










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.7-8 Pretoria ago. 2011
http://dx.doi.org/10.4102/sajs.v107i7/8.532
RESEARCH ARTICLE
Antidiabetic effects of Aloe ferox and Aloe greatheadii var. davyana leaf gel extracts in a low-dose streptozotocin diabetes rat model
Du Toit LootsI; Marlien PietersII; Md Shahidul IslamII; Lisa BotesII
ICentre for Human Metabonomics, School for Physical and Chemical Sciences, North-West University, Potchefstroom, South Africa
IICentre of Excellence for Nutrition, North-West University, Potchefstroom, South Africa
ABSTRACT
The medicinal use and commercialisation of the plants Aloe ferox and Aloe greatheadii are primarily based on research done on Aloe vera and Aloe arborescens. Consequently, in this study we investigated the possible antidiabetic effects of ethanol extracts of A. ferox and A. greatheadii var. davyana leaf gel in a streptozotocin (STZ)-induced type 2 diabetes rat model. Fifty male Wistar rats, weighing 200 g - 250 g, were randomly divided into five groups of n = 10: normal control rats, diabetic control rats, diabetic rats receiving A. ferox leaf gel extract (300 mg/kg), diabetic rats receiving A. greatheadii leaf gel extract (300 mg/kg), and diabetic rats receiving glibenclamide (600 µg/kg). Diabetes was induced by a single intraperitoneal injection of STZ (40 mg/kg). Rats were sacrificed 5 weeks after injection, following a 12-hour fast, and blood and tissue samples were collected. Compared to the normal control group, STZ significantly increased relative liver and kidney weights, end-point plasma glucose, fructosamine, oxidative stress, liver enzymes, total cholesterol (TC), triglycerides, very low density lipoprotein-cholesterol and TC: high density lipoprotein-cholesterol (HDL-C) values and reduced serum insulin levels. Treatment with A. greatheadii moderately increased serum insulin and HDL-C levels and moderately reduced end-point plasma glucose and liver alkaline phosphatase (ALP) and significantly decreased TC:HDL-C ratios. A. ferox supplementation similarly resulted in moderately increased serum insulin, accompanied by slight corrections in ALP and HDL-C, without any change to end-point plasma glucose values. A. greatheadii and, to a lesser extent, A. ferox, resulted in a clinically relevant improved diabetic state (indicated by moderate to high effect sizes), suggesting that these Aloe species may show promise for treating diabetes.
Introduction
The increased number of individuals with type 2 diabetes, especially in developing countries, has resulted in an upsurge in interest in the use of natural and traditional remedies for treating this disease.1 The world prevalence of diabetes amongst adults (aged 20 - 79 years) is approximately 6.4% of the total population, equating to 285 million adults, in 2010. As the population ages and becomes more obese, this prevalence is predicted to increase to 7.7% (439 million adults) by 2030.2 The significant worldwide increase in the prevalence of childhood obesity further amplifies the diabetes epidemic.3
Diabetes mellitus is a noncommunicable disease and is considered to be one of the five leading causes of death worldwide. Diabetes mellitus is characterised by hyperglycaemia and hyperlipidaemia, as a result of altered glucose and lipid metabolism.4 Recently, the search for suitable antidiabetic agents has focused on plants used in traditional medicine. Although diabetes is being managed and treated in many developed countries exclusively by conventional medication, in many developing countries, diabetic patients have resorted to traditional medicinal herbs for the treatment of this disease, largely because these are more accessible and less expensive for those living in poor socio-economic conditions.5
Of the many traditional treatments for diabetes, Aloe is the most well known by traditional healers. Beppu et al.6, Rajasekaran et al.7 and others,4,5 have confirmed the antidiabetic effects of certain Aloe species (A. arborescens and A. vera) in streptozotocin (STZ)-induced diabetic rats. However, these beneficial effects were not found by all researchers.8 Aloe ferox and Aloe greatheadii var. davyana, are indigenous to South Africa and, in addition to being widely used by traditional healers, are sold for inclusion in a variety of commercial tonics internationally available for the treatment of a number of ailments, including diabetes. However, the medicinal use and commercialisation of A. ferox and A. greatheadii is primarily based on anecdotal evidence and/or research done on A. vera and A. arborescens. The phytochemical composition and antioxidant capacity of various extracts of A. ferox and A. greatheadii have only recently been described.9,10 This characterisation indicated that the ethanol leaf gel extract, in particular, showed a large variety of phenolic acids or polyphenols, phytosterols, fatty acids, indoles, alkanes, pyrimidines, alkaloids, organic acids, aldehydes, dicarboxylic acids, ketones and alcohols. It was concluded in these studies that A. ferox and A. greatheadii may have potential health benefits, especially for treating diabetes, as a result of their high total polyphenol and total flavonoid contents, their high antioxidant capacities and the presence of a number of individual polyphenols and indoles.9,10
These previous reports subsequently motivated the current exploration to determine the in vivo antidiabetic effects of extracts of A. ferox and A. greatheadii. The extracts were prepared by ethanol extraction,9,10 administered at a similar high dosage level as has been previously described by other groups using other Aloe species,7,11 and tested in a STZ-induced diabetic rat model.
Methods
Collection of plant materials
Whole, freshly cut, A. greatheadii var. davyana leaves (100 kg) were harvested from approximately 200 plants in the month of May (2007) from a rural area in the Potchefstroom district of the North-West Province in South Africa (herbarium deposit site: AP Goossens Herbarium, code PUC, Potchefstroom, South Africa; voucher number: PUC 7951). All leaves were collected from mature plants with a circular diameter greater than 50 cm. Whole, freshly cut, A. ferox leaves (100 kg) were kindly supplied by the Aloe Ferox Trust (herbarium deposit site: AP Goossens Herbarium, code PUC, Potchefstroom, South Africa; voucher number: PUC 9940). These leaves were harvested in September 2007 from farms in the Albertinia region in the Western Cape of South Africa. Leaves were frozen at -20 ºC, transported on dry ice and processed immediately on arrival.
Preparation of Aloe extracts
The A. ferox and A. greatheadii var. davyana ethanol leaf gel extracts were prepared using the method previously described11 with slight modifications as described by Botes et al.9 and Loots et al.10 The ethanol was evaporated to dryness under reduced pressure in a rotary evaporator and the residue stored as aliquots in dry sterilised containers at -20 ºC until further use. These extracts were used within 1 month of their preparation.
Animals, induction of diabetes and interventions
This study was approved by the Ethics Committee (Evaluation Sub-committee for Experimental Animals) of the North-West University (Potchefstroom, South Africa) (Ref: 06D06) and the study was conducted in accordance with the principles of laboratory animal care (NIH Publication; Guide for the care and use of laboratory animals, No. 85-23, revised 1985). Fifty male Wistar rats weighing 200 g - 250 g at the start of the study, were randomly divided into five groups of ten rats each: normal control rats, STZ-induced diabetic control rats, STZ-induced diabetic rats receiving 300 mg/kg A. greatheadii var. davyana leaf gel extract, STZ-induced diabetic rats receiving 300 mg/kg A. ferox leaf gel extract and STZ-induced diabetic rats receiving 600 µg/kg glibenclamide as a positive control.7,11 Rats were housed in standard laboratory rodent cages in a thermally controlled environment with 12-hourly day : night cycles, had free access to water and were fed a standard rodent diet.
Following a 12-h fast, the rats in the diabetic groups were intraperitoneally injected with a single dose of 40 mg/kg STZ (Sigma, St. Louis, MO., USA) prepared in 0.1 M of sodium citrate buffer (pH 4.5). The rats in the normal control group received intraperitoneal injections of the same volume of citrate buffer.7 After 1 week, diabetes was confirmed in the diabetic groups by a blood glucose value > 14 mmol/L.11 Blood was taken by tail prick and blood glucose measured using an Accu-Check® glucometer (Roche diagnostics GMBH, Mannheim, Germany) and Onetouch Surestep glucose strips (Lot No. 285366A, LifeScan Inc., Milpitas, CA., USA). STZ induces diabetes12 by destroying the β-cells of the pancreas by oxidative stress, and thus can be used to develop a model for either type 1 or type 2 diabetes depending on the dosage used. At the dosage of STZ used in this study, a partial destruction of β-cells resulted in a mild insulin-deficient, but non-insulin-dependent state, more closely resembling that of type 2 diabetes than type 1 diabetes.13
Aqueous suspensions of the Aloe leaf gel extracts and glibenclamide, a conventional antidiabetic medication, were freshly prepared daily. Predetermined amounts of the extracts and glibenclamide were dissolved in double-distilled sterile deionised water, to concentrations such that each preselected rat received either 300 mg/kg of one of the two leaf gel extracts or 600 µg/kg glibenclamide (depending on their grouping) in a volume of approximately 1 mL7,11 via intragastric tube once daily for 5 weeks. The rats in the normal control and diabetic control groups received double-distilled water as a placebo via the same route. Rats were weighed weekly, immediately prior to receiving their respective interventions.
Blood and tissue sampling
At the end of the 5-week intervention, rats were fasted for 12 h and then sacrificed by cervical decapitation and blood was collected as described previously.7,11 Blood samples were prepared according to the requirements of the various analytical methods. The blood was centrifuged at 4 ºC for 15 min at 2000 g and plasma and serum were collected. Plasma was prepared in heparin tubes and serum in tubes containing no anticoagulant. Samples were subsequently frozen at -84 ºC until further analysis. The liver, kidney and pancreas of each rat were dissected, rinsed with saline and weighed.
Biochemical analyses
Fructosamine was determined by a colourimetric end-point reaction (Cobas®, Roche diagnostics).
Serum insulin was measured using an Ultrasensitive Rat Insulin ELISA kit (Mercodia AB, Sylveniusgatan, Sweden). The assay was performed as instructed by the supplier. Briefly, during incubation, insulin in the sample reacts with peroxidase-conjugated anti-insulin antibodies and anti-insulin antibodies bound to the microtitration well. The conjugate then reacts with 3,3',5,5'-tetramethylbenzidine in a colourimetric end-point reaction. Results were obtained by measuring an increased absorbance at 450 nm. This analysis was performed on a Multiscan Ascent spectrophotometer (Wiesbaden, Germany).
Serum triacylglycerol (TG), total cholesterol (TC), high-density lipoprotein-cholesterol (HDL-C), low-density lipoprotein-cholesterol (LDL-C) and very low density lipoprotein-cholesterol (VLDL-C) were measured by Pathcare Laboratories (Potchefstroom, South Africa). TC was determined using a polychromatic end-point technique employing horseradish peroxide, HDL-C using the accelerator selective detergent method and LDL-C and triglycerides using a biochromatic end-point technique. Alanine transaminase (ALT), alkaline phosphatase (ALP) and venous blood glucose levels were measured using a Vitros DT60 II Chemistry Analyser (Ortho-Clinical Diagnostics, Rochester, NY, USA), with Vitros reagents and controls.
Ferric-reducing antioxidant power (FRAP) in the serum samples was determined colourimetrically by the FRAP assay as previously described14 using a BioTek FL600 plate reader (Winooski, VT, USA) at a wavelength of 595 nm. The Diacron reactive metabolites (dROMs) test (Diacron International, Grosseto, Italy) was used to measure the serum reactive oxygen metabolite pool. The assay was performed as instructed by the supplier. This colourimetric assay was performed kinetically on a BioTek plate reader measuring change in 560 nm over a period of 15 min at 25 ºC. Quantification was done using a standard and expressed as Carratelli units (CARR U) where 1 CARR U corresponds to 0.08 mg/100 mL hydrogen peroxide.
Statistical analyses
Data is expressed as the mean and 95% confidence interval. Differences between the groups were determined using an analysis of variance (ANOVA). Where significance between the groups was indicated, the Tukey honest significant difference test for unequal n was used to determine where the differences occurred. Statistical significance tests are dependent on sample size and have a tendency to yield smaller p-values as the size of the data set increases.15 Small sample sizes (as is often the case in animal studies) may therefore lead to a failure to detect a true effect.15 In these cases, one can use effect size calculations, which are independent of sample size, as an objective measure of the likelihood of a difference having clinical relevance.16 In order to determine whether the results of this study had clinical relevance, effect sizes were calculated according to the following formula: d = |x1-x2|/smax, where x1 is the mean of one group and x2 the mean of the other group and smax the maximum standard deviation of the two groups. The likelihood of clinical relevance is reported as an effect size (d) and can be interpreted as follows: d < 0.2 is a small likelihood, d = 0.5 is a medium likelihood and d = 0.8 is a large likelihood for effect for parametric data.15 According to Rosenthal et al.16, a statistically nonsignificant effect with a large effect size may indicate a 'failure to detect the true effect', necessitating a larger sample size.
Results
As can be seen in Table 1, five rats did not develop diabetes and/or recovered spontaneously and nine rats died during the study, resulting in unequally sized groups.
Body weight
The normal control rats gained, on average, 72.8 g during the 5-week intervention period, whilst the diabetic control and glibenclamide groups gained, on average, 5.63 g and 21.61 g, respectively. However, the A. greatheadii and A. ferox groups showed mean weight reductions of 9.7 g and 10.9 g, respectively (Table 1).
Relative organ weight
Only small differences were observed in the mean pancreatic weights, expressed as a percentage of body weight, between the five groups (Table 1). Whilst the mean pancreatic weight of the normal control group was not significantly higher than that of the diabetic control group, the effect size calculation indicated that the difference in organ weight may have had a moderate clinical relevance. The glibenclamide intervention group had a significantly lower pancreatic weight compared to the normal control group (0.25% versus 0.29%). The normal control rats also had significantly lower mean liver and kidney weights compared to the diabetic control rats. All three interventions tested did little to change this finding, with all three groups having significantly higher mean liver and kidney weights than those of the normal control group.
Diabetes markers
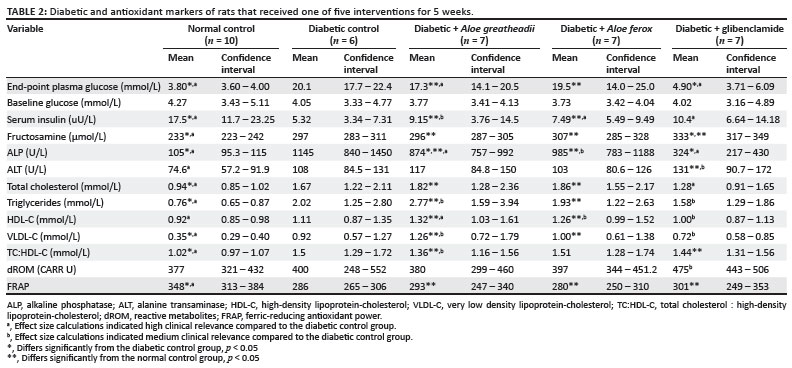
As described in Table 2, baseline glucose determinations done via tail prick revealed no significant differences between groups (p = 0.785), indicating that all the rats were in the same state of normal glycaemia before injection of STZ. End-point plasma glucose values, after the diabetes induction and the interventions that followed, showed significant differences between the groups (p = 0.0001) with the diabetic control group having significantly higher blood glucose values than the normal control group, as would be expected. A. greatheadii did not significantly decrease the diabetes-induced plasma hyperglycaemia, but, as indicated by the effect size calculation, the decrease may be clinically relevant. The A. ferox intervention had a comparatively lower influence. The glibenclamide intervention, on the other hand, had a greater effect, lowering plasma glucose to values that were no longer significantly different from those of the normal control group (p = 0.954). Fructosamine concentrations were significantly elevated in the diabetic control group compared to the normal control group (p = 0.0001) (Table 2). The interventions, however, had no effect on restoring these values. In fact, the fructosamine values in the glibenclamide group were not only increased compared to the normal control group, but also when compared to the diabetic control group. However, this increase was not considered to be of clinical relevance (d < 0.2). As expected, serum insulin levels in the diabetic control group were significantly lower than those of the normal control group (p = 0.001), with effect size calculations also indicating a high clinical relevance (d = 1.49). Serum insulin levels in the A. greatheadii, A. ferox and glibenclamide groups were not significantly different to those of the diabetic control rats, although values, especially in the glibenclamide group, were higher, with effect sizes for all three interventions indicating that these values may have been clinically relevant. Furthermore, these changes in serum insulin levels were inversely correlated to changes in end-point plasma glucose levels, as can be expected.
Liver enzymes
STZ-induced diabetes resulted in a significant increase in plasma ALP (p = 0.0001). The Aloe interventions both resulted in a reduction in diabetes-induced ALP, with a significant reduction noted in the A. greatheadii intervention when compared to the diabetic control group (874.3 U/L versus 1145.0 U/L, p = 0.01). Glibenclamide significantly reduced the diabetes-induced ALP to levels no longer significantly different from normal control levels (323.7 U/L versus 105.3 U/L, p = 0.077). Effect size calculations showed that these changes in all the intervention groups were large enough to have clinical relevance (Table 2). STZ-induced diabetes resulted in a clinically relevant increase in ALT levels, although not statistically significantly so. The Aloe interventions did not correct this increase and the glibenclamide significantly further increased plasma ALT levels (131.2 U/L versus 74.6 U/L).
Lipids
STZ-induced diabetes resulted in significantly increased levels of TC (p = 0.001), triglycerides (p = 0.015), VLDL-C (p = 0.018) and TC:HDL-C (p = 0.0001) in the diabetic control group when compared to the normal control group. Additionally, effect size calculations indicated a high clinical relevance for all these changes (Table 2). The increase in HDL-C in the diabetic control group compared to the normal control group was unexpected. Even so, rats receiving either of the Aloe interventions showed an even greater increase in HDL-C values, resulting in a modest improvement in the TC:HDL-C ratios, when compared to those of the diabetic control group. The glibenclamide intervention was comparatively better at restoring the majority of these diabetic lipid markers to levels no longer significantly different from those of the normal control group (TC: p = 0.060, triglycerides: p = 0.219, HDL-C: p = 0.992 and VLDL-C: p = 0.235), although they were also not significantly different from those of the diabetic control group, but had moderate to large effect sizes.
Oxidative stress and antioxidant markers
STZ-induced diabetes had no effect on dROM values. However, FRAP values were significantly lower in the diabetic control group compared to the normal control group. None of the three treatments significantly corrected this decrease in oxidative stress and antioxidant markers.
Discussion
This is the first intervention study investigating the possible antidiabetic properties of A. greatheadii and A. ferox. As previously described, the phytochemical composition analyses of A. greatheadii and A. ferox leaf gel extracts determined the presence of a number of possible antidiabetic phytochemicals, the majority of which included a variety of phenolic acids, polyphenols and indoles, with proven antioxidant capacity (as assessed by oxygen radical absorbance capacity and FRAP).9,10 Hence, these extracts are thought to be of potential use in the treatment of diabetes, possibly by alleviating hyperglycaemia-induced oxidative stress. These glucose lowering and β-cell protective effects, may additionally lead to improved insulin secretion, thereby further correcting the hyperglycaemia and dyslipidaemia associated with diabetes.5,7,17
Intervention dosages of 300 mg/kg ethanol leaf gel extracts of A. greatheadii and A. ferox (an upper dosage level comparative to other studies and identical to the dosage used for successfully testing ethanol leaf gel extracts of A. vera),7,11 were administered to STZ-induced diabetic rats over a period of 5 weeks. Chronic hyperglycaemia was induced 7 days after an intraperitoneal injection of 40 mg/kg STZ. STZ selectively destroys the pancreatic β-cells, resulting in insulin deficiency, which in turn results in hyperglycaemia.18 Furthermore, because of the uncontrolled diabetic state demonstrated by the chronic and end-point hyperglycaemia in the diabetic control group compared to the normal control group, an expected increase in fructosamine was observed in the diabetic control group as compared to the normal control group (Table 2).8 This chronic hyperglycaemia may have additionally resulted in a hyperglycaemia-induced oxidative stress,19 explaining the slightly increased dROM and significantly reduced FRAP values seen in this STZ-induced diabetes model. Furthermore, STZ-induced diabetes resulted in increased fasting levels of TC, TG, HDL-C, VLDL-C and TC:HDL-C, a finding characteristic of these models,18 as well as increased levels of ALT and ALP in the bloodstream.20 In addition to these biochemical changes, animals of this model of diabetes showed considerable reductions in weight gain, increased liver mass, increased kidney mass and decreased pancreatic mass compared to the control rats, which is consistent with previous findings.21,22,23,24,25
The fact that glibenclamide, a conventional type 2 diabetes medication, almost entirely normalised the induced diabetic state, shows that this STZ-induced diabetes model is satisfactory for investigating non-insulin-dependent diabetes. It has been shown that excessive glucose production, rather than insulin resistance, accounts for hyperglycaemia in recent-onset STZ-induced diabetic rats. Hence, the calculation for determining insulin resistance using standard methods, such as the homeostasis model of assessment, may not be appropriate for this particular model. A review by Islam and Loots18 further supports this assumption: they concluded that a STZ-induced diabetes model is not the most appropriate for monitoring changes in insulin resistance and that other models should be considered for more accurate investigations of this variable (e.g. a high fat diet diabetes model). For this reason, insulin resistance was not evaluated in this study.
The A. greatheadii leaf gel extract intervention resulted in a decreased hyperglycaemic state and increased insulin levels compared to the diabetic control group. Although not statistically significant, the moderate to large effect sizes does indicate that these changes may be clinically relevant. The A. ferox intervention resulted in a similar effect on insulin secretion but did not change the hyperglycaemia. Similarly, the fructosamine concentrations (an indicator of blood glucose control over a 21-day period26) of both Aloe intervention groups remained unchanged. These results suggest that A. greatheadii (and to a lesser extent A. ferox) may show some effect in ameliorating the diabetes-induced hyperglycaemia by increasing insulin secretion from the pancreatic β-cells. However, the unchanged fructosamine levels may be indicative that longer interventions with these extracts may be necessary, or that higher dosages may be required. Furthermore, A. greatheadii had a greater effect, although not significantly so, on correcting the abnormal dROM and FRAP values, when compared to the A. ferox intervention. A possible reason for these nonsignificant changes in oxidative stress markers in the intervention groups is that, for the purpose of measuring the other diabetic markers, the blood was collected after an overnight fast, approximately 24 h after the last Aloe dose was ingested. Hence, the blood sample would not have reflected the direct antioxidant capacity of the blood, from the direct presence of the polyphenols present in the extracts ingested, because these would have already been metabolised by the time the blood was taken. Additionally, effects of the interventions on the increased insulin levels and reduced hyperglycaemia may not have been large enough to have significantly influenced the diabetic rat's general antioxidant status over the 5-week period.
The glibenclamide intervention increased insulin secretion with large clinical relevance, which subsequently totally ameliorated the hyperglycaemic state (p = 0.0001). The fructosamine levels, on the other hand, were unexpectedly significantly increased relative to the diabetic control group. Fructosamine is a marker of long-term glucose control, and the fact that the weekly glucose determinations indicated that the glucose concentrations only returned to normal during the last week of the intervention (data not shown), may explain the lack of improvement in the fructosamine levels. Interestingly, despite glibenclamide restoring the diabetes-induced hyperglycaemia, it had little effect on normalising the oxidative stress markers (dROM and FRAP). Therefore, as is the case with the Aloe interventions, a longer duration of stable blood glucose levels by glibenclamide may be required before these markers return to normal.
Regarding the abnormal lipid profiles induced by the STZ-induced diabetic state, the A. ferox and A. greatheadii interventions resulted in a nonsignificant general increase in the lipid markers analysed, with moderate clinical relevance established for the A. greatheadii intervention (except with respect to TC). This generalised hyperlipidemia seen in these groups may be attributed to an increase in fat absorption via the gut as a result of abnormally increased levels of small intestinal acyl-CoA : cholesterol acyltransferase activity,27 which is known to be elevated when insulin is deficient.28 Unfortunately, the exact effect of insulin on acyl-CoA : cholesterol acyltransferase ratios is still unclear. Another possible explanation could be an increased activation of hormone-sensitive lipase.28 In contrast, the glibenclamide intervention resulted in a general nonsignificant reduction in the elevated hyperlipidemia, with moderate to large effect sizes. Considering the mechanism by which these lipids are elevated during the diabetic state, the reduction in hyperglycaemia as a result of the glibenclamide-induced increased insulin secretion, could have resulted in an inhibition of lipase in the adipose tissue, thereby lowering the amount of free fatty acids released into the bloodstream and the subsequent cascade, leading to the abnormal diabetic lipid profile.29
From these results it is evident that, although most of the changes that occurred in the diabetic state as a result of the two Aloe interventions were not statistically significant, A. ferox and A. greatheadii may (given the effect size calculations) show some potential for restoring hyperglycaemia through increased insulin secretion. There is therefore a possibility that a higher dosage of these or alternative extracts of the same Aloe species, given over a longer period and investigated using larger experimental groups, may result in more significant changes to the variables investigated.
Conclusions
Although not consistently so, many previous studies conducted in the same manner, using identically prepared extracts of other Aloe species, at similar dosages, have reported significant antidiabetic effects over shorter intervention periods. In our pilot study, improvements were observed in end-point glucose, serum insulin, HDL-C and TC:HDL-C, using A. greatheadii leaf gel extracts, in a STZ-induced diabetic rodent model. The A. ferox intervention also showed similar positive effects, however, to a lesser extent. Although the majority of these effects were statistically nonsignificant (likely because of the small sample sizes), they were clinically relevant. We therefore suggest that further in vivo investigations into the use of these Aloe species, using alternative Aloe extracts, longer treatment durations, larger dosages and/or larger sample sizes are needed before conclusive claims can be made regarding the use of these Aloe species for treating diabetes.
Acknowledgement
Funding was provided by the North-West University. Funding did not influence the conduct of the research or the preparation of the article.
References
1. Vasudevan AR, Garber AJ. Insulin resistance syndrome. A review. Minerva Endocrinol. 2005;30:101-119. [ Links ]
2. Shaw JE, Sicree RA, Zimmet PZ. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Atlas. 2010;87:4-14. [ Links ]
3. Murugesh K, Yeligar V, Dash DK, Sengupta P, Maiti BC, Maity TK. Antidiabetic, antioxidant and antihyperlipidemic status of Heliotropium zeylanicum extract on streptozotocin-induced diabetes in rats. Biol Pharm Bull. 2006;29:2202-2205. doi:10.1248/bpb.29.2202 [ Links ]
4. Ducorps M, Ndong W, Jupkwo B, et al. Diabetes in Cameroon. Classification difficulties in Africa. Médecine tropicale: revue du Corps de santé colon. 1996;56:264-270. [ Links ]
5. Agarwal OP. Prevention of atheromatous heart disease. Angiology. 1985;36:485-492. doi:10.1177/000331978503600801, PMid:2864002 [ Links ]
6. Beppu HT, Nagamura Y, Fujita K. Hypoglycemic and antidiabetic effects in mice of Aloe arborescens Miller var. natalensis Berger. Phytother Res. 1993;7:S37-S42. doi:10.1002/ptr.2650070713 [ Links ]
7. Rajasekaran S, Sivagnaman K, Subramanian S. Antioxidant effect of Aloe vera gel extract in streptozotocin-induced diabetes in rats. Pharmacol Rep. 2005;57:90-96. PMid:15849382 [ Links ]
8. Elliot RB, Wasmuth H, Hill J, Songini M, Bottazzo GF, Sardinian IDDM Study Group. Diabetes and cow's milk. Lancet. 1996;348(9042):1657. [ Links ]
9. Botes L, Van Der Westhuizen FH, Loots DT. Phytochemical content and antioxidant capacities of two Aloe greatheadii var. davyana extracts. Molecules. 2008;13:2169-2180. doi:10.3390/molecules13092169, PMid:18830148 [ Links ]
10. Loots DT, Van Der Westhuizen FH, Botes L. Aloe ferox leaf gel phytochemical content, antioxidant capacity, and possible health benefits. J Agric Food Chem. 2007;55:6891-6896. doi:10.1021/jf071110t, PMid:17661490 [ Links ]
11. Rajasekaran S, Sivagnaman K, Subramanian S. Modulatory effects of Aloe vera leaf gel extract on oxidative stress in rats treated with streptozotocin. J Pharm Pharmacol. 2005;57:241-246. doi:10.1211/0022357055416, PMid:15720789 [ Links ]
12. Szkudelski T. The mechanism of alloxan and streptozotocin action in b-cells of the rat pancreas. Physiol Res. 2001;50:536-546. [ Links ]
13. Portha B, Giroix MH, Kergoat M, Bailbe D, Blondel O, Serradas P. Animal models of non-insulin-dependent diabetes induced in the rat by experimental reduction of β cell mass. J Annu Diabetol Hotel Dieu. 1988:33-36.
14. Benzie IF, Strain JJ. Feric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999;299:15-27. doi:10.1016/S0076-6879(99)99005-5 [ Links ]
15. Ellis SM, Steyn HS. Practical significance (effect sizes) versus or in combination with statistical significance (p-values). Manag Dyn. 2003;12:51-53. [ Links ]
16. Rosenthal R, Rosnow RL, Rubin DB. Contrasts and effect sizes in behavioural research: A correlational approach. Cambridge: Cambridge University Press; 2000. [ Links ]
17. Beppu HT, Koike K, Shimpo T, Chihara M, Hoshino CI, Kuzuya H. Radical-scavenging effects of Aloe arborescens Miller on prevention of pancreatic islet β-cell destruction in rats. J Ethnopharmacol. 2003;89:37-45. doi:10.1016/S0378-8741(03)00268-X
18. Islam MS, Loots DT. Experimental rodent models of type 2 diabetes: A review. Methods Find Exp Clin Pharmacol. 2009;31:249-261. doi:10.1358/ mf.2009.31.4.1362513, PMid:19557203 [ Links ]
19. Brownlee M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes. 2005;54:1615-1625. doi:10.2337/diabetes.54.6.1615, PMid:15919781 [ Links ]
20. Grove J, Daly AK, Bassendine MF, Day CP. Association of a tumor necrosis factor promoter polymorphism with susceptibility to alcoholic steatohepatitis. Hepatology. 1997;26:143-146. doi:10.1002/hep.510260119, PMid:9214463 [ Links ]
21. Brownlee M. A radical explanation for glucose-induced β cell dysfunction. J Clin Invest. 2003;112:1788-1790. doi:10.1172/JCI20501, PMid:14679173, PMCid:297003
22. Burcelin R, Eddouks M, Maury J, Kande J, Assan R, Girard J. Excessive glucose production, rather than insulin resistance, accounts for hyperglycemia in recent-onset streptozotocin-diabetic rats. Diabetologia. 1995;38:283-290. doi:10.1007/BF00400632, PMid:7758874 [ Links ]
23. Garcia-Compean D, Jaquez-Quintan JO, Maldonado-Gaza H. Hepatogenous diabetes. Current reviews of an ancient problem. Ann Hepatol. 2009;8:13-20. PMid:19221528 [ Links ]
24. Satriano J, Vallon V. Primary kidney growth and its consequences at the onset of diabetes mellitus. Amino Acids. 2006;31:1-9. doi:10.1007/s00726-006-0326-x, PMid:16733619 [ Links ]
25. Valentovic MA, Alejandro N, Carpenter B, Brown A, Ramos K. Streptozotocin (STZ) diabetes enhances benzo(alpha)pyrene induced renal injury in Sprague Dawley rats. Toxicol Lett. 2006;164:214-220. doi:10.1016/j.toxlet.2005.12.009, PMid:16460892 [ Links ]
26. Lugman W, Abdella N, Moro M. Serum fructosamine concentration as measure of blood glucose control in insulin dependent diabetes. Br Med J. 1985;290:1075-1076. doi:10.1136/bmj.290.6474.1075-a, PMid:3921112, PMCid:1418361 [ Links ]
27. Jiao S, Matsuzawa Y, Matsubara K, et al. Increased activity of intestinal acyl-CoA:cholesterol acyltransferase in rats with streptozotocin-induced diabetes and restoration with insulin supplementation. Diabetes. 1988;37:342-346. doi:10.2337/diabetes.37.3.342, PMid:2967215 [ Links ]
28. Kusunoki J, Aragane K, Kitamine T, et al. Postprandial hyperlipidemia in streptozotocin-induced diabetic rats is due to abnormal increase in intestinal acyl coenzyme A: cholesterol acyltransferase activity. Arterioscler Thromb Vasc Biol. 2000;20:171-178. PMid:10634814 [ Links ]
29. Krauss RM. Lipids and lipoproteins in patients with type 2 diabetes. Diabetes Care. 2004;27:1496-1504. doi:10.2337/diacare.27.6.1496, PMid:15161808 [ Links ]
 Correspondence to:
Correspondence to:
Du Toit Loots
Private Bag X6001
Potchefstroom 2531, South Africa
Email: dutoit.loots@nwu.ac.za
Received: 24 Nov. 2010
Accepted: 31 Mar. 2011
Published: 14 July 2011
© 2011. The Authors. Licensee: OpenJournals Publishing. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}