










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.107 n.3-4 Pretoria Mar./Apr. 2011
http://dx.doi.org/10.4102/sajs.v107i3/4.331
RESEARCH ARTICLE
Hominin palaeoecology in late Pliocene Malawi: first insights from isotopes (13C, 18O) in mammal teeth
Hervé BocherensI; Oliver SandrockII; Ottmar KullmerIII; Friedemann SchrenkIV
IFachbereich Geowissenschaften, Biogeologie, Universität Tübingen, Tübingen, Germany
IIGeologisch, Paläontologisch und Mineralogische Abteilung, Hessisches Landesmuseum Darmstadt, Germany
IIIForschungsinstitut Senckenberg, Palaeoanthropologie, Frankfurt am Main, Germany
IVInstitut für Ökologie, Evolution und Diversität, Johann Wolfgang Goethe-Universität, Frankfurt am Main, Germany
ABSTRACT
Carbon-13 and oxygen-18 abundances were measured in large mammal skeletal remains (tooth enamel, dentine and bone) from the Chiwondo Beds in Malawi, which were dated by biostratigraphic correlation to ca. 2.5 million years ago. The biologic isotopic patterns, in particular the difference in carbon-13 abundances between grazers and browsers and the difference in oxygen-18 abundances between semi-aquatic and terrestrial herbivores, were preserved in enamel, but not in dentine and bone. The isotopic results obtained from the skeletal remains from the Chiwondo Beds indicate a dominance of savannah habitats with some trees and shrubs. This environment was more arid than the contemporaneous Ndolanya Beds in Tanzania. The present study confirms that robust australopithecines were able to live in relatively arid environments and were not confined to more mesic environments elsewhere in southern Africa.
Introduction
Links between the environment and key episodes of hominin evolution in Africa have often been suggested.1,2,3,4,5,6 It is therefore of prime importance to have the most accurate knowledge of the environmental conditions at the time and place that hominins were present. Isotopic investigations of mammal remains from Plio-Pleistocene hominin sites have proved highly informative regarding palaeoecosystems in southern and eastern Africa.7,8,9,10,11,12,13,14,15,16,17To date no such investigation has been attempted in Malawi - an area that has yielded hominin fossils important for closing the gap between localities in southern and eastern Africa.18,19,20 A recent study of the mammalian fauna indicated a palaeoecological reconstruction that was relatively arid, with a strong affinity to the Somalia-Masai ecozone, and more arid than it is today.21 However, the study was made difficult by the taphonomic biases of the localities22 leading to the preferential preservation of large species. To help in refining this palaeoecological reconstruction, we present here preliminary isotopic results on mammal remains from unit 3A of the Chiwondo Beds that yielded Homo rudolfensis and Paranthropus boisei.18,23 The fauna from both hominin sites are biostratigraphically correlated to the Plio-Pleistocene boundary, around 2.5 million years ago (MYA), a period for which an aridification event has been tentatively linked to a turnover and a radiation of hominin species, with the first appearance of the genera Paranthropus and Homo.24,25 For almost 50 years, the conventional opinion on this dichotomy of the hominin clade has favoured the hypothesis that the major adaptive differences between the Homo and Paranthropus lineages developed because of the increasing dietary specialisation of Paranthropus as a result of climate change, while Homo adopted a generalist strategy.26 Wood and Strait27 tested the diet/climate hypothesis by analysing a set of criteria and suggested that both genera probably were eurytrope and that derived morphology does not always mirror specialised adaptation towards a particular diet or habitat preference. Therefore it is striking to see how many of the habitat reconstructions for early Homo and Paranthropus draw a mosaic picture, interpreting that both hominin groups existed in variable environments. The goal of this study is to evaluate the state of preservation of the fossil mammal material for carbonate isotopic investigations and to draw some first conclusions on the palaeoenvironment inhabited by these early hominins in south-east Africa in order to improve our knowledge about their environmental distribution and constraints.
Significance of isotopic (palaeo-)ecology
The relative abundance of carbon-13 (13C) in mammalian tissues is directly linked to their average diet. In African tropical environments, two types of vegetation are clearly distinguished by their carbon isotopic signatures: plants using the C3 photosynthetic pathway (trees and shrubs) and plants using the C4 photosynthetic pathway (grasses). As herbivores record the carbon isotopic signature of their plant food, analysing their fossil remains allows the reconstruction of some aspects of the vegetation in their preferred habitat, as long as the isotopic signatures have not been significantly modified post-mortem. Some species exhibit a restricted dietary spectrum and can be used as tracers of possible diagenetic alteration.28 For example, giraffes and the proboscidian Deinotherium are exclusively browsers and therefore exhibit C3-consumer isotopic signatures, while the Alcelaphini antelopes exclusively consume C4 grasses.10,29,30,31,32 In contrast, other species are more opportunistic and their carbon isotopic signatures can be used to evaluate the relative importance of C3 and C4 plants available to them. For instance, hippopotamuses consume different amounts of C3 plants depending on their availability.8,33,34,35
Oxygen-18 (18O) abundances in skeletal tissues are linked to those of drinking water and water contained in the food, and they vary according to temperature and aridity. They typically increase with increasing temperature and decreasing precipitation.36,37 Different habitats and water conservation strategies between herbivores lead to coherent patterns in the oxygen isotopic signatures of African herbivores, with giraffes systematically 18O-enriched compared to other terrestrial herbivores11,38 and hippopotamuses systematically 18O-depleted compared to terrestrial herbivores.8,34,39 These biogenic patterns can also be used to track possible diagenetic alteration in fossil material. The variations of δ18O values of a given herbivorous species through space and time can be tentatively related to changes in climatic conditions, but caution is necessary in the interpretation of small datasets as terrestrial mammals typically exhibit significant variation of their δ18O values within a living population.39
Materials and methods
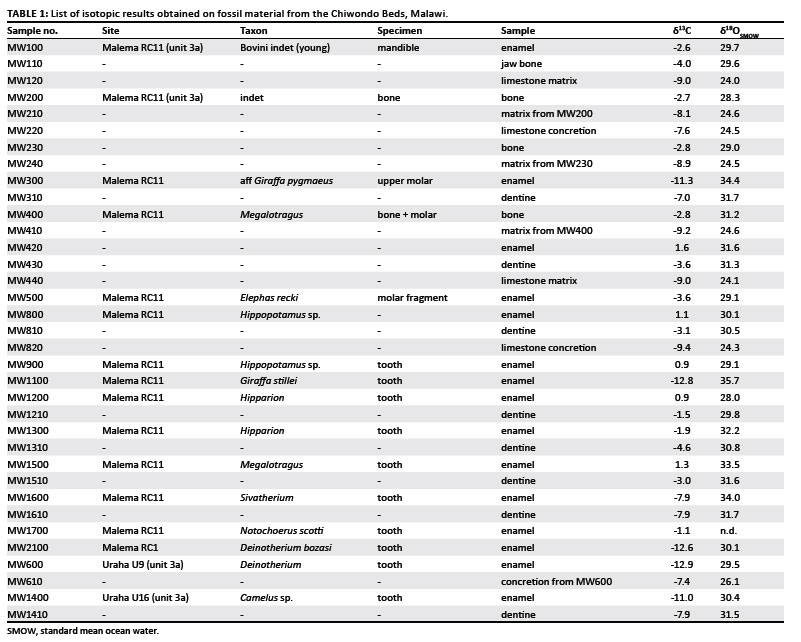
All specimens studied were derived from unit 3A-2 in the Chiwondo Beds (Malawi), either from Malema RC11 (12 specimens) or from Uraha (2 specimens). Biozone 2 in unit 3A is relatively dated by the correlation of suid remains with other radiometric dated sites in eastern and South Africa to between 2.5 MYA and 2.3 MYA. Most of the material comes from the Malema excavation site RC11 from the same level that yielded the Paranthropus maxilla fragment HCRP-RC-911.20,21 Tooth or jawbone fragments with teeth have been selected from the following taxa: proboscidians (Deinotherium sp., Elephas recki), giraffids (Giraffa sp., Sivatherium sp.), bovids (one undetermined specimen, Megalotragus sp.), camel (Camelus sp.), Hippopotamus sp. and equids (Hipparion sp.). Small enamel fragments were retrieved, as well as fragments of dentine and bone for some specimens (Table 1). Whenever possible, the calcareous matrix was also sampled. Comparing enamel, dentine, bone and limestone isotopic signatures allows evaluation of possible diagenetic alteration of the isotopic signatures.40,41,42
A chemical pre-treatment aimed at the removal of exogenous contaminants, such as organic residues and carbonates, was applied to the enamel, dentine and bone samples.8 Powdered samples were soaked in 2-3% NaOCl for 20 h at 20 ºC to oxidise organic residues, rinsed with distilled water, then treated with 1 M acetic acid-Ca acetate buffer (pH = 4.75) for 20 h at 20 ºC to remove exogenous carbonate. Carbon dioxide was produced from the treated powders (± 15 mg) by dissolution in 100% H3PO4 at 50 ºC for 5 h. Carbon dioxide was collected and purified by cryogenic distillation in a vacuum line, and carbon isotope compositions were measured on a VG Optima gas source mass spectrometer (Manchester, UK). The isotopic ratios are expressed as δ13C and δ18O values with an analytical precision better than 0.1‰ and 0.2‰, respectively; the international standards are PeeDee belemnite (PDB) and standard mean ocean water (SMOW) for δ13C and δ18O values, respectively.8
Results
The carbon and oxygen isotopic signatures of enamel exhibited a much larger variation than those of dentine, bone and embedding limestone (Table 1, Figure 1). The δ13C values ranged from -12.9‰ to +1.6‰ in enamel, from -7.9‰ to -1.5‰ in dentine, from -4.0‰ to -2.7‰ in bone and from -9.4‰ to -7.4‰ in embedding limestone. A similar pattern was observed for δ18O values: they ranged from 28.0‰0 to 35.7‰ in enamel, from 29.8‰ to 31.7‰ in dentine, from 28.3‰ to 31.2‰ in bone and from 24.0‰ to 26.1‰ in embedding limestone.

The isotopic signatures of fossil enamel were clearly related to the specific origin of the samples. The browsing taxa, such as Giraffa and Deinotherium, clearly exhibited δ13C values indicating exclusive consumption of C3 plants (-12.9‰ to -11.3‰), while Megalotragus, a specialised grazer, exhibited δ13C values typical of exclusive C4-plant consumption (1.3‰ to 1.6‰). The variations of the δ18O values were also related to taxonomic affinity of the samples: the Giraffa samples exhibited the most positive δ18O values (34.4‰ and 35.7‰) while Hippopotamus specimens exhibited slightly less positive δ18O values (29.1‰ and 30.1‰).
Discussion
Effects of diagenetic alteration and preservation of enamel isotopic signals
The predicted biogenic patterns in δ13C values were found for the enamel isotopic signatures in exclusive browsers such as Giraffa and Deinotherium, in comparison to grazers such as Megalotragus. Giraffids exhibited more positive δ18O values than other herbivores and Hippopotamus exhibited lower δ18O values than the average of terrestrial herbivores. In contrast, the isotopic signatures of dentine and bone from the same specimens did not display such a clear trend. For instance the δ13C values tended to gather around intermediate values ranging from -8‰ to -1‰, instead of clustering according to the predicted dietary preferences (Figure 1). Interestingly, the δ13C values of carbonate from the surrounding matrix and concretions showed δ13C values between -10‰ and -7‰, which suggests that the δ13C values of dentine were influenced by this background carbon, especially for grazers that tended to shift towards these values. A similar pattern was visible for the δ18O values, in which the differences related to taxonomic affiliation exhibited in enamel were not reflected in bone and dentine (Figure 1). This pattern suggests that the δ18O values of bone and dentine were also affected by diagenetic alteration, in contrast with those of enamel. These results are similar to those published by Lee-Thorp28 on South African material. In this study, there was no evidence that the enamel isotopic ratios were significantly altered by diagenesis.
Palaeoenvironmental reconstruction of the Chiwondo Beds
Although the small number of individuals analysed per species does not allow statistically based inferences, the isotopic patterns seen in various taxa will be used to draw preliminary palaeoenvironmental conclusions. An interesting pattern is observed in the giraffid Sivatherium, which exhibited less negative δ13C values than the purely browsing Giraffa, -7.9‰ compared to -11.3‰ to -12.8‰ (Table 1 and Figure 2). This fossil genus has been interpreted as a mixed feeder based on hypsodonty and tooth wear,43 and it seems that the studied individual from Malema incorporated some C4 grass in its diet. This was, however, not the case for Sivatherium marusium from Makapansgat member 3 dated to 3 MYA, that exhibited δ13C values similar to those of Giraffa in the same site.15 This difference could indicate that Malema is a more open environment than the South African site, Makapansgat.

Both Hippopotamus analysed in this study exhibited high δ13C values, indicating a purely C4 diet. Such values are reached in modern Hippopotamus living in open surroundings without closed forests.33 These isotopic values suggest a predominance of open environments dominated by C4 plants.
In conclusion, the environmental reconstruction that can be tentatively drawn from the available isotopic data suggests a dominance of savannah habitats, based on the δ13C values of Hippopotamus and possibly Sivatherium, with some trees and shrubs yielding C3 biomass present, as indicated by the low δ13C values of Deinotherium, Camelus and Giraffa.
Regional comparison
A preliminary comparison is possible with contemporaneous sites in eastern and southern Africa; specifically the Ndolanya Beds at Laetoli (Tanzania) and Swartkrans 1 (South Africa), for which the carbon and oxygen isotopic signatures of fossil material have been published.11,44 Comparison of the isotopic data with Swartkrans is made difficult by the fact that δ18O values are not published and the taxa are different from those in the Chiwondo Beds. Nevertheless, the bovid assemblage in general indicates more arid conditions at Swartkrans than at Sterkfontein,45,46 suggesting a less woodland and more open habitat at Swartkrans - a conclusion also supported by a comparison of the micromammal fauna47 and the stable carbon isotopes of fossil mammals48 from these cave sites. In contrast, there are common taxa in the Chiwondo Beds and Ndolanya Beds, such as Sivatherium, Alcelaphini, Hipparion (=Eurygnathohippus), elephantids (Loxodonta cf. exoptata and Elephas recki, respectively) providing δ13C values that can be compared between both sites (Figure 3). Elephantids and Hipparion do not exhibit a clear pattern for δ13C and δ18O values, but for Alcelaphini and Sivatherium, the δ13C values appear slightly more positive in the Chiwondo Beds and the δ18O clearly more positive. This could reflect more arid conditions for the Chiwondo Beds than for the Ndolanya Beds. The latter has been interpreted as semi-arid bushland,49 comprising a faunal mixture of 75% from bushland and 25% from grassland habitats50 and corresponding to an area dominated by wooded grassland, ranging from limited areas of closed woodland to much more extensive areas of open bushland and grassland.51 At present, the preliminary results on Sivatherium, present in both sites, suggest more arid conditions in south-eastern than in eastern Africa.
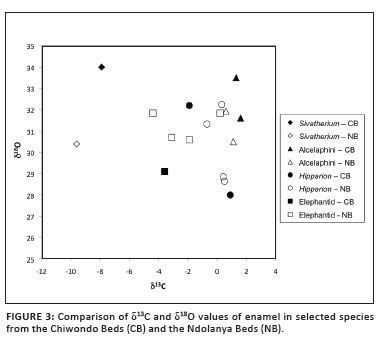
Implications for hominin palaeoecology
The discoveries of Paranthropus aethiopicus in the Ndolanya Beds,52 and of P. boisei in the Chiwondo Beds19 imply that the eastern African robust australopithecines could adapt to open bushland and grassland conditions. This finding is quite striking because the Laetoli area has never been adjacent to a lake, while the Chiwondo Beds' fauna was associated with a lake throughout most of its succession. Wood and Constantino53 summarised the combined evidence of locality reconstructions and indicated that P. boisei favours grassland and/or open woodland habitats. Although Shipman and Harris54 placed P. boisei towards the closed habitat spectrum, Wood and Constantino53 and Wood and Strait27 argue that P. boisei possibly had a broad ecological tolerance. According to Reed55, P. robustus appears to be slightly more arid adapted than eastern African P. boisei. In any case, the appearance of P. boisei in the Chiwondo Beds about 2.5 MYA suggests ecological conditions that may be comparable to the ones of A. robustus in South Africa after 2 MYA.
Interestingly, carbon isotopic data have been recently published for P. boisei from Tanzania, at Olduvai East and Peninj.13 In contrast with the robust australopithecines from South Africa that present δ13C values indicative of limited amounts of C4 biomass in their diet, those from eastern Africa exhibit δ13C values of about -1.0‰, indicating a C4 biomass in their diet of up to 80%. It remains unclear if this very high C4 component was acquired directly through C4 grasses, especially their seeds (though these are highly seasonal resources), or indirectly through the consumption of the flesh of herbivores that consumed C4 plants, other small mammals or invertebrates, such as termites.55,56 Another possibility is the consumption of sedges such as papyrus, a semi-aquatic plant that bears perennial underground storage organs available to hominins all year round.13,57
The geographic position of the Malawi Rift in south-eastern Africa is of special interest when comparing the dietary adaptations of east and South African robust australopithecines. Ungar et al.58 showed that the diet of P. boisei included similar amounts of tough (or fibrous) foods to that of A. africanus. In contrast, the food of P. robustus was much harder (including seeds, nuts and hard fruits) but less tough (i.e. there were fewer fibrous foods such as leaves) than that of P. boisei. Additional isotopic data may prove whether this distinction is as a result of different diets or different feeding strategies.
Conclusion
The isotopic preservation of tooth enamel carbonate fraction in the fossil mammals of the Chiwondo Beds was sufficient to allow preliminary palaeoenvironmental reconstruction. This reconstruction, based on the analysis of δ13C and δ18O values of large herbivorous mammal tooth enamel, was consistent with that based on the analysis of the faunal community.20,21,59The environment was dominated by a savannah ecosystem with some trees, and was probably drier than the modern ecosystem in this region. The environment in the Chiwondo Beds was possibly drier than in Tanzanian sites at the same period, about 2.5 MYA, but additional isotopic data are necessary to confirm this trend.
The present study supports the view that P. boisei was able to live in arid environments and underlines the broad ecological spectrum of this hominin.
Acknowledgements
We acknowledge the technical assistance of D. Billiou and A. Zazzo in the Department of Earth Sciences of University Paris 6 (France) in the preparation and isotopic analysis of the specimens. We also thank the Department of Antiquities, Lilongwe and the Cultural and Museum Centre Karonga for permitting access to the material. We express our gratitude to W. Joyce for his help in editing this manuscript, and to A. Morris and three anonymous reviewers for the suggested improvements to this manuscript.
References
1. Foley RA. Speciation, extinction and climatic change in hominid evolution. J Hum Evol. 1994;26:275-289. doi:10.1006/jhev.1994.1017 [ Links ]
2. Vrba ES. Ecological and adaptive changes associated with early hominid evolution. In: Delson E, editor. Ancestors: The hard evidence. New York: Alan R Liss Inc., 1985; p. 63-71. [ Links ]
3. Potts R. Environmental hypotheses of Pliocene human evolution. In: Bobe R, Alemseged Z, Behrensmeyer AK, editors. Hominin environments in the East African Pliocene: An assessment of the faunal evidence. Dordrecht: Springer, 2007; p. 25-49. doi:10.1007/978-1-4020-3098-7_2 [ Links ]
4. Schrenk F, Kullmer O, Sandrock O, Bromage TG. Early hominid diversity, age and biogeography of the Malawi-Rift. Hum Evol. 2002;17:113-122. doi:10.1007/BF02436432 [ Links ]
5. Schrenk F, Sandrock O, Kullmer O. The "open source" perspective of earliest hominid origins. Collegium Anthropologicum. 2004;28:113-119. PMid:15571086 [ Links ]
6. Trauth MH, Maslin MA, Deino AL, Strecker MR, Bergner AGN, Dühnforth M. High- and low-latitude forcing of Plio-Pleistocene East African climate and human evolution. J Hum Evol. 2007;53:475-486. doi:10.1016/j.jhevol.2006.12.009, PMid:17959230 [ Links ]
7. Lee-Thorp JA, van der Merwe NJ. Carbon isotope studies of Swartkrans fauna and hominids. Transvaal Mus Mon. 1993;8:251-256. [ Links ]
8. Bocherens H, Koch PL, Mariotti A, Geraads D, Jaeger J-J. Isotopic biogeochemistry (13C, 18O) of mammal enamel from African Pleistocene hominid sites: Implications for the preservation of paleoclimatic isotopic signals. Palaios. 1996;11:306-318. doi:10.2307/3515241 [ Links ]
9. Lee-Thorp JA, Thackeray JF, van der Merwe NJ. The hunters and the hunted revisited. J Hum Evol. 2000;39:565-576. doi:10.1006/jhev.2000.0436, PMid:11102267 [ Links ]
10. Sponheimer M, Lee-Thorp JA, de Ruiter D, et al. Diets of southern African Bovidae: Stable isotope evidence. J Mammal. 2003;84:471-479. doi:10.1644/1545-1542(2003)084<0471:DOSABS>2.0.CO;2 [ Links ]
11. Kingston JD, Harrison T. Isotopic dietary reconstructions of Pliocene herbivores at Laetoli: Implications for early hominin paleoecology. Palaeogeogr Palaeoclimatol Palaeoecol. 2007;243:272-306. doi:10.1016/j.palaeo.2006.08.002 [ Links ]
12. Levin NE, Simpson SW, Quade J, Cerling TE, Frost SR. Herbivore enamel carbon isotopic composition and the environmental context of Ardipithecus at Gona, Ethiopia. Geol Soc Am Spec Pap. 2008;446:215-234. [ Links ]
13. Van der Merwe NJ, Masao FT, Bamford MK. Isotopic evidence for contrasting diets of early hominins Homo habilis and Australopithecus boisei of Tanzania. S Afr J Sci. 2008;104:153-155. [ Links ]
14. Plummer TW, Ditchfield PW, Bishop LC, et al. Oldest evidence of toolmaking hominins in a grassland-dominated ecosystem. PLoS ONE. 2009;4(9):e7199. doi:10.1371/journal.pone.0007199, PMid:19844568, PMCid:2746317 [ Links ]
15. Sponheimer M, Lee-Thorp JA. Biogeochemical evidence for the environments of early Homo in South Africa. In: Grine FE, Fleagle JG, Leakey RE, editors. The first humans: Origin and early evolution of the genus Homo. Dordrecht: Springer, 2009; p. 185-194. doi:10.1007/978- 1-4020-9980-9_16 [ Links ]
16. White TD, Ambrose SH, Suwa G, et al. Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science. 2009;326:87-93. doi:10.1126/science.1175822 [ Links ]
17. Bedaso Z, Wynn JG, Alemseged Z, Geraads D. Paleoenvironmental reconstruction of the Asbole fauna (Busidima Formation, Afar, Ethiopia) using stable isotopes. Geobios. 2010;43:165-177. doi:10.1016/j.geobios.2009.09.008 [ Links ]
18. Kullmer O, Sandrock O, Schrenk F, Bromage TG. The Malawi Rift: Biogeography, ecology and coexistence of Homo and Paranthropus. J Sci Man Anthropol. 1999;37:221-231. [ Links ]
19. Kullmer O, Sandrock O, Abel R, Schrenk F, Bromage TG, Juwayeyi YM. The first Paranthropus from the Malawi Rift. J Hum Evol. 1999;37:121-127. doi:10.1006/jhev.1999.0308, PMid:10375478 [ Links ]
20. Kullmer O. The Plio-Pleistocene suids (Suidae; Artiodactyla) of the Chiwondo Beds, northern Malawi. J Vert Paleontol. 2008;28:208-216. doi:10.1671/0272-4634(2008)28[208:TFSFTP]2.0.CO;2 [ Links ]
21. Sandrock O, Kullmer O, Schrenk F, Juwayeyi YM, Bromage TG. Fauna, taphonomy, and ecology of the Plio-Pleistocene Chiwondo Beds, northern Malawi. In: Bobe R, Alemseged Z, Behrensmeyer AK, editors. Hominin environments in the East African Pliocene: An assessment of the faunal evidence. Dordrecht: Springer, 2007; p. 315-332. doi:10.1007/978-1-4020-3098-7_12 [ Links ]
22. Sandrock O, Dauphin Y, Kullmer O, Abel R, Denys C. Malema: Preliminary taphonomic analysis of an African hominid locality. C R Acad Sci Paris. 1999;328:133-139. [ Links ]
23. Schrenk F, Bromage TG, Betzler CG, Ring U, Juwayeyi YM. Oldest Homo and Pliocene biogeography of the Malawi Rift. Nature. 1993;365:833-836. doi:10.1038/365833a0, PMid:8413666 [ Links ]
24. Kimbel W. Hominid speciation and Pliocene climatic change. In: Vrba ES, Denton GH, Partridge TC, Burckle LH, editors. Paleoclimate and evolution, with emphasis on human origins. New Haven: Yale University Press, 1995; p. 425-437. [ Links ]
25. Bobe R, Behrensmeyer AK. The expansion of grassland ecosystems in Africa in relation to mammalian evolution and the origin of the genus Homo. Palaeogeogr Palaeoclimatol Palaeoecol. 2004;207:399-420. doi:10.1016/S0031-0182(04)00049-5 [ Links ]
26. Robinson JT. The genera and species of the Australopithecinae. Am J Phys Anthrop. 1954;12:181-200. doi:10.1002/ajpa.1330120216, PMid:13188956 [ Links ]
27. Wood B, Strait D. Patterns of resource use in early Homo and Paranthropus. J Hum Evol. 2004;46:119-162. doi:10.1016/j. jhevol.2003.11.004, PMid:14871560 [ Links ]
28. Lee-Thorp JA. Stable carbon isotopes in deep time: The diets of fossil fauna and hominids. PhD thesis, Cape Town, University of Cape Town, 1989. [ Links ]
29. Cerling TE, Harris JM, Leakey MG. Browsing and grazing in elephants: The isotope record of modern and fossil proboscideans. Oecologia. 1999;120:364-374. doi:10.1007/s004420050869 [ Links ]
30. Cerling TE, Harris JM, Passey BH. Diets of east African Bovidae based on stable isotopic analysis. J Mammal. 2003;84:456-470. doi:10.1644/1545-1542(2003)084<0456:DOEABB>2.0.CO;2 [ Links ]
31. Codron D, Codron J, Lee-Thorp JA, et al. Diets of savanna ungulates from stable carbon isotope composition of faeces. J Zool. 2007;273:21-29. doi:10.1111/j.1469-7998.2007.00292.x [ Links ]
32. Bocherens H, Jacques L, Ogle N, et al. Reply to the comment by A. Zazzo, W.P Patterson and T.C. Prokopiuk on 'Implications of diagenesis for the isotopic analysis of Upper Miocene large mammalian herbivore tooth enamel from Chad' by Jacques et al. in Palaeogeogr Palaeoclimatol Palaeoecol. 2008;266:200-210. Palaeogeogr Palaeoclimatol Palaeoecol. 2009;277:269-271. [ Links ]
33. Boisserie JR, Zazzo A, Merceron G, et al. Diets of modern and late Miocene hippopotamids: Evidence from carbon isotope composition and microwear of tooth enamel. Palaeogeogr Palaeoclimatol Palaeoecol. 2005;221:153-174. doi:10.1016/j.palaeo.2005.02.010 [ Links ]
34. Cerling TE, Harris JM, Hart JA, et al. Stable isotope ecology of the common hippopotamus. J Zool. 2008;276:204-212. doi:10.1111/j.1469-7998.2008.00450.x [ Links ]
35. Brachert TC, Brügmann GB, Mertz DF, et al. Stable isotope variation in tooth enamel from Neogene hippopotamids: Monitor of meso and global climate and rift dynamics on the Albertine Rift, Uganda. Int J Earth Sci. (Geol Rundsch). 2010;99:1663-1675. doi:10.1007/s00531-010-0518-1 [ Links ]
36. Fricke HC, O'Neil JR. The correlation between 18O/16O ratios of meteoric water and surface temperature: Its use in investigating terrestrial climate change over geologic time. Earth Planet Sci Lett. 1999;170:181-196. doi:10.1016/S0012-821X(99)00105-3 [ Links ]
37. Levin NE, Cerling TE, Passey BH, Harris JM, Ehleringer JR. A stable isotope aridity index for terrestrial environments. Proc Natl Acad Sci USA. 2006;103:11201-11205. doi:10.1073/pnas.0604719103, PMid:16840554, PMCid:1544065 [ Links ]
38. Quade J, Cerling TE, Andrews P, Alpagut B. Paleodietary reconstruction of Miocene faunas from Pasalar, Turkey using stable carbon and oxygen isotopes of fossil tooth enamel. J Hum Evol. 1995;28:373-384. doi:10.1006/jhev.1995.1029 [ Links ]
39. Clementz MT, Holroyd PA, Koch PL. Identifying aquatic habits of herbivorous mammals through stable isotope analysis. Palaios. 2008;23:574-585. doi:10.2110/palo.2007.p07-054r [ Links ]
40. Lee-Thorp JA, van der Merwe NJ. Aspects of the chemistry of modern and fossil biological apatites. J Archaeol Sci. 1991;18:343-354. doi:10.1016/0305-4403(91)90070-6 [ Links ]
41. Bocherens H, Sen S. Pliocene vertebrate locality of Çalta, Ankara, Turkey. 11. Isotopic investigation. Geodiversitas. 1998;20:487-495. [ Links ]
42. Lee-Thorp JA, Sponheimer M. Three case studies used to reassess the reliability of fossil bone and enamel isotope signals for paleodietary studies. J Anthropol Archaeol. 2003;22:208-216. doi:10.1016/S0278-4165(03)00035-7 [ Links ]
43. Franz-Odendall TA, Solounias N. Comparative dietary evaluations of an extinct giraffid (Sivatherium hendeyi) (Mammalia, Giraffidae, Sivatheriinae) from Langebaanweg, South Africa (early Pliocene). Geodiversitas. 2004;26:675-685. [ Links ]
44. Lee-Thorp JA, van der Merwe NJ, Brain CK. Diet of Australopithecus robustus at Swartkrans from stable carbon isotopic analysis. J Hum Evol. 1994;27:361-372. doi:10.1006/jhev.1994.1050 [ Links ]
45. Vrba ES. Chronological and ecological implications of the fossil Bovidae at the Sterkfontein Australopithecine site. Nature. 1974; 250:19-23. doi:10.1038/250019a0 [ Links ]
46. Vrba ES. Some evidence of chronology and paleoecology of Sterkfontein, Swartkrans and Kromdraai from the fossil Bovidae. Nature. 1975; 254:301-304. doi:10.1038/254301a0 [ Links ]
47. Avery DM. The Plio-Pleistocene vegetation and climate of Sterkfontein and Swartkrans, South Africa, based on micromammals. J Hum Evol. 2001;41:113-132. doi:10.1006/jhev.2001.0483, PMid:11437522 [ Links ]
48. Lee-Thorp JA, Sponheimer M, Luyt J. Tracking changing environments using stable carbon isotopes in fossil tooth enamel: An example from the South African hominin sites. J Hum Evol. 2007;53:595-601. doi:10.1016/j. jhevol.2006.11.020, PMid:17920103 [ Links ]
49. Kovarovic K, Andrews P, Aiello L. The palaeoecology of the Upper Ndolanya Beds at Laetoli, Tanzania. J Hum Evol. 2002;43:395-418. doi:10.1006/jhev.2002.0580, PMid:12234550 [ Links ]
50. Andrews P. Taphonomic effects of faunal impoverishment and faunal mixing. Palaeogeogr Palaeoclimatol Palaeoecol. 2006;241:572-589. doi:10.1016/j.palaeo.2006.04.012 [ Links ]
51. Kovarovic K, Andrews P. Bovid postcranial ecomorphological survey of the Laetoli paleoenvironment. J Hum Evol. 2007;52:663-680. doi:10.1016/j. jhevol.2007.01.001, PMid:17353031 [ Links ]
52. Harrison T. The first record of fossil hominins from the Ndolanya Beds, Laetoli, Tanzania. Am J Phys Anthropol. 2002;34(Suppl):83. [ Links ]
53. Wood B, Constantino P. Paranthropus boisei: Fifty years of evidence and analysis. Am J Phys Anthropol. 2007;134(S45):106-132. doi:10.1002/ajpa.20732, PMid:18046746 [ Links ]
54. Shipman P, Harris JM. Habitat preference and paleoecology of Australopithecus boisei in Eastern Africa. In: Grine FE, editor. Evolutionary history of the "robust" australopithecines. New York: Aldine de Gruyter, 1988; p. 343-381. [ Links ]
55. Reed K. Early hominid evolution and ecological change through the African Plio-Pleistocene. J Hum Evol. 1997;32:289-322. doi:10.1006/jhev.1996.0106, PMid:9061560 [ Links ]
56. Peters CR, Vogel JC. Africa's wild C4 plant foods and possible early hominid diets. J Hum Evol. 2005;48:219-236. doi:10.1016/j.jhevol.2004.11.003, PMid:15737391 [ Links ]
57. Sponheimer M, Lee-Thorp JA, de Ruiter D, et al. Hominins, sedges, and termites: New carbon isotope data from the Sterkfontein valley and Kruger National Park. J Hum Evol. 2005;48:301-312. doi:10.1016/j. jhevol.2004.11.008, PMid:15737395 [ Links ]
58. Ungar PS, Grine FE, Teaford MF. Dental microwear and diet of the Plio-Pleistocene hominin Paranthropus boisei. PLoS One. 2008;3(4):e2044. doi:10.1371/journal.pone.0002044, PMid:18446200, PMCid:2315797 [ Links ]
59. Frost S, Kullmer O. Cercopithecidae from the Pliocene Chiwondo Beds, Malawi-rift. Geobios. 2008;41:743-749. doi:10.1016/j.geobios.2008.01.005 [ Links ]
 Correspondence to:
Correspondence to:
Hervé Bocherens
Postal address:
Fachbereich Geowissenschaften, University Tübingen
Sigwartstr. 10
72076 Tübingen, Germany
email: herve.bocherens@uni-tuebingen.de
Received: 28 June 2010
Accepted: 20 Dec. 2010
Published: 07 Mar. 2011
© 2011. The Authors. Licensee: OpenJournals Publishing. This work is licensed under the Creative Commons Attribution License.


{kind=link}