










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.107 no.1-2 Pretoria ene./feb. 2011
http://dx.doi.org/10.4102/sajs.v107i1/2.230
RESEARCH LETTERS
Pathologies in the Early Pliocene phocid seals from Langebaanweg, South Africa
Romala GovenderI; Graham AveryII,III; Anusuya ChinsamyI
IDepartment of Zoology, University of Cape Town, South Africa
IICenozoic Studies, Iziko South African Museum, Cape Town, South Africa
IIIDepartment of Archaeology, University of Cape Town, South Africa
ABSTRACT
Abundant vertebrate fossils have been recorded from the Early Pliocene locality, Langebaanweg, South Africa. This study documents the pathologies evident in the 5 million-year-old fossil phocid seal assemblage. Careful anatomical assessment of the remains revealed that 0.73% showed evidence of disease and/or trauma. The majority of the ailments were forms of osteoarthritis, although periodontitis and osteomyelitis were also evident. Some bones also showed healed fractures, suggesting that the individuals survived the traumatic event. Two cases of dental pathologies were also noted.
Introduction
Langebaanweg, approximately 110 km north-east of Cape Town in South Africa is a prolific source of well-preserved Late Tertiary fossils (24 MYA - 1.7 MYA), which were exposed during phosphate mining.1,2 'E' Quarry was considered a prime locality for palaeoecological studies because it has yielded a rich Early Pliocene (5 MYA) faunal assemblage and palaeopalynological information during a period of change from moderate to more cooler climates.1,3 Studies of the fossils have provided a detailed understanding of the terrestrial fauna and flora along the west coast of South Africa during the Late Tertiary.2,3 The Early Pliocene represents a time when climatic change included a decrease in global temperatures and movement of the African continent deflected the polar currents resulting in the development of the nutrient-rich Benguela upwelling system off the South African coast.1,4 Hendey1 considered that the Langebaanweg area was experiencing dry summers and wet winters at the time represented by the Muishond Fontein Phosphatic Sand Member.5
The Early Pliocene fossils were recovered from the Muishond Fontein Phosphatic Sand Member of 'E' Quarry, which is considered to represent a shallow marine environment between an offshore bar and beach and includes fluvial and bankside deposits.1,4,5 The sediments were most likely laid down during a single transgression phase during the Early Pliocene.1 Fossil seal remains occur in Beds 3aS and 3aN1,4 (Muishond Fontein Phosphatic Sand Member5). It represents a river channel deposit most likely deposited during flooding.1 The seals have been identified as Homiphoca capensis (Family: Phocidae)6,7 and, based on the presence of neonates as well as adults, it was concluded that a resident population existed at the time.1
Renewed interest in the marine component at Langebaanweg has led to a reassessment of the morphology and palaeoecology of H. capensis. Previous analysis of the well-preserved seal remains has documented that some of the seals had pathologies but these were not studied further. Hendey2 noted that the Langebaanweg carnivores exhibited a higher prevalence of pathologies than the herbivores and presumed this to be as a result of their longer lifespans. Many of the herbivores from Langebaanweg (e.g. sivatheres) show enamel hypoplasia and tooth wear, which indicate dietary and environmental stresses.2,8 During the current study we were able to analyse the pathologies on the seal bones and make some inferences as to the causes and effects of the pathologies.
Materials and methods
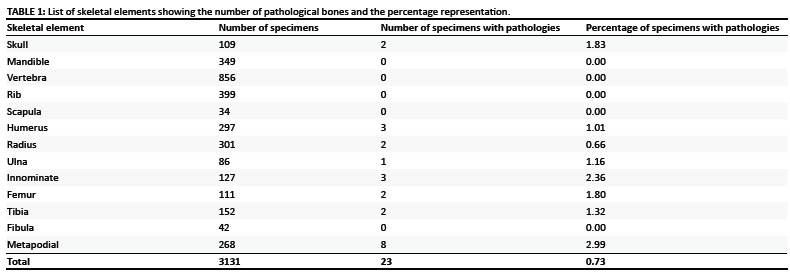
The phocid seal assemblage from 'E' Quarry comprises a total of 3131 complete, incomplete and fragmentary cranial and postcranial elements (Table 1). Most of the pathological specimens are representative of adults. The bones that show pathological changes were isolated i.e. not associated with other postcranial or cranial material. Pathologies were documented in 21 postcranial and 2 cranial specimens. This represents 0.73% of the total number of seal remains with the majority of pathologies (91.3%) occurring on postcranial elements. The two cranial pathologies observed were on premaxillae while the postcranial pathologies occurred on both complete and incomplete limb and flipper bones. Definitions of pathologies follow Defrance9 and Waldron10.
Description of skeletal pathologies
Osteoarthritis
Distal epiphyses of two humerii have distinct pathological changes, whereas a third, complete humerus has changes at various points along its length. In SAM-PQ-L56007 (Figure 1a) the distal articulation has been obliterated by exostoses (additional bone growth9) and the surface is pitted (changes to the contour of the bone9) with excavations. In SAM-PQ-L60194 (Figure 1b), only the medial condyle is affected; there are minor exostoses, the surface is porous and periosteal lesions are present. Marginal osteophytes (formation of new bone around the margins of the joint10) and exostoses are evident proximally on SAM-PQ-L40860. The humeral head shows lipping and has a porous surface. In addition, damage on the head laterally reveals subchondral trabeculae.
The posterior surface of the distal femur (SAM-PQ-L62914) is covered by exostoses (Figure 1c) and the articulating surface is eroded. Normal development appears to have been affected in the femur of SAM-PQ-L50935 (Figure 1d), which resulted in it being shorter and more robust than any of the other femora preserved. The lateral border is uneven and pitted in appearance as a result of extensive exostoses. The articulating surface of the lateral condyle has been damaged, showing a split that has filled with new bone growth and extensive bone growth along its anterior border.
Changes to the proximal joint and surrounding surfaces of a tibia (SAM-PQ-L50300) are evident. Exostoses extend proximally and medially, forming a bony plate in front of the lateral articulation; medially there is minor growth along the articulating surface edge. Erosion of the articulating surface has resulted in the outline of the articulating condyles being barely visible (Figure 1e). The articulating surface is covered by bone proliferation, which was most likely as a result of increased vascularisation10 as well as pitting resulting in perforations. It is well documented in human studies that pitting results in holes which allow communication with subchondral cysts.10 A second proximal tibia (SAM-PQ-L45841) has lipping anteriorly on the articulating surface and porosity on the articulating surface, suggestive of the early stages of osteoarthritis.
Proximal and distal articulations of metapodials show new bone growth as well as pitting and erosion. An incomplete phalanx (SAM-PQ-L42205) shows extensive bone growth on its ventral surface. A fragment (SAM-PQ-L31064) shows disruption of the articulating surface and porosity of the surface. On the ventral surface of the metapodial fragment (SAM-PQ-L64075) there is evidence of periosteal lesions.
The articulating surfaces of the metapodials are eroded, showing visible trabeculae and sharp, scalloped edges, which suggest that this damage may have affected the joint structures. SAM-PQ-L42205 and SAM-PQ-L48033 show extensive exostoses on the ventral surface similar to that described in camelids that worked on hard surfaces, most likely as pack animals.9
Our analysis of the postcranial elements shows that they deviate from the normal bones in the collection, showing the presence of marginal osteophytes, proliferation of bone and pitting on the joint surface showing subchondral bone that indicates that these seals most likely suffered from osteoarthritis.10,11,12 The lack of eburnation of the joint surfaces is similar to that in South American camelids.9 The proliferation of bone on the distal articulating surfaces of SAM-PQ-L56007, SAM-PQ-L60194 and SAM-PQ-L50300 suggests an extreme inflammatory response in the joint which implies that the arthritis was severe. On the lateral side of a humeral head (SAM-PQ-L40860) there is an oval area that shows subchondral bone attached to the head suggesting that it may have been caused by a subchondral cyst. After observing other humeri in the collection, the deeper muscle attachment scarring on the lateral surface of SAM-PQ-L56007 suggests that muscles of the elbow were overly developed and had a stronger action.
Trauma
SAM-PQ-L33262 consists of an associated ulna and radius. The ulna has a healed fracture with displacement of the proximal half of the bone (Figure 1f). Subchondral bone is visible on the proximal articulating surface of the ulna. Exposed trabeculae, scalloped edges and a scooped floor on the articulation surface is similar to the ulna figured and described by Waldron10, which would indicate the possibility of rheumatoid arthritis. The edges of this area are smooth, which might indicate healing. In chimpanzees, traumatic injuries are often accompanied by degenerative joint diseases.13 The associated radius (SAM-PQ-L33262) shows bone proliferation, which resulted in a lip forming along the distal edge of the articulation and the loss of periosteal bone posteriorly on the head, indicating the early stages of arthritis as demonstrated in the human remains described by White and Folkens12 that had developed traumatic arthritis.
Most of the damage on the metapodials is seen on the articulating surface, except for SAM-PQ-L48033 (Figure 2a) which had been fractured about a third of the way from the distal articulation. The presence of bone growth on the dorsal surface indicates that, although the fracture healed, the distal end of the bone is slightly bent as a result of misalignment caused by trauma.
Disease
Some examples suggest disease as the causal factor for the skeletal pathologies. Subchondral bone has been exposed in the right acetabulum of SAM-PQ-L30080 (Figure 2b). Surrounding the acetabulum and extending onto the ilium and the ischium there is extensive periosteal bone reaction with resulting exostoses similar to that seen in SAM-PQ-L45976 (Figure 2c), but the latter is more porous.
In SAM-PQ-L45976, the rim of the acetabulum is much higher, which indicates that there may have been a change in the position of the femoral head. As a result of the change in the position of the femur secondary osteoarthritis developed. The periosteal reaction also points to possible infection. Part of the ischium, pubis and the posterior part of the acetabulum are preserved on the left innominate (SAM-PQ-L30080). There is evidence of a periosteal bone reaction on the ischium in particular, although minor growth is evident on the pubis. In both innominate specimens there is evidence of bone growth, possible drainage channels and an increase in the size of the bone which would suggest osteomyelitis (an infection affecting the bone and bone marrow).10 The radius (SAM-PQ-L31111) shows a reaction on the bone surface (periosteum) represented by minor bone growth along the distal expansion's medial surface and posterior margin, similar to that described in Iron Age horse vertebrae, ribs and sacra, and possibly caused by bacterial infection.14
Dental pathologies
Dental pathologies are evident on two premaxilla fragments. SAM-PQ-L70460 (Figure 2d) has all four sockets while only the two left sockets are present in SAM-PQ-L70483 (Figure 2e). A small supernumerary tooth socket is present in the midline of SAM-PQ-L70460 and its general shape suggests that it was well developed. Supernumerary teeth in extant northern elephant seals were relatively common on mandibles.15 None of the seal mandibles exhibited supernumerary teeth or sockets.
Both tooth sockets of SAM-PQ-L70483 appear to be enlarged. The pathologically excavated socket walls are concave and the periosteum is damaged showing the subchondral bone and the presence of vascular foramina10,15 suggesting periodontitis (an inflammatory reaction in the periodontium which can spread to the surrounding alveolar bone causing its destruction10). Reabsorption is evident in the median wall separating the medial sockets and the smooth edges suggest that healing had occurred (Figure 2f). The bone loss in the wall separating the medial incisors was possibly because of an abscess,11 although it may be further evidence of an infection that caused periodontitis.10,12
Discusssion
Although E' Quarry has yielded a substantial sample of seal bones it is likely that mining removed many fossils prior to the collecting initiated by the museum. Our analysis of the fossil seals from Langebaanweg has demonstrated a relatively low incidence of pathologies (0.73%) with the majority (91.3%) represented on the postcranial elements. Only a small proportion of specimens collected were afflicted by disease and/or trauma. Currently, minimal comparative data is available with regard to the incidence of pathologies in modern seal populations.
Osteoarthritis has been noted mainly on single joints of limbs at the elbow and knee as well as on foot bones. The tibia (SAM-PQ-L50300), femur (SAM-PQ-L62914) and humerus (SAM-PQ-L56007), in particular, show fairly severe reactions, which would have affected the functioning of the joints. The epiphyses of these particular elements are fused, indicating the adult status of the individuals. The bone growth on the tibia surrounding the joint surface to the lateral side could have been a way of accommodating damage to the joint, as seen in a bull mammoth where there was possibly subluxation (partial dislocation) of the knee.16 Phocid seals hunted and interacted socially in water, so these arthritic joints may not have hampered their movements. A number of the foot bones had developed osteoarthritis which would have affected the animal's movement as it matured. Eburnation was not noted in any of the arthritic joints of the Langebaanweg seals possibly because the individuals died before the disease had progressed to the advanced stage of bone on bone contact.
The fractured ulna and its associated radius show that the muscles of the elbow joint possibly went into spasm with the result that the ulna became shortened, in a manner similar to that recorded for a young pig from a Roman settlement.17 Again, because the fracture had healed, the animal not only survived but also lived for some time after the trauma, although the shorter limb may have hampered movement on land and perhaps to a lesser degree, in the water as well. Seals need to be agile and manoeuvrable in order to hunt and to avoid other seals and especially to evade predators. Such damage to the forelimb may well have resulted in this particular individual being unable to venture far from the shore where food was more readily accessible. The fractured phalange had healed with slight deformation, but probably did not hamper movement in spite of secondary osteoarthritis that had developed. The fractured metapodial shows signs of remodelling as a result of exostoses.11,18
There is evidence of possible osteomyelitis on the pelvic regions of two specimens. In SAM-PQ-L30080 the infection has extended bilaterally but the left side is incomplete and appears not to have been as severe. Phocid seals interact socially at sea. These interactions include mating displays as well as intra-specific male aggression for mates. Male on male interaction increases towards the end of the mating season when there are more females in oestrous and this is when neck and hind flipper injuries increase.19 It is thus possible that the osteomyelitis of SAM-PQ-L30080 was caused by a bite. Unfortunately there is no evidence of this on the bone, but damage to surrounding muscle would have been sufficient to provide an entry point for infection. Associated vertebral epiphyses are unfused, indicating its immature ontogenetic status. It is possible that it may have been attacked by an adult (intra-specific aggression), although this injury does not rule out a predator attack. The second innominate (SAM-PQ-L45976) also has bone growth on the lateral surface of the ilium as well as indications that the femoral head had shifted its position. This infection may have been as a result of a subluxation of the femur which caused damage to the joint, bleeding and inflammation in the joint capsule, or an infection as a result of trauma to the surrounding tissue that resulted in secondary osteoarthritis.
The dental pathologies represent only 8.7% of the total pathologies and both incidences involved teeth of the maxilla. Currently there are no recognisable pathological changes to 349 mandibular remains from Langebaanweg. Damage to the maxillary region could have resulted from accidents during feeding, playing or fighting, which could have caused damage to the periodontium that may have become infected and later spread to the surrounding bone causing its destruction. The maxillae are fragments and it is difficult to comment further on the exact nature of the injury.
Conclusion
The analysis of Pliocene phocid seals from Langebaanweg showed that a relatively small proportion of bones showed signs of pathology (0.73%) and that 91.3% of the pathologies were observed on postcranial elements. The bones predominantly afflicted were those of the forelimb, pelvic girdle, hindlimb and foot, as well as the skull. Fusion of epiphyses indicates that most of the afflicted were adults. The majority of the individuals suffered from varying degrees of osteoarthritis, but osteomyelitis and periodontitis also were evident; one individual had a supernumerary incisor. Healing of a fractured ulna indicates that the individual survived, although muscle action had caused displacement, which was evident on the bone. Our analysis of the pathologies evident in the fossil seals suggests that ontogenetic maturity, as well as intra-population interactions, led to pathological changes in the skeletons.
Acknowledgements
We thank the National Research Foundation of South Africa for funding (grant number 68889 awarded to AC), Deano Stynder and Kerwin Van Willingh for providing access to the Langebaanweg collections at Iziko South African Museum in Cape Town and Valerie Mienies for counting of the remains of the seals from the Langebaanweg collection.
References
1. Hendey QB. Palaeoecology of the Late Tertiary fossil occurrences in 'E' Quarry, Langebaanweg, South Africa, and a reinterpretation of their geological context. Ann S Afr Mus. 1981;84(1):1-104. [ Links ]
2. Hendey QB. Langebaanweg: A record of past life. Cape Town: South African Museum; 1989. [ Links ]
3. Hendey QB. The Pliocene fossil occurrences in 'E' Quarry, Langebaanweg, South Africa. Ann S Afr Mus. 1976;69(9):215-247. [ Links ]
4. Pether J, Roberts DL, Ward JD. Deposits of the West Coast. In: Partridge TC, Maud RR, editors. The Cenozoic of southern Africa. Oxford: Oxford University Press, 2000; p. 33-54. [ Links ]
5. Roberts DL. Varswater Formation (including the Langeenheid Clayey Sand, KoningVlei Gravel, Langeberg Quartz Sand and Muishond Fontein Phosphatic Sand member). In: Johnson MR, editor. Catalogue of South African lithostratigraphic units. Pretoria: South African Committee on Stratigraphy, 2006; p. 27-31. [ Links ]
6. Hendey QB, Repenning CA. A Pliocene phocid seal from South Africa. Ann S Afr Mus. 1972;59(4):71-98. [ Links ]
7. Muizon de C, Hendey QB. Late Tertiary seals from the southern Atlantic ocean. Ann S Afr Mus. 1980;82(3):91-128. [ Links ]
8. Franz-Odendaal T, Lee-Thorp J, Chinsamy A. Insights from stable light isotopes on enamel defects and weaning in Pliocene herbivores. J Biosci. 2003;28:765-773. [ Links ]
9. Defrance SD. Paleopathology and health of native and introduced animals on southern Peruvian and Bolivian Spanish colonial sites. Int J Osteoarchaeol. 2010;20:508-524. [ Links ]
10. Waldron T. Palaeopathology. New York: Cambridge University Press; 2009. [ Links ]
11. Rothschild BM, Martin LD. Paleopathology: Disease in the fossil record. Boca Raton: CRC Press Inc.; 1993. [ Links ]
12. White TD, Folkens PA. Human osteology. 2nd ed. San Deigo: Academic Press; 2000. [ Links ]
13. Carter ML, Pontzer H, Wrangham RW, Peterhans JK. Skeletal pathology in Pan troglodytes schweinfurthii in Kibale National Park, Uganda. Am J Phys Anthropol. 2008;135:389-403. [ Links ]
14. Bendrey R, Taylor GM, Bouwman AS, Cassidy JP. Suspected bacterial disease in two archaeological horse skeletons from southern England: Palaeopathological and biomolecular studies. J Archaeol Sci. 2008;35:1581-1590. [ Links ]
15. Abbott C, Verstraete FJM. The dental pathology of Northern elephant seals (Mirounga angustirostris). J Comp Pathol. 2005;132:169-178. [ Links ]
16. Clarke EA, Goodship AE. A severely disabled mammoth - the palaeopathological evidence. Quaternary International. 2010; doi: 10.1016/j.quaint.2010.07.020. [ Links ]
17. Bartosiewicz L. Taphonomy and palaeopathology in archaeozoology. Geobios. 2009;40:69-77. [ Links ]
18. Dzierzęcka M, Charuta A, Kobryń H. Pathological changes of horse bones in the middle ages in Poland - photographic records. B Vet I Pulawy. 2008;52:689-694. [ Links ]
19. Coltman DW, Bowen WD, Boness DJ, Iverson SJ. Balancing foraging and reproduction in the male harbour seal, an aquatically mating pinniped. Anim Behav. 1997;54:663-678. [ Links ]
 Correspondence to:
Correspondence to:
Romala Govender
Postal address: Department of Zoology, University of Cape Town
Private Bag X3, Rondebosch 7701, South Africa
email: marinefossils@gmail.com
Received: 19 Apr. 2010
Accepted: 02 Nov. 2010
Published: 28 Jan. 2011


{kind=link}