova University, Villanova, Pennsylvania, USA
VDepartment of Anthropology, Texas A&M University, College Station, Texas, USA
ABSTRACT
Florisbad is a Middle Stone Age locality in the Free State Province, South Africa, well known for an archaic Homo sapiens cranium discovered there in 1932. Whilst substantial work has been accomplished on the materials excavated from this site, there is still more to be learned about the palaeoenvironment from the microvertebrates. In broader terms, the make-up and distribution of the Plio-Pleistocene small animal fauna of the Free State Province is underrepresented relative to other provinces, which negatively impacts our understanding of geographic and temporal ranges of many Plio-Pleistocene taxa. Much of the Florisbad small vertebrate material is fragmentary, with diagnostic elements primarily limited to isolated molars. Analysis of this material found a small but diverse assemblage including springhares, rabbits, rodents and reptiles. The small mammal fauna is dominated by springhares, lagomorphs and otomyine and gerbilline rodents. In agreement with previous research on sediments and large mammal fauna, the small animal fauna described here is consistent with an open, treeless grassland.
Introduction
The Florisbad locality, in the Free State Province of South Africa (Figure 1), ranks amongst the country's more important Middle Stone Age fossil localities. This importance is primarily as a result of the recovery of a largely intact archaic Homo sapiens partial cranium in 1932.1 In addition to the hominin cranium, the site has produced a large and diverse fauna over several decades of research.2,3 The fauna associated with this cranium, recovered from what is primarily a spring deposit approaching 12 m in depth, has informed us on the palaeoenvironment of the interior of South Africa in the Middle Pleistocene. However, this prior research has focused primarily on the large mammal component of the fauna whilst the small animal component of the site's fauna has yet to be fully analysed. The specimens described here were excavated from the Florisbad spring during the late 1920s, in 1932 by T.F Dreyer and in 1952 by A.C. Hoffman and J.D. Meiring.2
Whilst springhares (Pedetes sp.) and hares (Lepus sp.) have been identified,2,4 little has been published regarding the remaining small animals of the Florisbad fauna. Kuman and Clark4 list the rodents Tatera sp. (now classified as Gerbilliscus), Otomys spp. (two or three species reported), Parotomys sp. and the possible presence of Cryptomys sp. (identifications attributed to D.M. Avery). However, no further description of the small animal fossils is available. The research presented here is a deeper examination of the small mammals and reptiles recovered from the locality and what they add to our understanding of the palaeoenvironment of this important locality.
Results and discussion
Five specimens (four isolated molars and one mandibular fragment with an associated molar) within the collection are identified to the family Leporidae (rabbits and hares). Unfortunately, identification to lower taxonomic levels is hindered by both an absence of incisors and premolars in the sample and a lack of well-defined dental apomorphies for molars. There are three genera within the Leporidae found in South Africa today - Pronolagus, Lepus and Bunolagus - and six species - P. crassicaudatus, P. randensis, P. rupestris, L. saxatilis, L. capensis and B. monticularis.5,6 Whilst the presence of Leporidae is reported at numerous contemporary fossil localities including Blombos Cave,7 Border Cave,8 Die Kelders Cave I,9,10 Klasies River Mouth Caves,9,11,12 Rose Cottage Cave13 and Sibudu Cave,14 identification is generally at the generic level with Lepus reported most frequently. Of the six extant species found in South Africa, three species (P. rupestris, L. saxatilis and L. capensis) have geographic ranges that overlap with Florisbad.5 Unfortunately, without lower-level taxonomic identifications, the fossil leporids are of minimal use for palaeoenvironmental reconstruction at Florisbad.
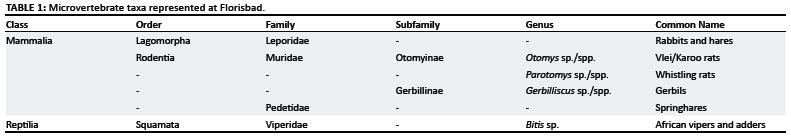
Based on fragmentary jaws, isolated molars and isolated incisors, the rodent fauna is composed primarily of taxa in the Otomyinae (n = 15 molars). Of these 15 specimens, 13 are in the genus Otomys (Figure 2A; Table 1) and 2 are attributed to the genus Parotomys (Figure 2B). It should be noted that whilst some taxonomic treatments classify two of the six species of Otomys reported for the subregion5 as the separate genus Myotomys,6 here we do not make this distinction because of varying degrees of overlap in dental characters between the two genera. Lower first molar specimens with four laminae are present in our sample, and, whilst four laminae in the lower molars are generally associated with modern species of Parotomys,15 such morphology also occurs in some modern species of Otomys.15,16 However, in the genus Parotomys, the third and fourth laminae are joined by an enamel ridge, whilst the third and fourth molars of Otomys remain separate.15 As lower molar morphology overlaps for species within Otomys and also for species within Parotomys, none of the isolated lower molar specimens from either genus can be diagnosed to species. Likewise, the presence of multiple species of Otomys suggested by prior research4 can neither be ruled out nor supported by the lower first molars attributed to the Otomyinae here.
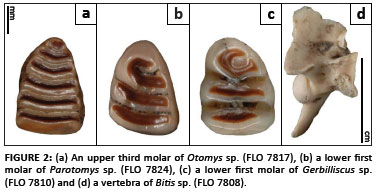
There are also several laminate upper third molars from taxa within the Otomyinae. These specimens all possess six laminae (Figure 2A). The presence of six laminae is consistent with modern species of Otomys. Overlapping morphological characteristics of the third molars for taxa within this genus prevent diagnosing the Florisbad specimens to the species level. Again, the presence of multiple species of Otomys can neither be ruled out nor supported by the upper third molars attributed to the Otomyinae here. Several isolated upper first and second molars, as well as lower second and third molars, are also present. These molars can confidently be attributed to the Otomyinae but are not diagnostic at lower taxonomic levels. Otomyine rodents are common in Middle Stone Age assemblages across southern Africa.17
Three lower first molars are attributed to the family Gerbillinae. The first molars of the Gerbillinae are distinctive in their general pattern because they have an anterior loop of enamel followed by two elongate rings of enamel.5 The molars analysed here are placed in the genus Gerbilliscus (formerly classified as Tatera) (Figure 2C; Table 1). The lower first molars of Gerbilliscus are distinguished from other genera in the subfamily in that Gerbilliscus has no enamel breaking the dentine in the second lamina, and the first lamina has an enamel islet not present in the other genera (although this character may be obscured by wear in older individuals). However, overlapping morphologies in the lower first molars of modern Gerbilliscus do not allow the fossil specimens to be diagnosed to the level of species.15,16 Gerbilline rodents are also common components of many Middle Stone Age assemblages across southern Africa.17
Whilst rodent incisor morphology is not generally sufficient for the diagnosis of taxa, there are at least three incisor morphotypes in the Florisbad fauna. Morphotype 1 consists of upper incisors exhibiting two strong grooves and is consistent with modern members of the genus Otomys. These incisors are also relatively large and fall within the size range of those of modern Otomys. Morphotype 2 consists of lower incisors which also exhibit grooves, although these grooves vary in depth. Whilst Morphotype 2 is consistent in size and shape with those found in modern members of the genus Otomys, the range of variation in incisor morphology is poorly defined for modern taxa in this genus. Morphotype 3 has no grooves. Many modern rodents lack grooves in their incisors, including some members of the genus Otomys. As such, Morphotype 3 specimens cannot be ruled out as belonging to Otomys. In short, whilst three morphotypes are discernable by the difference in their incisors, they may represent a single, variable species within the genus Otomys. Alternatively, they are equally likely to represent multiple taxa from multiple genera, including but not limited to Otomys. Whilst incisors from species within Gerbilliscus generally have incisors narrower than those in Otomys, there is considerable overlap. Species within Gerbilliscus also possess grooved incisors, and several of the Florisbad incisors are both grooved and within the size range of these taxa. Mean values for isolated incisor maximum widths that can be reasonably attributed as either maxillary (n = 29) or mandibular (n = 13) are 2.08 mm and 2.00 mm, respectively. These values are similar to values from a sample of six Otomys specimens housed in the Transvaal Museum, corresponding to each of the six species found within the subregion (maxillary: 1.96 mm; mandibular 1.84 mm). However, these results differ from a sample of two Gerbilliscus specimens (G. leucogaster and G. brantsii) housed in the Transvaal Museum (maxillary: 1.22 mm; mandibular 1.15 mm).
Springhares, in the genus Pedetes, are also common (n = 23) in the Florisbad fauna.2 Whilst fossil species of springhare have been identified from several African sites,17,18,19,20,21 the dental descriptions of these fossil taxa are not sufficient to allow their differentiation from the modern springhare, Pedetes capensis. The taxonomic resolution of the Florisbad springhares is therefore appropriate at the generic level. Modern springhares are ubiquitous in southern Africa and have wide habitat tolerance, although they typically avoid rocky landscapes.22,23
The reptiles in the Florisbad assemblage are represented by two isolated snake trunk vertebrae (Figure 2D; Table 1). Similarities in condition and preservation of the Florisbad specimens suggest that these vertebrae belong to one individual. An hypophysis more than one-third the width of the condyle, the presence of a paracondyler foramen, and enlarged cotyles and condyles place the Florisbad specimens within the family Viperidae (vipers).24,25 Three modern genera - Causus, Bitis and Proatheris - can be found in southern Africa today.26 Both vertebrae have a centrum length of 7.5 mm and a neural spine height of 8 mm, suggesting that they belonged to a large viper. The size of the fossil vertebrae falls within the range of variation for Bitis (African vipers and adders) only. As such, the Florisbad specimens are tentatively assigned to Bitis sp. However, size and biogeography are not derived characteristics for this genus so it is important to maintain a tentative status on this identification. Modern African vipers and adders can be found throughout southern Africa and may be terrestrial, arboreal or aquatic,26,27 and thus provide little for palaeoenvironmental reconstruction.
The large mammal component of the site suggests an open, treeless grassland with a body of water in the immediate vicinity.2,3,4,28,29 Unfortunately, diagnosed to present taxonomic levels the springhares, rabbits and snakes of the Florisbad assemblage offer little information regarding the palaeoenvironment. Whilst the presence of Otomys is often suggested as being indicative of a permanent water source, some populations of modern species within the genus are found in drier habitats.16,22 Likewise, modern species within the genus Gerbilliscus are widely distributed throughout Africa and vary in their habitat preference.16,22,23 However, the small animal fauna from Florisbad offers no reason to suspect that the current palaeoenvironmental picture provided by the large mammal component of the assemblage is incorrect.
Acknowledgements
The authors thank the director of the National Museum in Bloemfontein for the loan of the fossil material. Access to comparative material was granted by the Transvaal Museum in Pretoria.
References
1. Dreyer TF. A human skull from Florisbad, Orange Free State, with a note on the endocranial cast, by C.U. Ariens Kappers. Proc Koninklijke Nederlandse Akademie Wetenschappen. 1935;38:3-12. [ Links ]
2. Brink JS. The archaeozoology of Florisbad, Orange Free State. Mem Nat Mus Bloemfontein. 1987;24:1-151. [ Links ]
3. Brink JS. The evolution of the black wildebeest, Connochaetes gnou, and modern large mammal faunas in Central Southern Africa. DPhil dissertation, Stellenbosch, University of Stellenbosch, 2005. [ Links ]
4. Kuman K, Clarke RJ. Florisbad - New investigations at a Middle Stone Age hominid site in South Africa. Geoarchaeology. 1986;1:103-125. doi:10.1002/gea.3340010201 [ Links ]
5. Skinner JD, Chimimba T. The mammals of the southern African subregion. 3rd ed. Cape Town: Cambridge University Press; 2005. [ Links ]
6. Wilson DE, Reeder DM, editors. Mammal species of the world. A taxonomic and geographic reference. 3rd ed. Baltimore: Johns Hopkins University Press; 2005. [ Links ]
7. Henshilwood CS, Sealy JC, Yates R, et al. Blombos Cave, Southern Cape, South Africa: Preliminary report on the 1992-1999 excavations of the Middle Stone Age levels. J Archaeol Sci. 2001;28:421-448. doi:10.1006/ jasc.2000.0638 [ Links ]
8. Klein RG. The mammalian fauna from the Middle and Later Stone Age (Later Pleistocene) levels of Border Cave, Natal Province, South Africa. S Afr Archaeol Bull. 1977;32(125):14-27. doi:10.2307/3887843 [ Links ]
9. Klein RG. Middle Stone Age man-animal relationships in southern Africa: Evidence from Die Kelders and Klasies River Mouth. Science. 1975;190(4211):265-267. [ Links ]
10. Klein RG, Cruz-Uribe K. Middle and Later Stone Age large mammal and tortoise remains from Die Kelders, Cave 1, Western Cape Province, South Africa. J Hum Evol. 2000;38:169-195. doi:10.1006/jhev.1999.0355, PMid:10627402 [ Links ]
11. Klein RG. Environment and subsistence of prehistoric man in the Southern Cape Province, South Africa. World Archaeol. 1974;5(3):249-284. doi:10.10 80/00438243.1974.9979574, PMid:16468222 [ Links ]
12. Klein RG. The mammalian fauna of the Klasies River Mouth sites, Southern Cape Province, South Africa. S Afr Archaeol Bull. 1976;31(123/124):75-98. doi:10.2307/3887730 [ Links ]
13. Plug I, Engela R. The macrofaunal remains from recent excavations at Rose Cottage Cave, Orange Free State. S Afr Archaeol Bull. 1992;47(155):16-25. doi:10.2307/3888988 [ Links ]
14. Clark JL, Plug I. Animal exploitation strategies during the South African Middle Stone Age: Howiesons Poort and post-Howiesons Poort fauna from Sibudu Cave. J Hum Evol. 2008;54:886-898. doi:10.1016/j. jhevol.2007.12.004, PMid:18234284 [ Links ]
15. Ellerman JR. The families and genera of living rodents. Vol. 4. London: British Museum (Natural History); 1941. [ Links ]
16. De Graaff G. The rodents of southern Africa. Durban: Butterworth; 1981. [ Links ]
17. De Ruiter DJ, Brophy JK, Lewis PJ, Churchill SE, Berger LR. Faunal assemblage composition and paleoenvironments of Plovers Lake, a Middle Stone Age locality in Gauteng Province, South Africa. J Hum Evol. 2008;55:1102-1117. doi:10.1016/j.jhevol.2008.07.011, PMid:18954892 [ Links ]
18. Cooke HBS. Some fossils in the South African Museum collections. S Afr Archaeol Bull. 1952;26:59-69. doi:10.2307/3887445 [ Links ]
19. Cooke HBS. Pleistocene mammal faunas of Africa, with particular reference to southern Africa. In: Bishop WW, Clark JD, editors. Background to evolution in Africa. Chicago: University of Chicago Press, 1964; p. 175-184. [ Links ]
20. Dreyer TF, Lyle A. New fossil mammals and man from South Africa. Bloemfontein: Nasionale Pers; 1931. [ Links ]
21. Zeally AEV. A breccia of mammalian bones at Bulawayo Waterworks Reserve. Proc Rhod Sci Assoc. 1916;15:5-16. [ Links ]
22. Apps P, editor. Smithers' mammals of southern Africa: A field guide. Cape Town: Struik; 2000. [ Links ]
23. Roberts A. The mammals of South Africa. Johannesburg: The Mammals of South Africa Book Fund; 1951. [ Links ]
24. Rage JC. Serpentes, part 11, encyclopedia of paleoherpetology. Stuttgart: Gustav Fischer; 1984. [ Links ]
25. Ikeda T. A comparative morphological study of the vertebrae of snakes occurring in Japan and adjacent regions. Curr Herpetol. 2007;26:13-34. doi:10.3105/1345-5834(2007)26[13:ACMSOT]2.0.CO;2 [ Links ]
26. Branch B. Field guide to snakes and other reptiles of southern Africa. Cape Town: Struik; 1998. [ Links ]
27. Alexander G, Marais J. A guide to the reptiles of southern Africa. Cape Town: Struik; 2007. [ Links ]
28. Brink JS, Lee-Thorp JA. The feeding niche of an extinct springbok, Antidorcas bondi (Antilopini, Bovidae), and its palaeoenvironmental meaning. S Afr J Sci. 1992;88:227-229. [ Links ]
29. Codron D, Brink JS, Rossouw L, et al. Functional differentiation of African grazing ruminants: An example of specialized adaptations to very small changes in diet. Biol J Linn Soc Lond. 2008;94:755-764. doi:10.1111/j.1095-8312.2008.01028.x [ Links ]
 Correspondence to:
Correspondence to:
Patrick Lewis
PO Box 2116
Huntsville, Texas 77341, USA
Email: pjlewis@shsu.edu
Received: 31 Jan. 2011
Accepted: 20 Apr. 2011
Published: 18 July 2011
© 2011. The Authors. Licensee: AOSIS OpenJournals. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}