










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.107 n.5-6 Pretoria May./Jun. 2011
http://dx.doi.org/10.4102/sajs.v107i5/6.456
RESEARCH LETTER
DNH 109: A fragmentary hominin near-proximal ulna from Drimolen, South Africa
Andrew Gallagher; Colin G. Menter
Department of Anthropology and Development Studies, University of Johannesburg, Johannesburg, South Africa
ABSTRACT
We describe a fragmentary, yet significant, diminutive proximal ulna (DNH 109) from the Lower Pleistocene deposits of Drimolen, Republic of South Africa. On the basis of observable morphology and available comparative metrics, DNH 109 is definitively hominin and is the smallest African Plio-Pleistocene australopith ulna yet recovered. Mediolateral and anteroposterior dimensions of the proximal diaphysis immediately distal to the m. brachialis sulcus in DNH 109 yield an elliptical area (π/4 *m-l*a-p) that is smaller than the A.L. 333-38 Australopithecus afarensis subadult from Hadar. Given the unusually broad mediolateral anteroposterior diaphyseal proportions distal to the brachialis sulcus, the osseous development of the medial and lateral borders of the sulcus, and the overall size of the specimen relative to comparative infant, juvenile, subadult and adult comparative hominid ulnae (Gorilla, Pan and Homo), it is probable that DNH 109 samples an australopith of probable juvenile age at death. As a result of the fragmentary state of preservation and absence of association with taxonomically diagnostic craniodental remains, DNH 109 cannot be provisionally assigned to any particular hominin genus (Paranthropus or Homo) at present. Nonetheless, DNH 109 increases our known sample of available Plio-Pleistocene subadult early hominin postcrania.
Introduction
The fossil hominin site of Drimolen, currently dated by faunal correlation to between 2.0 MYA and 1.5 MYA, has yielded a wealth of craniodental specimens attributed to Paranthropus robustus (n = 60) and a more modest sample of early Homo (n = 9).1,2,3,4 Most of the Australopithecus, Paranthropus and early Homo hypodigms from the Cradle of Humankind consist of craniodental specimens in varying states of preservation.2,3,4,5,6,7,8,9 Postcranial specimens attributed to P. robustus and early Homo are comparably rare and only three localities - Swartkrans, Kromdraai and Drimolen - have yielded postcranial specimens that are confidently assigned to these taxa.2,3,4,10,11,12,13,14 Morphological and metrical affinities of available postcranial remains have generally failed to reject the hypothesis that a single locomotor 'morph' is represented amongst Lower Pleistocene hominins in South Africa.10,15,16,17,18,19,20,21,22,23,24 We describe here a fragmentary, yet potentially important, near-proximal ulna (DNH 109) recovered from the decalcified sediments in the Main Quarry of Drimolen. DNH 109 was excavated in 1995 from grid square N199, E203 at a height of -2.00 m to -2.10 m below the Drimolen datum.
Materials and methods
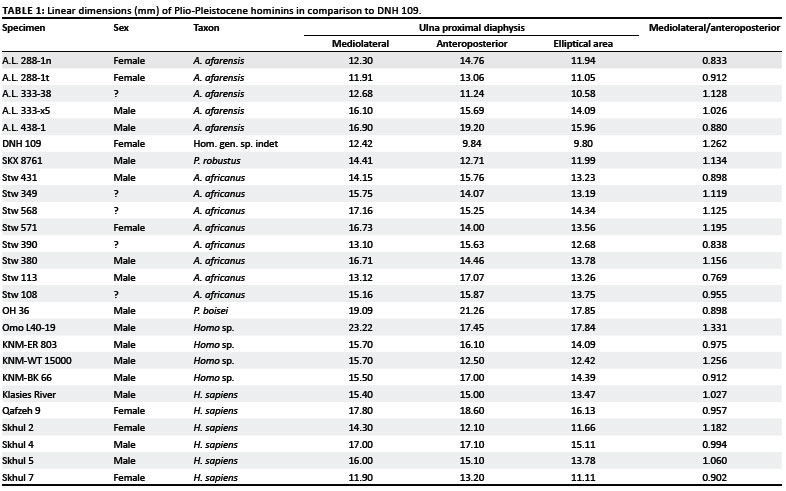
Comparative infant, juvenile and subadult samples of Pan (including P. paniscus,n = 53), Gorilla (n = 29) and Homo (n = 162) and adult samples of P. paniscus (n = 15), P. t. troglodytes (n = 85), G. g. gorilla (n = 99), H. sapiens (n = 118) and Papio (n = 56) were measured at the Cleveland Museum of Natural History (USA), Kent State University (USA), the Natural History Museum (USA), the Powell-Cotton Museum (UK), the University of Dundee (UK), the Royal Museum of Central Africa (Belgium) and the University of the Witwatersrand (SA). The human postnatal growth series comprised individuals aged 0-9 years at death, whereas the Pan and Gorilla series included individuals ranging from newborn to adolescent (prior to epiphyseal fusion). The human sample from the Raymond Dart Collection at the University of the Witwatersrand included South African individuals of European and African ancestry. Original South African fossils and high resolution casts of available East African specimens were measured in the School of Anatomical Sciences, University of the Witwatersrand. Linear dimensions of the proximal ulnar diaphysis were taken at the terminus of the m. brachialis sulcus25,26 using MitutoyoTM digital calipers (Mitutoyo Corporation, Aurora, IL, USA), accurate to the nearest 0.01 mm. Additional published data for A.L. 438-1, KNM-BK 66, KNM-ER 803, KNM-WT 15000, KRM and Skhul/Qafzeh were used in the analysis.26,27
Preservation and morphology of DNH 109
Preservation
DNH 109 is a near-proximal portion of a right diaphysis which is broken distal to the coronoid process (Figure 1). No proximal articulation is preserved. The specimen is missing the olecranon, trochlear notch, radial notch and coronoid process. The shaft fragment at its greatest length is 52.6 mm. The break at the proximal end of the specimen is just below the anterior border of the coronoid process. Inferior to this the ulnar tuberosity and a very small supinator crest are preserved. The ulnar tuberosity is missing a 1-cm long, narrow flake of bone that would form the proximomedial border of the rim of the ulnar tuberosity. There are several hairline cracks running longitudinally through the ulnar tuberosity. The medial rim of the ulnar tuberosity displays a number of small grooves that run from the medial edge onto the posterolateral surface of the shaft, possibly evidence of taphonomic damage by a carnivore 'worrying' the bone. Some small tooth scores are evident. On the anterior surface of the bone just proximal to the distal break a nutrient foramen is evident. Whilst the supinator crest is evident, it is abraded over most of its length.

Morphology
As no proximal articulation is preserved there is little morphology that can be discussed, which makes the taxonomic attribution of this specimen problematic. DNH 109 is both smaller and less robust than the smallest accepted adult Plio-Pleistocene ulna, A.L. 288-1. The ulnar tuberosity or area for attachment of m. brachialis is well-defined, proximodistally elongate and forms an ovoid depression reminiscent of the A. afarensis subadult specimen, A.L. 333-98. The medial (anterior border) and lateral margins of the ulnar tuberosity are less well developed than in A.L. 288-1. The nutrient foramen in DNH 109 lies inferior to the terminus of the brachialis sulcus and is slightly medial to the lateral border, as in the A.L. 288-1n,t (A. afarensis) paired left and right ulnae. The only linear dimensions that can be measured on DNH 109 include the mediolateral and anteroposterior dimensions of the distal margin of the ulnar tuberosity. The linear dimensions of DNH 109 and its calculated elliptical area are at the lower size range of the available Plio-Pleistocene specimens, particularly with respect to the anteroposterior depth (Table 1). Although the supinator crest is slightly abraded, it is clear that it is very weakly expressed; in relation to the fossil hominin sample, it is one of the more weakly expressed supinator crests. The lack of development of the supinator crest suggests that the specimen is a subadult individual. In distal view the shaft is triangular in cross section.
Taxonomic status and age at death
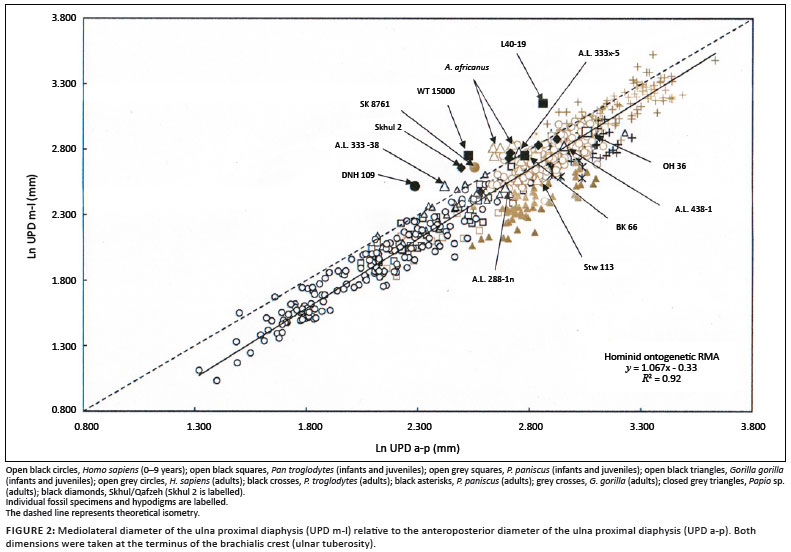
The proximal diaphyseal proportions of DNH 109 are consistent with a taxonomic allocation to the Hominini tribe. The majority of Plio-Pleistocene hominin (Australopithecus and early Homo) ulnae contrast with Pan, Gorilla and more recent humans (ontogenetic and adult, n = 561) in their broad mediolateral proximal diaphysis relative to the anteroposterior depth at the terminus of the brachialis sulcus25,26,27,28,29,30,31,32 (Figure 2), and this proportion is accentuated in certain individuals, including DNH 109, assigned to Australopithecus, Paranthropus, and Homo. The mediolateral/anteroposterior proportions of DNH 109 are almost equivalent to other Plio-Pleistocene African australopiths, yet the anteroposterior diameter in DNH 109 is absolutely small. Whilst DNH 109 is similar to East African early Homo (Omo L-40-19, KNM-WT 15000) with respect to its extreme mediolateral/anteroposterior proportions (Table 1 and Figure 2), it also approaches A.L. 333-38, a putative subadult A. afarensis proximal ulna from Hadar.32 Stw 113 from Sterkfontein Member 4 (STM 4) falls at the lower extremes of the A. africanus distribution by virtue of its narrow mediolateral/anteroposterior proximal diaphyseal proportions (Table 1 and Figure 2), yet clusters with Stw 431, Stw 108, and Stw 390. The mediolateral/anteroposterior proportions of proximal ulnar diaphyses of African late Pliocene australopiths and Lower, Middle and early Upper Pleistocene Homo show considerable variation, within and between genera (A.L. 288-1, Figure 2). As such, no obvious conclusions can be drawn as to the precise taxonomic affinities of DNH 109. The extreme diminution of DNH 109 relative to comparative infant and juvenile hominid ulnae (including P. paniscus) and representative Plio-Pleistocene hominin comparatives (including A.L. 333-38) is consistent with an inference of probable late juvenile or early adolescent age at death.
Conclusion
DNH 109 is definitively hominin and likely represents one of the youngest (ontogenetically) available Plio-Pleistocene hominin ulnae. The absolute small size of DNH 109 is
primarily a function of the extremely shallow anteroposterior depth of the proximal diaphysis at the terminus of the m. brachialis crest. DNH 109 is a potentially important new hominin specimen from Drimolen whose size and proportions may have significance in interpreting the locomotor adaptations of associated craniodental and postcranial juvenile specimens such as DIK-1-1 (A. afarensis) and MH 1 (A. sediba) for which accurate dental developmental age at death estimates are available.33,34
Acknowledgements
We thank D. Smith and J. Smith, on whose property Drimolen is situated, for permission to conduct the excavation and for their continued assistance. Dr J. Hailie-Selassie and L. Jellema (Cleveland Museum of Natural History), Dr C.O. Lovejoy (Kent State University) and Mr M. Harman (Powell-Cotton Museum) provided access to specimens in their care. Original South African fossils were studied at the University of the Witwatersrand and the Northern Flagship Institute. Dr B. Zipfel and Ms S. Potze kindly facilitated the measurement of specimens in their care. The Drimolen excavation is supported by grants to CGM from the National Research Foundation (NRF African Origin Platform, grant number 65155 and a SA-Italy Cooperation agreement, grant number 65155) and the Leakey Foundation. AG is supported by an NRF/University of Johannesburg Postdoctoral Research Fellowship. Financial assistance was also provided by the Ford Foundation and the Faculty of Health Sciences, University of the Witwatersrand.
References
1. Keyser A. The Drimolen skull: The most complete australopithecine cranium and mandible to date. S Afr J Sci. 2000;96:189-193. [ Links ]
2. Keyser A, Menter CG, Moggi-Cecchi J, Pickering TR, Berger LR. Drimolen: A new hominid-bearing site in Gauteng, South Africa. S Afr J Sci. 2000;96:193-197. [ Links ]
3. Herries AIR, Curnoe DC, Adams JW. A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa. Quart Internat. 2009;202:14-28. doi:10.1016/j.quaint.2008.05.017 [ Links ]
4. Moggi-Cecchi J, Menter C, Boccone S, Keyser A. Early hominin dental remains from the Plio-Pleistocene site of Drimolen, South Africa. J Hum Evol. 2010; 58:374-405. doi:10.1016/j.jhevol.2010.01.006, PMid:20362324 [ Links ]
5. Broom R, Robinson JT, Schepers GWH. Sterkfontein ape men. Mem Transv Mus. 1950; No. 4. [ Links ]
6. Clarke RJ. Early Acheulian with H. habilis at Sterkfontein. In: Tobias PV, editor. Hominid evolution: Past, present and future. New York: Alan R Liss, 1985; p. 287-298. [ Links ]
7. Clarke RJ. Australopithecus and early Homo in southern Africa. In: Delson E, editor. Ancestors: The hard evidence. New York: Alan R Liss, 1985; p. 171-177. [ Links ]
8. Lockwood CA, Tobias PV. Morphology and affinities of new hominin cranial remains from member 4 of the Sterkfontein formation, Gauteng province, South Africa. J Hum Evol. 2002;42:389-450. doi:10.1006/jhev.2001.0532, PMid:11908955 [ Links ]
9. Moggi-Cecchi J, Grine FE, Tobias PV. Early hominid dental remains from members 4 and 5 of the Sterkfontein formation (1966-1996 excavations): Catalogue, individual associations, morphological descriptions and initial metrical analyses. J Hum Evol. 2006;50:239-328. doi:10.1016/j. jhevol.2005.08.012, PMid:16309732 [ Links ]
10. Robinson JT. Early hominid posture and locomotion. Chicago: Chicago University Press; 1972. [ Links ]
11. Clarke RJ, Howell FC, Brain CK. More evidence of an advanced hominid at Swartkrans. Nature. 1970;225:1219-1222. doi:10.1038/2251219a0, PMid:5435352 [ Links ]
12. Day MH. A guide to fossil man. London: Cassell; 1986. [ Links ]
13. Wood B, Richmond BG. Human evolution: Taxonomy and paleobiology. J Anat. 2000;196:19-60. doi:10.1046/j.1469-7580.2000.19710019.x, PMid:10999270, PMCid:1468107 [ Links ]
14. Wood B, Lonergan N. The hominin fossil record: Taxa, grades and clades. J Anat. 2008;212:354-376. doi:10.1111/j.1469-7580.2008.00871.x, PMid:18380861, PMCid:2409102 [ Links ]
15. Day MH. Locomotor features of the lower limb in hominids. In: Zuckerman S, editor. The concepts of human evolution. Symp Zool Soc Lond. 1973;33:29-51. [ Links ]
16. Day MH. Bipedalism: Pressures, origins and modes. In: Wood BA, Martin LB, Andrews PA, editors. Major topics in primate and human evolution. Cambridge: Cambridge University Press, 1986; p. 188-202. [ Links ]
17. Day MH, Wood BA. Functional affinities of the Olduvai hominid 8 talus. Man, Second Series. 1968;3:440-455. [ Links ]
18. Oxnard CE. Some African fossil foot bones: A note on the interpolation of fossils into a matrix of extant species. Am J Phys Anthropol. 1972;37:3-12. doi:10.1002/ajpa.1330370103, PMid:5064732 [ Links ]
19. Oxnard CE. Uniqueness and diversity in human evolution: Morphometric studies of the australopithecines. Chicago: Chicago University Press; 1975. [ Links ]
20. Lisowski FP, Albrecht GH, Oxnard CE. African fossil tali: Further multivariate studies. Am J Phys Anthropol. 1974;45:5-18. doi:10.1002/ ajpa.1330450103, PMid:961834 [ Links ]
21. McHenry HM, Corruccini RS. Distal humerus in hominoid evolution. Folia Primatol. 1975;23:227-244. doi:10.1159/000155673 [ Links ]
22. Lague MR, Jungers WL. Morphometric variation in Plio-Pleistocene hominid distal humeri. Am J Phys Anthropol. 1996;101:401-427. doi:10.1002/(SICI)1096-8644(199611)101:3<401::AID-AJPA8>3.0.CO;2-0 [ Links ]
23. Bacon A-M. Principal components analysis of distal humerus shape from Pliocene to recent African hominids: The contribution of geometric morphometrics. Am J Phys Anthropol. 2000;111:479-488. doi:10.1002/ (SICI)1096-8644(200004)111:4<479::AID-AJPA4>3.3.CO;2-R [ Links ]
24. Menter CG. Hominid distal humeri, proximal radii and proximal ulnae from Sterkfontein, in comparison with the elbow bones of other Plio-Pleistocene fossil hominids. PhD thesis, Johannesburg, University of the Witwatersrand, 2003. [ Links ]
25. McHenry HM, Corruccini RS, Howell FC. Analysis of an early hominid ulna from the Omo basin, Ethiopia. Am J Phys Anthropol. 1976;44:295-304. doi:10.1002/ajpa.1330440212, PMid:816207 [ Links ]
26. McHenry HM, Brown CC, McHenry LJ. Fossil hominin ulnae and the forelimb of Paranthropus. Am J Phys Anthropol. 2007;134:209-218. doi:10.1002/ajpa.20656, PMid:17596856 [ Links ]
27. Pearson OM. Postcranial morphology and the origins of modern humans. PhD thesis, New York, Stony Brook State University of New York, 1997. [ Links ]
28. Howell FC, Wood BA. Early hominid ulna from the Omo basin, Ethiopia. Nature. 1974;249:174-176. doi:10.1038/249174a0, PMid:4833521 [ Links ]
29. Day MH. Functional interpretations of the morphology of postcranial remains of early African hominids. In: Jolly CJ, editor. Early hominids of Africa. London: Duckworth, 1978; p. 311-345. [ Links ]
30. Aiello LC, Wood BA, Key C, Lewis M. Morphological and taxonomic affinities of the Olduvai ulna (OH 36). Am J Phys Anthropol. 1999;109:89-110. doi:10.1002/(SICI)1096-8644(199905)109:1<89::AID-AJPA8>3.0.CO;2-4 [ Links ]
31. Drapeau MSM, Ward CV, Kimbel WH, Johanson DC, Rak Y. Associated cranial and forelimb remains attributed to Australopithecus afarensis from Hadar, Ethiopia. J Hum Evol. 2005;48:593-642. doi:10.1016/j. jhevol.2005.02.005, PMid:15927662 [ Links ]
32. Lovejoy CO, Johanson DC, Coppens Y. Hominid upper limb bones recovered from the Hadar formation: 1974-1977 collections. Am J Phys Anthropol. 1982;57:637-649. doi:10.1002/ajpa.1330570409 [ Links ]
33. Alemseged Z, Spoor F, Kimbel WH, et al. A juvenile early hominin skeleton from Dikika, Ethiopia. Nature. 2006;443:296-301. doi:10.1038/nature05047, PMid:16988704 [ Links ]
34. Berger LR, de Ruiter DJ, Churchill SE, et al. Australopithecus sediba: A new species of Homo-like australopith from South Africa. Science. 2010;328:195-204. doi:10.1126/science.1184944, PMid:20378811 [ Links ]
 Correspondence to:
Correspondence to:
Andrew Gallagher
Postal address: PO Box 524
Auckland Park 2006, South Africa
Email: agal1815@gmail.com
Received: 05 Oct. 2010
Accepted: 06 Mar. 2011
Published: 12 May 2011
© 2011. The Authors. Licensee: OpenJournals Publishing. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}