










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.107 n.3-4 Pretoria Mar./Apr. 2011
http://dx.doi.org/10.4102/sajs.v107i3/4.291
RESEARCH ARTICLE
Lake St Lucia, Africa's largest estuarine lake in crisis: Combined effects of mouth closure, low levels and hypersalinity
Digby CyrusI; Hendrik JerlingI; Fiona MacKayII; Leon VivierI
ICoastal Research Unit of Zululand, Department of Zoology, University of Zululand, KwaZulu-Natal, South Africa
IIOceanographic Research Institute, Durban, KwaZulu-Natal, South Africa
ABSTRACT
The Lake St Lucia estuarine system is the most important nursery ground for juvenile marine fish and prawns along the KwaZulu-Natal coast. The estuary mouth closed in June 2002 because of drought and remained so for 4 years and 9 months. A study to determine the impacts of extended mouth closure, hypersalinity and low lake levels on the mesozooplankton, macrobenthic invertebrates and fish fauna was initiated in 2004. Zooplankton and benthic invertebrate diversity declined, benthic invertebrate community composition changed and the diversity and abundance of fish decreased between 2004 and 2007. In the case of fish, the declines were related to die-offs in the lake and the failed recruitment of post-larvae and juveniles from the marine environment as a result of the mouth having been closed. Options for management intervention under closed-mouth conditions are limited at this time, particularly in the short term, to breaching the mouth and facilitating the inflow of sea water. In the medium term, as was the historical situation, the reconnection of the Mfolozi system to St Lucia should be viewed as a major priority.
Introduction
Drought conditions that started in 2002 resulted in the closure, in June, of the mouth of the Lake St Lucia estuarine system, on the south-east coast of Africa, which lasted 4 years and 9 months. Lake St Lucia is the most important estuarine system on the south-east coast of southern Africa, comprising approximately 80% of the estuarine area of KwaZulu-Natal, South Africa.1 The system forms part of the iSimangaliso Wetland Park, a designated World Heritage Site. Originally forming a common mouth with the Mfolozi River,2 it has a history of natural and human-induced disturbance. Because of the expansion of agriculture in the Mfolozi Catchment and the associated increase in sediment runoff and accumulation in the St Lucia/Mfolozi Estuary as a result of the loss of filtering capacity by the Mfolozi swamps,1,2 a decision was taken to separate the two systems3 in 1952.
Lake St Lucia naturally experiences large fluxes in physico-chemical characteristics, both temporally and spatially,1,4 as a result of climatic events such as floods and droughts. During droughts, reductions in freshwater flow have resulted in periods of mouth closure5 with salinities that have reached as high as 102 parts per thousand (ppt)6. These reductions have been seriously exacerbated by human interventions such as the separation of the Mfolozi from St Lucia. Several ecological 'states', from fresh through estuarine and marine to hypersaline, may occur in the lake system at different times, with the marine-estuarine being the dominant state.7
Lake St Lucia underwent major changes over the 5 years following mouth closure in 2002 as a result of the drought, exacerbated by human-related decreases in freshwater inflow. High evaporation rates further reduced water levels, and during the 4 years and 9 months of this study there was no link between the lake and the sea. In March 2007, a combination of equinox high tides driven higher by strong onshore winds and very high seas caused by Cyclone Gamede caused the mouth to breach. The system remained open for only 6 months before closing again in August 2007. During the initial closed period the lake reached its lowest level ever recorded in July 2006, with water only covering an estimated 10% of the lake's 325 km2 surface area whilst salinities reached an all time high of 200 ppt towards the end of 2003.
Lack of key information regarding the fauna under previous extreme conditions in the system prompted the initiation of surveys of the fish, zooplankton and macrobenthic invertebrates. We reviewed the impacts of hypersaline and low lake level conditions resulting from the mouth closure in June 2002 until the breaching in March 2007, on the mesozooplankton, macrobenthos and fish and the functioning of the St Lucia system as a nursery for marine fish and prawns.
Materials and methods
Study area
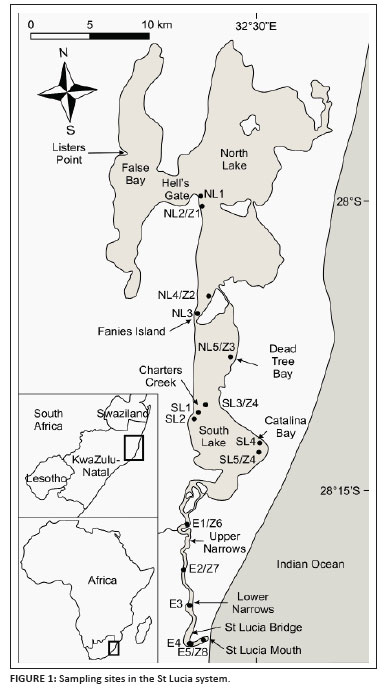
Lake St Lucia (mouth at 28º15'S 32º30'E) covers 325 km2, and is the largest estuarine system in Africa (Figure 1). The system is shallow (the normal average depth is 0.90 m). Our sampling focused on the upper and lower reaches of the estuary (referred to as the Upper and Lower Narrows), the eastern and western regions of South Lake, and Fanies Island (Site NL3) and Hell's Gate (Site NL1) (Figure 1).
Salinity
Weekly salinity measurements were recorded in situ at Charters Creek, Listers Point and St Lucia Bridge (Figure 1) by Ezemvelo KZN Wildlife staff using an optical salinometer (Atago, Tokyo, Japan).
Zooplankton
Mesozooplankton sampling took place during March 2005, May 2006 and November 2006. Samples were collected from eight sites (Figure 1: Z1 to Z8). This was undertaken during daytime as logistical issues, particularly navigation through the shallow system and the presence of crocodiles and hippopotami, precluded night sampling.
Samples were collected using a double plankton net, each 2 m long, with a mouth diameter of 300 mm and a 200-µm mesh. One net was fitted with a flow meter to quantify the samples. The nets were towed in mid-water using a small, motorised boat at a slow speed for about 3 min. Sites Z1 and Z3 were very shallow and could not be sampled with the procedure described above, so water samples (>100 L) were collected at these sites and filtered through the plankton net.
Samples were preserved in approximately 10% formaldehyde solution in estuarine water. In the laboratory a sub-sampling method8 was used to analyse zooplankton samples. Taxa were identified and counted in a Bogorov tray (constructed in the Science Faculty Workshop, University of Zululand) using a stereo dissecting microscope (Leica MZ6, Tokyo, Japan).
Macrobenthos
Samples were collected from 15 sites (Figure 1, North Lake: NL1-NL5, South Lake: SL1-SL5 and Estuary: E1-E5) during December 2004, March 2005, May 2006 and November 2006. At each site, five replicate random samples were collected by means of a Zabalocki-Type Eckman substrate grab (constructed in the Science Faculty Workshop, University of Zululand). Each grab uniformly samples 0.0236 m2 of sediment to a maximum depth of 0.05 m. After separating out the sand particles, the fauna and debris retained were preserved in 10% formalin solution and the vital dye Phloxine B (Merck Chemicals, Johannesburg, South Africa) was added to aid in sorting and counting the invertebrates in the laboratory.
Fish
Fish were sampled on three occasions (December 2004, March 2006 and November 2006) at six stations across the system (Figure 1 -NL2, NL3, SL2, SL4, E1 and E4). Three sampling methods were used, namely gill netting, and small and large seine netting. A set of gill net panels, ranging from 50 mm to 150 mm stretch bar mesh, were deployed for a minimum of 2 h at each site. The small seine net (10 m x 1.5 m) was deployed 5 m - 10 m from the shore and then pulled onto an exposed bank. The large seine net (70 m x 1.5 m) was either deployed by boat or walked out, depending on boat accessibility. All fish collected were identified, measured (Standard Length) and then returned to the system.
Data analysis
The spatial and temporal distribution of the faunal communities were analysed using cluster and non-metric multidimensional scaling (MDS) and SIMPER methods of the PRIMER V6 software9 suite of multivariate programs.10 Cluster and MDS analyses were based on the Bray-Curtis measure of similarity on fourth root-transformed data. Hierarchical clustering was based on rank orders of similarities. The analysis of similarity (ANOSIM) routine of the PRIMER software was used to determine if the biotic communities in different sections of the St Lucia system could be separated statistically on the basis of their respective assemblages. All data were input into PRIMER with the aim of testing sample relations amongst and between each other. The data were coded according to 'season', 'year' and 'lake'. To test whether observed differences were significant, either between year or area groups, data were subject to an ANOSIM. These analyses consider abundance and the number of different individuals.
Results
Salinity
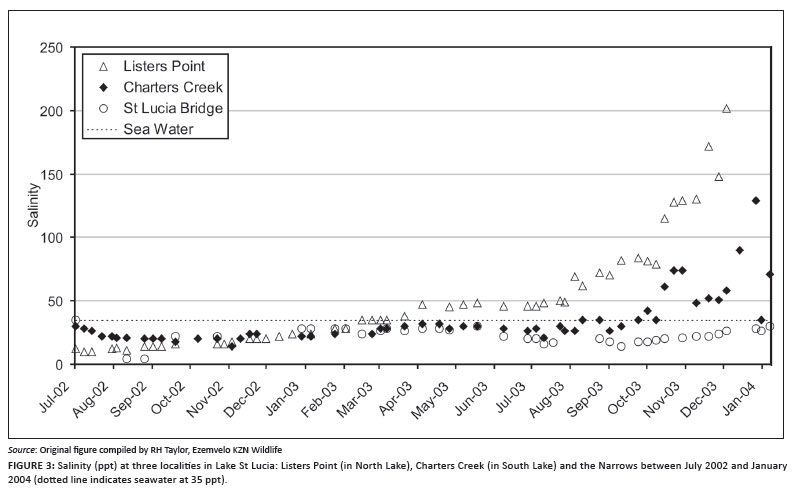
At mouth closure in June 2002, salinities throughout the system were below 35 ppt, with South Lake (Figure 3) at 30 ppt and False Bay at 10 ppt. By the beginning of March 2003 salinities in False Bay reached that of seawater. Between March and December 2003, the whole system became hypersaline; False Bay increased almost exponentially, reaching highs of 150 ppt - 200 ppt, and Charters Creek increased to between 70 ppt and 130 ppt (Figure 3). High summer evaporation, while increasing saline concentrations, also resulted in major decreases in lake levels. By December 2003 only 25% of the lake's surface area was covered with water (Figure 2).

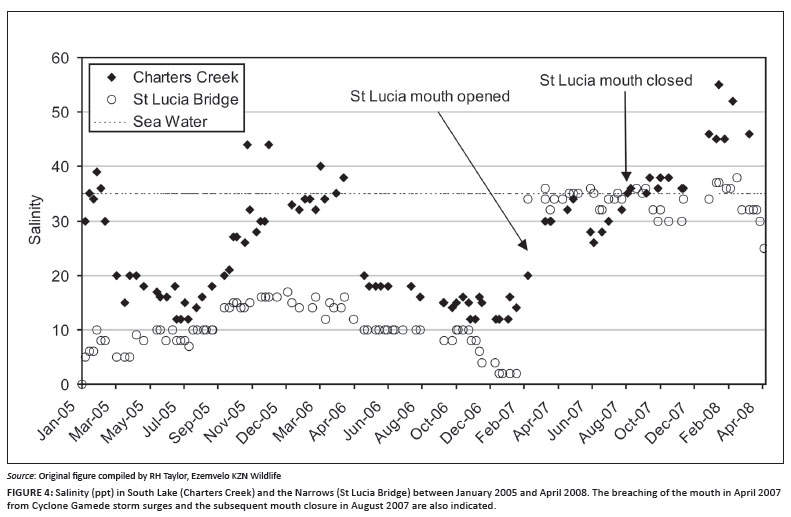
Significant rainfall at the end of January 2004 reduced salinities throughout the system to below 35 ppt. They remained slightly hyposaline until September 2004 when drought conditions again resulted in hypersaline conditions at False Bay, and by November 2004 salinities in this area had risen to >100 ppt. Late summer rains during January/February 2005 resulted in salinities being lower during the winter months but the onset of summer 2005/2006 saw salinities again starting to rise as evaporation exceeded freshwater input. By July 2006 the bulk of the system was effectively dry with only approximately 10% of the total surface area of the lake being covered with water. The onset of summer rains resulted in water coverage again reaching about 25% by December 2006. In April 2007, storm surge swells related to Cyclone Gamede moved onshore and this, coupled with high equinox tides, resulted in the breaching of the St Lucia estuary mouth. Sea water flowed into St Lucia until the mouth closed in August 2007, filling the lake to approximately 75% of capacity and stabilising salinities at 35 ppt (Figure 4).
Zooplankton
A total of 21 mesozooplankton taxa were recorded, with the estuarine calanoid copepod Pseudodiaptomus stuhlmanni dominating throughout most of the system during all three sampling sessions (Figure 5). Very high densities of mesoplankton, often exceeding 30 000/m3, were recorded in the Estuary and South Lake, even though samples were taken during daytime, which may under-record plankters such as Pseudodiaptomus because of their vertical migration.11 St Lucia is, however, a shallow and turbid system and light attenuation in the water column as a result of high turbidity could lead to an absence of vertical migration by some zooplankton species.12 A maximum of 90 000/m3 were recorded for this species at station Z5 in South Lake during May 2006.
High densities of the calanoid copepod Acartia natalensis, which is generally common in KwaZulu-Natal estuaries, were also recorded. Mesopodopsis africana was the most common mysid, especially in South Lake (Figure 5). Some meroplankton taxa were present in low numbers. The North Lake had relatively high densities of the freshwater and brackish water cyclopoid Apocyclops sp. with P. stuhlmanni also present (Figure 5). Relatively high numbers of freshwater Corixidae were recorded in the plankton at the Fanies Island station (Z2).

The MDS analysis showed two major groups at an arbitrary similarity level of 60%, with six outliers (Figure 6). The stress level in this analysis (0.05) was below 0.1, which gives a good ordination with no real prospect of a misleading interpretation.6 The outliers are all samples from stations Z1 and Z2 in the North Lake. All the South Lake samples, a few samples from the Estuary as well as from station Z3 in the North Lake, contribute to the larger of the two groups whereas the smaller group consists only of samples from the Estuary.

ANOSIM indicated a significant difference between the different sections of the system (Estuary, South Lake and North Lake, p = 0.01) but the calculated global R-value of 0.4 showed that there was nevertheless substantial overlap in the zooplankton composition of the different sections.
SIMPER analysis indicated that the factor responsible for most of the differences between the sections in the present study was density of the dominant calanoid P. stuhlmanni. Other plankters that also contributed substantially to separation of the Estuary, South and North Lakes were the calanoid A. natalensis, cyclopoid Apocyclops sp. and mysid M. africana.
Macrobenthos
Up to and in November 2006, 78 macrobenthic taxa were found at 15 sites. As these taxa included samples collected in 2004 and 2005, they could be used to compare the same sites under hypersaline and low lake level conditions. The widest range of fauna was sampled in December 2004 (58 taxa). Species richness diminished to approximately half of what was encountered initially (25 to 15 and 35 to 20 taxa between 2004 and 2006 in the North Lake and South Lake, respectively). The numbers of taxa in the Estuary remained approximately stable at between 21 (March 2005) and 25 (March 2006). In general there was a decline in the number of taxa sampled between 2004 and 2006.
A number of different groups were found, the majority of which were crustaceans (17 taxa), including Amphipoda, Brachyura, Cumacea, Isopoda, Macrura and Tanaidacea. Annelida were present in several forms (26 taxa) of which 17 taxa were classified as Polychaeta. The remaining fauna sampled included Insecta (6 taxa), Mollusca (11 taxa), and other non-annelid worms (Nematoda to Nemertea and Sipuncula).
Few taxa were ubiquitous throughout the system. Common taxa were primarily those that are truly estuarine and widespread or abundant in other KwaZulu-Natal estuaries, such as the polychaetes Ceratonereis keiskama, Dendronereis arborifera, Capitella capitata and, to a limited extent, Prionospio sexoculata. The tanaid Apseudes digitalis was also present at most sites from 2004. Cyathura estuaria was particularly dominant in December 2004 in South Lake and the Estuary (22.2% and 42.4% of the total abundance), where A. digitalis numbers were the lowest measured throughout the 5-year period. Pencil bait, Solen cylindraceus, was not found in the North Lake after December 2004, although many empty shell valves were seen along the shoreline and buried within the sediments. There were more types of molluscs early on in the study and mollusc richness decreased from the upper to lower reaches of the system. Insecta were sampled in the North Lake despite the prevalence of hypersalinity.
Overall, the highest density of macrobenthos in the system was in the estuary. The large numbers of invertebrates sampled in December 2004 (49 300 ± 44 708/m2) declined consistently and dramatically until November 2006 to 10 070 ± 2103/m2 (Figure 7C). The lowest abundances overall were recorded in North Lake and while numbers dropped to 3150 ± 3089/m2 in May 2006 (Figure 7A), they appeared to have recovered by November 2006 to approximately the densities measured in 2004. South Lake densities generally showed an improvement in macrobenthic numbers with time, ranging from 6567 ± 5303/m2 in 2004 to just under 15 000/m2 in March 2005 and November 2006.

Initially, the macrobenthos residing within the Estuary were more abundant than those in North Lake (Figure 8). In November 2006, abundances in the Lakes increased markedly to approximately triple and double in North Lake and South Lake, respectively, from the May 2006 survey. Numbers in the Estuary declined similarly so that South Lake had the highest numbers in the system at that time. Mean abundance was 9001 ± 8688/m2 in North Lake, 12 655 ± 9728/m2 in South Lake and 10 041 ± 5789/m2 in the Estuary.

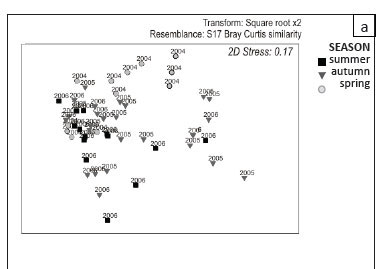
Samples collected in summer and spring were separate from each other in a two-dimensional depiction of the ordination (2D stress 0.17; Figure 9a). Autumn samples were not distinguishable from either of these seasons. Inter-annual variability was not clear from these results, although samples collected in 2004 and 2006 seemed the least similar. Coding the samples according to area within the system did show, with several exceptions, that samples collected in North Lake were different from other areas in the system. Although South Lake and the Estuary appear similar and are depicted in the three-dimensional plot as being in close proximity to each other, within this group there does appear to be some temporal separation along a top-to-bottom plot continuum of 2004, 2005 and 2006 (Figure 9b).

The global R-statistic resulting from the test for differences between sampling years showed a significant result (R = 0.575; p < 0.05). Individual pairwise tests showed significant differences between comparisons of macrobenthos from North Lake and South Lake (R = 0.682), North Lake and the Estuary (R = 0.840) and South Lake and the Estuary (R = 0.340). Differences between sample sites (North Lake, South Lake and the Estuary), were also significant (R = 0.225; p < 0.05) across the three study years. However, individual pairwise tests showed that only comparisons between 2004 and 2006 communities, and 2005 and 2006 communities were significant.
Fish fauna
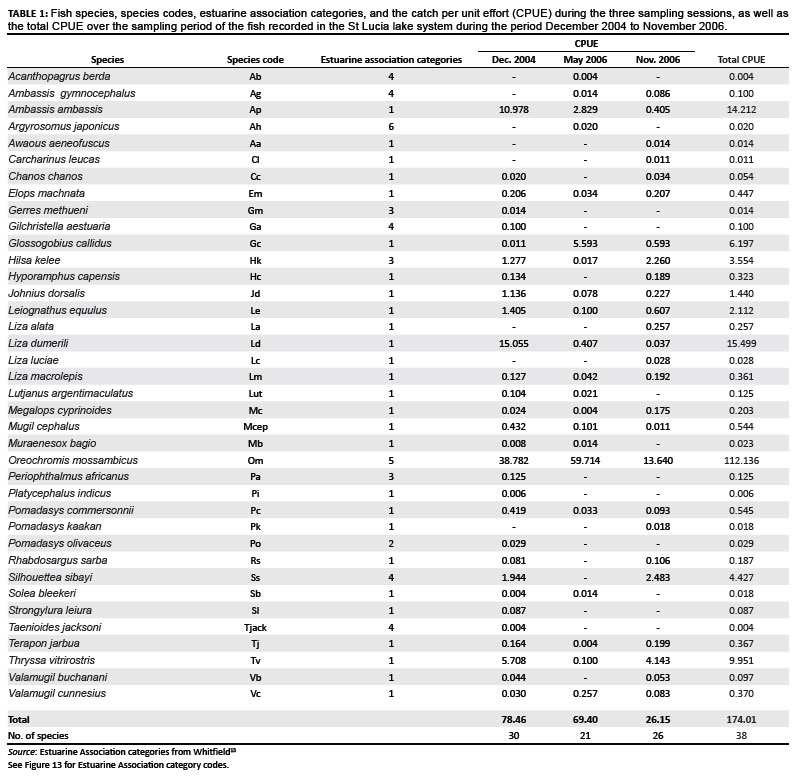
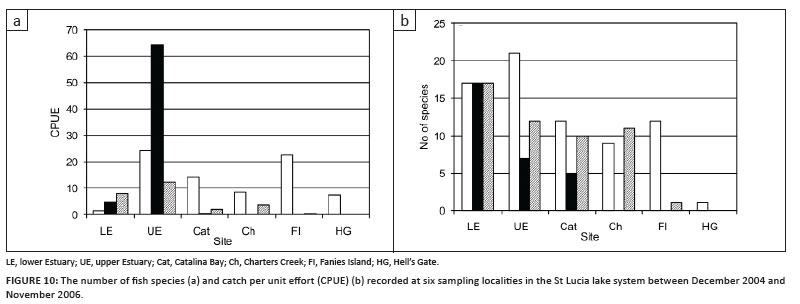
A total of 6543 fish of 38 species were recorded during the three surveys, details including the catch per unit effort (CPUE) and size ranges are given in Tables 1 and 2. There was a decrease both in the number of species recorded and the CPUE from 2004 to 2006 (Figure 10). The highest (30) and lowest (21) number of species were recorded during December 2004 and May 2006, respectively. The highest total CPUE was also recorded during December 2004, decreasing almost three-fold from December 2004 (78.46) to November 2006 (26.15).

The highest number of species was recorded in the upper and lower Estuary during all three sampling surveys (Figure 10), with the species count gradually decreasing towards the northern parts of the system. During the two 2006 sampling events, no fish were recorded at Hell's Gate (Site NL1) because of extreme low lake levels, while during May 2006, no fish were recorded at Charters Creek (SL1) and Fanies Island (NL3) (Figure 1). Similar numbers of species were recorded in the lower Estuary during all three sampling trips, whereas species counts were consistently higher during December 2004 compared to the 2006 samples at all other sites (Figure 10a). The highest CPUE (58% of total CPUE) was recorded in the upper Estuary, with the catches decreasing throughout South Lake towards Hell's Gate in the northern parts of the lake (Figure 10). The high CPUE in the upper estuary during May 2006 was dominated by a large catch (n = 570) of Oreochromis mossambicus (Figure 10b).
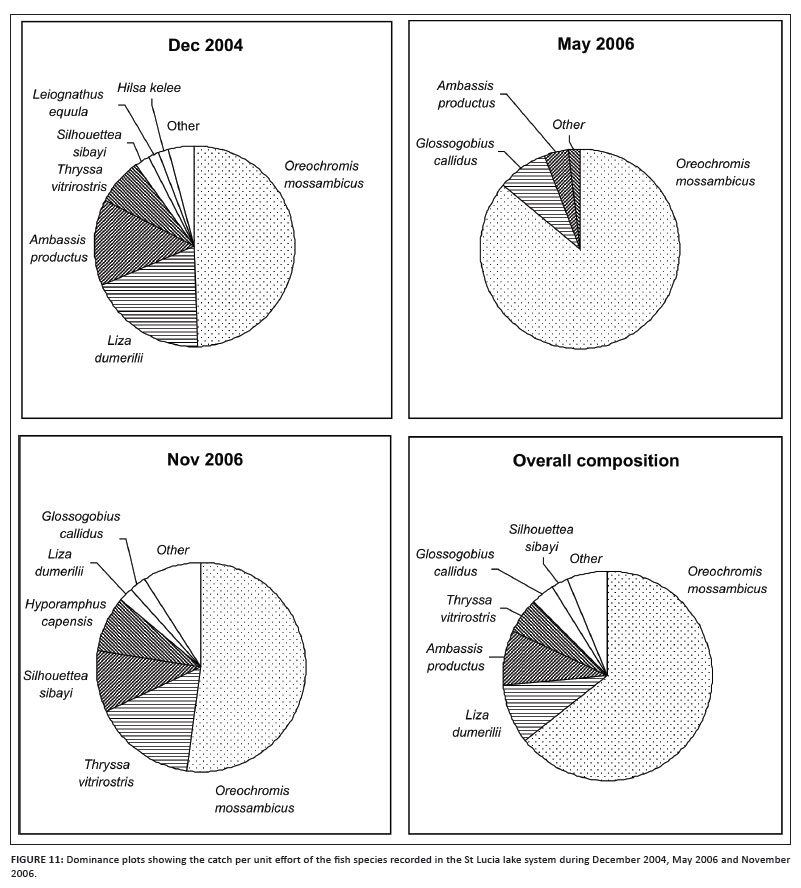
The fish catch was dominated throughout the system by the freshwater species O. mossambicus, which accounted for 46% of the total catch (Table 1; Figure 11). The other numerically dominant species included Ambassis productus (7.7%), Liza dumerili (19.7%), Johnius dorsalis (3.9%) and Thryssa vitrirostris (4.7%). O. mossambicus remained abundant during all three surveys, whereas the numbers of A. productus and L. dumerili dropped sharply after the December 2004 survey. T. vitrirostris was abundant during December 2004 and November 2006. During May 2006, O. mossambicus accounted for 86% of the CPUE.
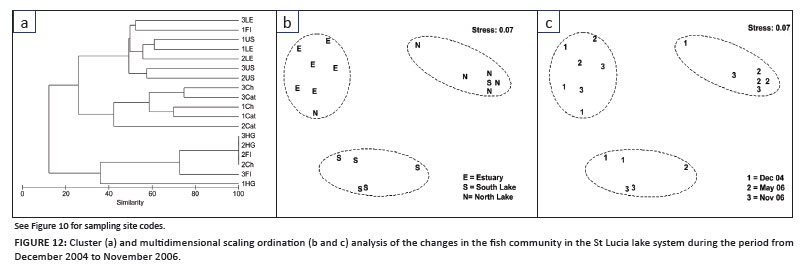
Multivariate analysis emphasised the effect of the drought and concurrent low water levels on the fish community, showing that spatial, rather than temporal, changes were the dominant factor structuring the fish community (Figure 12). The analysis revealed distinct spatial differences in the community, with three separate MDS groups formed (at 30% similarity) which represented the three compartments of the system (Figure 12a and b). There was much less inter-sample change in the fish community in each of the three compartments between sampling sessions, from December 2004 to November 2006, compared to the changes seen between the different areas (Figure 12c).
The fish community was dominated by estuarine-dependent marine immigrant species (n = 26), followed by estuarine migrants (n = 5) and estuarine residents (n = 2) (Table 1, Figure 13). Marine migrants are marine breeding species whose juveniles show varying degrees of dependence on estuaries as nurseries.14 This relatively high number of marine migrants emphasises the importance of the nursery function of the St Lucia system. However, prolonged closure of the mouth has largely prevented recruitment of post-larvae of marine species into the system during the study period resulting in a loss of the nursery function. The numerical dominance of the freshwater immigrant O. mossambicus indicated that this opportunistic species thrived in the harsh drought conditions (Table 1, Figure 13). In contrast, there was a group of 19 species, containing 12 marine migrants, such as Liza alata, Liza luciae, Acanthopagrus vagus and Argyrosomus japonicus that only occurred in low numbers. These species would be expected to be most intolerant to extended mouth closure and drought-related changes in the system.

As expected, the freshwater and resident estuarine species continued to breed during the study period, regardless of conditions. Of greater importance is the presence of juvenile marine immigrants in the system despite the extended period of mouth closure. These species include Elops machnata, Hilsa kelee, J. dorsalis, L. dumerilli and T. vitrirostris. This indicates either that juveniles of these species managed to enter the system during a short period when the Mfolozi River flooded and overtopped into St Lucia, or that these species were able to breed opportunistically in the system. However, the latter is considered unlikely.
Discussion and conclusions
Zooplankton
It is evident that the zooplankton community species richness of St Lucia has been reduced substantially during the drought period, compared to that reported by Grindley15. A few resident estuarine species thrived under present conditions, largely because normal estuarine salinities prevailed in the South Lake and Estuary areas during periods in the current study. A linkage to the sea is required in order for the stenohaline and euryhaline marine components to be able to recover.
Several different zooplankton communities were identified in the St Lucia system by Grindley15: Stenohaline and euryhaline marine components were present in the Estuary mouth area, while freshwater zooplankton was recorded where the rivers entered the lakes. Grindley15reported that most of the system was, however, dominated by an estuarine community. This latter component was dominated by the calanoids Pseudodiaptomus stuhlmanni and A. natalensis. Because of prolonged mouth closure during the present study it is not surprising that the stenohaline and euryhaline marine components of the zooplankton were not recorded. The community composition was rather typical of that of the estuarine components of other local systems such as Mhlathuze and Kosi Bay; dominated by estuarine calanoid copepods A. natalensis and P. stuhlmanni.16,17. Compared to other estuarine systems, however, the mesozooplankton diversity, in terms of species richness, was relatively low, mostly because of the lack of the marine components. This is especially evident in the copepod group; a total of 40 copepod species were recorded by Grindley15 compared to only 5 species recorded in the present study. The stenohaline component was completely absent in the present study.
Densities of the dominant species during the present study, P. stuhlmanni and A. natalensis, were however remarkably high. Although the present study consisted of only daytime sampling, densities recorded for these dominant calanoids in St Lucia, are comparable to those recorded for Pseudodiaptomus hessei and Acartia natalensis in the productive Sundays River estuary8 and much higher than those recorded in the Mhlathuze estuary and harbour, where peak densities of about 2000/m3 for both species were recorded.18 These high densities were as a result of a more stable estuarine waterbody in South Lake.
Although large parts of particularly the North Lake dried up, at times rain or seepage water was present which resulted in a poor zooplankton community. These occasional freshwater inputs prevented further hypersaline conditions during parts of the study period and allowed the presence of fresh and brackish water plankters such as the cyclopoid Apocyclops sp. This is, as far as could be determined, a first record for the Apocyclops genus in South Africa.
Macrobenthos
Overall, the macrobenthos within St Lucia was variable in terms of interannual differences and according to where it was sampled in the system. This seems to be typical of recent studies on these communities.19 A comparison of the abundance of macrobenthos between different studies is difficult as, apart from the most recent published survey conducted under the current drought,19 no other studies have surveyed St Lucia as a whole system. Rather, the macrobenthos has been studied in smaller discreet sampling areas and never over a prolonged, comparable period.
Macrobenthic diversity in the St Lucia system has been reported as low,6 perhaps attributable to the size of the system and its inherent variability. Nonetheless, up to 84 taxa were sampled in the first comprehensive study of the macrobenthos of St Lucia.4,20,21 More recently, only 23 taxa were reported for the survey in 2005 under the current drought.19 This study sampled 78 taxa under the extreme conditions of the drought that presented physiological challenges of hypersalinity and even desiccation of infauna when lake levels receded.
Under similar hypersaline conditions, 9 species in North Lake and 21 in South Lake were recorded.21 Although various areas were depauperate at times (e.g. North Lake in autumn 2006), faunal abundance was relatively high considering the extreme environmental conditions to which these animals were subject.
Macrobenthic assemblages in the St Lucia system have previously been dominated by amphipods, brachyurans and gastropods.6 Amphipods and polychaetes were the most numerically abundant taxa in this study. The macrobenthic fauna of the St Lucia system were characterised by typically estuarine and euryhaline marine taxa. The tanaid A. digitalis was present at most sites from 2004 and is a filter-feeding, detritivore that constructs tubes in sandy substrata.22 It is speculated that, because of their distribution differences, a competitive relationship exists between A. digitalis and the anthurid isopod, C. estuaria, possibly for space, habitat and/or diet. C. estuaria was particularly dominant in December 2004 in South Lake and the Estuary, where A. digitalis numbers were the lowest measured throughout the study period.
The resilience of macrobenthic organisms to environmental change is manifested in their ability to withstand or adapt to changes in abiotic components. The majority of macrobenthos currently in St Lucia are presumably opportunistic in nature, owing to the fairly rapid recolonisation of low-density regions following periods of high salinity and low lake levels. These opportunists or pioneering species are generally characterised by high reproductive rates, good dispersal potential and inferior competitive ability following the return to stable conditions.23 Several taxa found may be regarded as opportunists or r-selected species. The polychaete C. capitata and, to a limited extent, P. sexoculata are tolerant organisms that are resistant to large habitat and physicochemical conditions and are able to tolerate even adverse conditions where few other taxa are able to exist.24 A considerable decline with time in r-selected species did not occur during the present study, as has been previously found.23 This may be attributed to the continued drought-induced pressures, and hence inhibition of K-selected species establishment within the macrobenthos.
Despite the severe and prolonged environmental conditions within St Lucia, several important estuarine species were able to exist and still dominate without being overcome in numbers by pioneering tolerant species such as C. capitata. A. digitalis and Grandidierella bonnieroides are notable in this regard. The deposit-feeding tubicolous amphipod G. bonnieroides inhabits muddy substrata. It is classified as a tropical euryhaline, eurythermal marine species and has a salinity tolerance range of 10 ppt - 50 ppt.6,25
Fish fauna
Hypersaline conditions recorded during periods with salinities above 70 ppt caused numbers of many marine migrant fish species to decline markedly. These results coincide with reports by Whitfield et al.26, who found that less than 40% of the marine migrants were recorded in salinities above 65 ppt in the St Lucia system, while very few species were able to survive in salinities above 70 ppt. Cyrus and Vivier27 found only one species, O. mossambicus, surviving at Site NL1 in the North Lake (Figure 1) when salinities exceeded 110 ppt. The net result was that the nursery function for juveniles of marine origin was largely lost, while no recruitment of marine post-larvae has taken place during the study period. This is further exacerbated by the extensive period of mouth closure affecting the recruitment of younger cohorts, of species such as Rhabdosargus sarba, whose adult stocks in the marine environment are being impacted on as a result of the lack of maturing individuals leaving St Lucia and joining the offshore spawning stocks.28
The breaching of the mouth in March 2007 would have resulted in some recruitment of post-larval fish and invertebrates that had been spawned offshore and were trying to make their way into the estuary. The extent of the recruitment would have been limited as the system only remained open between March to August 2007 and did not coincide with the peak spring and summer recruitment peaks.14 The breaching did however allow marine water to partially fill the system and re-establish the role as a nursery for juveniles of those species that managed to recruit into the system during this period. However, the mouth has remained closed from August 2007 to October 2010 and salinities have once again started to increase. Should this closure continue, then failure of the nursery function is likely, resulting in further declines in offshore breeding stocks of the estuarine-associated species.
Function as a nursery for marine fish
This study has shown that the three major faunal groups in St Lucia system have been severely impacted by the extended periods of mouth closure, low lake levels and hypersalinity events to the extent that it is now considered to be in crisis. This is of concern, as the St Lucia estuary is one of the most important estuaries in South Africa.17,29
The lack of a connection to the sea during the study, coupled with major changes and fluctuations in the physical conditions within the system, have also had an impact on the offshore marine ecosystem where a lack of recruitment over a 5-year period could affect the breeding stock of marine species whose life cycles are associated with estuaries. This has already been shown for the marine spawning fish R. sarba.28 The offshore fish and prawn species that use St Lucia as a nursery habitat will be most affected by these events. Changes in the zooplankton and macrobenthic invertebrates also directly impact on the fish fauna as many of the species present feed on these groups.
Implications for future management of the system
The St Lucia system historically shared a common mouth with the Mfolozi estuary, known as St Lucia Bay. It has been concluded that historically, when drought conditions affected these systems and a sand bar formed across the bay, water from the Mfolozi, which has the second largest catchment in KwaZulu-Natal, would have flowed up the Narrows (Figure 1) and into the lake.4 This would have contributed to more frequent mouth breaching, to keeping lake levels high and also with stabilising salinities within the system.
Since the closure of the mouth in August 2007, the potential implications for the biota of closing the Mfolozi Estuary and diverting it into Lake St Lucia have been assessed.30 The negative impacts were considered relatively insignificant in relation to what benefit there would be to the receiving system. The two systems were therefore joined over the winter of 2008. Despite some water reaching St Lucia, winter flows that year were extremely low and did not yield the envisaged result. Under the current, long-term drought conditions there is a need for a more permanent connection between St Lucia and Mfolozi, to bring it closer to the historical condition.2
A current initiative involves an application by the iSimangaliso Wetland Park Authority to investigate the potential for long-term reconnection of the St Lucia and Mfolozi estuaries and the reclamation of low-lying swamplands that have been lost to agriculture. Such an initiative could alleviate some of the ecological stresses affecting Lake St Lucia. Also, this action will make a significant contribution to maintaining some ecosystem function through major drought/low-level periods. However, the project is still in its infancy and it is unlikely that any major changes will be made for several years.
Freshwater inflow contributes up to 50% of the water to the system under normal years, and the other 50% comes from direct input from rainfall (Taylor RH 2007, personal communication, February 21). Inflow from rivers has been compromised, and rainfall has recently been erratic, although data on these inflows are not available. This means that for the 5 years prior to the mouth breach in 2002, fauna within the system have had to tolerate and survive conditions of abnormally high salinity. Those post-larval fish and prawns which recruited into the system when it opened in 2007 now face similar conditions.
If the system is subject to a similar set of conditions that it endured during this study from 2002 to 2006, the fauna recruited in 2007 will not survive. Similarly, the impacts on the offshore marine environment, which is potentially of greater significance, could have significant negative effects for the entire KwaZulu-Natal marine-estuarine ecosystem. Manual breaching of the St Lucia mouth may need to be considered once the marine-associated fauna inside the Lakes reach an age of first maturity. A connection to the sea at such a time should result in recruitment into the offshore breeding stocks of many marine species whose juveniles are dependent on St Lucia as their nursery ground. However, the importance of reconnecting the Mfolozi to St Lucia cannot be understated because of its historical role of sustaining the lake during the closed-mouth condition.2,3 Until such time, St Lucia and its fauna will have to rely on limited and erratic freshwater sources, be it via river flow, direct rainfall or temporary connections to the Mfolozi.
Acknowledgements
We thank the staff from the Department of Zoology, University of Zululand and Oceanographic Research Institute for assistance with fieldwork, Ricky Taylor (Ezemvelo KZN Wildlife) for provision of data for Figures 2, 3 and 4, Caroline Fox (EKZNW) for assistance in the field, and iSimangaliso Wetlands Park Authority for access to operate on the St Lucia system. This article forms part of the St Lucia Estuarine System Research Programme funded as a joint venture through Marine and Coastal Management, Department of Environmental Affairs and Tourism, and the National Research Foundation, South Africa, with additional funding from the Research Committee, University of Zululand and the Oceanographic Research Institute.
References
1. Begg G. The estuaries of Natal. Natal Town and Regional Planning Report 41. 1978; p. 657. [ Links ]
2. Whitfield AK, Taylor R. A review of the importance of freshwater inflow to the future conservation of Lake St Lucia. Aquatic Conservation: Marine and Freshwater Ecosystems. 2009;19:838-848. [ Links ]
3. Taylor RH. Patterns of physical variability in the St Lucia Estuary. Paper 1. In: Ecological responses to changes in the physical environment of the St Lucia estuary. Norwegian University of Life Sciences. Doctor scientarium thesis. 2006;7:1-41. [ Links ]
4. Day JH, Millard NAH, Broekhuysen GJ. The ecology of South African estuaries. Part V: The St Lucia system. Trans R Soc S Afr. 1954;34;129-156. [ Links ]
5. Blaber SJM. Fish communities of south-east African coastal lakes. In: Leveque C, Bruton MN, Ssentongo GW, editors. Biology and ecology of African freshwater fishes. Paris: Orstom, 1988; p. 351-362. [ Links ]
6. Day J. Summaries of current knowledge of 43 estuaries in southern Africa. In: Day JH, editor. Estuarine ecology with particular reference to southern Africa. Cape Town: Belkama, 1981; p. 251-330. [ Links ]
7. Taylor RH. Biological responses to changing salinity. In: Taylor RH, editor. Proceedings of the workshop on water requirements for Lake St Lucia. Pretoria: Department of Environment Affairs, 1993; p. 1-83. [ Links ]
8. Jerling HL, Wooldridge TH. Plankton distribution and abundance in the Sundays River estuary, South Africa, with comments on potential feeding interactions. S Afr J Mar Sci. 1995;15:169-184. [ Links ]
9. PRIMER. Version 6. Ivybridge: Primer Enterprisers. [ Links ]
10. Clarke KR, Warwick RM. Change in marine communities: An approach to statistical analysis and interpretation. Plymouth: Plymouth Marine Laboratory, 1994; p. 144. [ Links ]
11. Grindley JR. The vertical migration behaviour of estuarine plankton. Zool Afr. 1972;7:13-20. [ Links ]
12. Champalbert G, Pagano M, Sene P, Corbin D. Relationships between meso- and macro-zooplankton communities and hydrology in the Senegal River Estuary. Estuar Coast Shelf Sci. 2007;74:381-394. [ Links ]
13. Whitfield AK. An estuary-association classification for the fishes of southern Africa. S Afr J Sci. 1994;90:411-417. [ Links ]
14. Whitfield AK. Biology and ecology of fishes in southern African estuaries. Ichthyological. Monographs of the J.L.B. Smith Institute of Ichthyology. 1998;(2):1-223. [ Links ]
15. Grindley JR. The role of zooplankton in the St Lucia system. In: Taylor RH, editor. St Lucia research review. Pietermaritzburg: Ezemvelo KZN Wildlife, 1982; p. 88-107. [ Links ]
16. Jerling HL. The zooplankton community of the Mhlathuze (Richards Bay) estuary: Two decades after construction of the harbour. Afr J Mar Sci. 2003;25:289-299. [ Links ]
17. Jerling HL. The zooplankton community of Richards Bay Harbour and adjacent Mhlathuze Estuary, South Africa. Afr J Mar Sci. 2008;30:55-62. [ Links ]
18. Jerling HL, Weerts S. Mesozooplankton community structure of the Kosi Bay Lake system. Paper presented at: Joint SASAQS/PSSA Conference; 2006 Jun 19-22; Maputo, Mozambique. [ Links ]
19. Pillay D, Perissinotto R. The benthic macrofauna of the St Lucia Estuary during the 2005 drought year. Estuar Coast Shelf Sci. 2008;77:35-46. [ Links ]
20. Millard NAH, Broekhuysen GJ. The ecology of South African estuaries. Part X: St Lucia: A second report. Zool Afr. 1970;5:277-307. [ Links ]
21. Boltt RE. The benthos of some southern African lakes. Part V: The recovery of the benthic fauna of St Lucia Lake following a period of excessively high salinity. Trans R Soc S Afr. 1975;41:295-323. [ Links ]
22. Reavell PE, Cyrus DP. Preliminary observations on the macrocrustacea of coastal lakes in the vicinity of Richards Bay, Zululand, South Africa. S Afr J Aquat Sci. 1989;15:103-128. [ Links ]
23. Santos SL, Simon JL. Responses of soft-bottom benthos to annual catastrophic disturbance in a South Florida estuary. Mar Ecol: Prog Ser. 1980;3:347-355. [ Links ]
24. Grassle JF, Grassle JP. Opportunistic life histories and genetic systems in marine benthic polychaetes. Mar Res. 1974;32:253-284. [ Links ]
25. Nair KKC, Gopalakrishnan TC, Venugopal P, Peter MG, Jayalakshmi KV, Rao TSS. Population dynamics of estuarine amphipods in Cochin backwaters. Mar Ecol: Prog Ser. 1983;10:289-295. [ Links ]
26. Whitfield AK, Taylor RH, Fox C, Cyrus DP. Fishes and salinities in the St Lucia system -a review. Rev Fish Biol Fish. 2006;16:1-20. [ Links ]
27. Cyrus DP, Vivier L. Status of the estuarine fish fauna in the St Lucia estuarine system, South Africa, after 30 months of mouth closure. Afr J Aquat Sci. 2006;31(1):71-81. [ Links ]
28. Mann BQ, Pradervand P. Declining catch per unit effort of an estuarine-dependent fish, Rhabdosargus sarba (Teleostei: Sparidae), in the marine environment following closure of the St Lucia Estuarine System, South Africa. Afr J Aquat Sci. 2007;32:133-138. [ Links ]
29. Turpie JK, Adams JB, Joubert J, et al. Assessment of the conservation priority status of South African estuaries for use in management and water allocation. Water SA. 2002;28(2):191-206. [ Links ]
30. Cyrus DP, Jerling HL, Owen RK, Vivier L. Ecological impacts of a short-term diversion of the uMfolozi/uMsunduzi Estuary into Lake St Lucia. CRUZ Investigational Report. 2008;123:1-14. [ Links ]
 Correspondence to:
Correspondence to:
Digby Cyrus
Postal address:
Coastal Research Unit of Zululand, Department of Zoology, University of Zululand
Private Bag X1001
KwaDlangezwa 3886 KwaZulu-Natal, South Africa
email: dcyrus@pan.uzulu.ac.za
Received: 01 July 2009
Accepted: 22 Nov. 2010
Published: 09 Mar. 2011
© 2011. The Authors. Licensee: OpenJournals Publishing. This work is licensed under the Creative Commons Attribution License.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}