










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.106 n.7-8 Pretoria Jul./Aug. 2010
RESEARCH LETTER
The effect of certain N-tritylated phenylalanine conjugates of amino-adenosine-3',5'-cyclic monophosphate on moloney murine leukaemia virus reverse transcriptase activity
Johann M. van ZylI; Mario AriattiII; Arthur O. HawtreyI
IPharmacology Division, Department of Medicine, Stellenbosch University, Tygerberg campus, South Africa
IIDepartment of Biochemistry, University of KwaZulu-Natal, Westville campus, South Africa
ABSTRACT
Moloney murine leukaemia virus (M-MuLV) is a member of the retrovirus family. Its cloned reverse transcriptase (RT), similarly to HIV type 1 reverse transcriptase (HIV-1 RT), exhibits DNA-polymerase and ribonuclease H (RNase H) activities capable of converting the single-stranded retroviral RNA genome into double-stranded DNA. The latter is then integrated into the host chromosome during viral infection. M-MuLV RT is, therefore, an attractive enzyme to help understand mutations in HIV-1 RT and its use in inhibition studies can help facilitate new drug designs. In this study, conjugates consisting of N-trityl derivatives of p-fluoro, p-nitro and p-iodo-DL-phenylalanine were coupled to 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate and examined for their effect on DNA synthesis by M-MuLV RT. Synthesis was studied in a system containing poly (rA).oligo d(pT)15 as a template-primer with [3H] dTTP. The iodo-derivative, N-trityl-p-iodo-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate was found to be a very active inhibitor of the RT enzyme (IC50 = 1 µM), while the p-nitro (IC50 = 45 µM) and p-fluoro (IC50 = 65 µM) were weak inhibitors. Further work will be aimed at determining the mode of binding of the N-tritylated conjugates and also of various substituted amino acids and short peptides to M-MuLV RT to elucidate the mechanisms of inhibition.
Keywords: inhibitor; Moloney murine leukaemia virus; N-tritylated phenylalanine-nucleotide; reverse transcriptase; ribose-sugar
INTRODUCTION
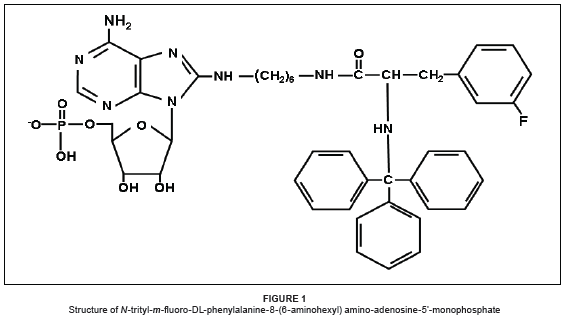
In a recent article1 we described the synthesis of a number of N-acylated derivatives of 8-(6-aminohexyl) amino-adenosine-5'-monophosphate. These various acylated derivatives, as well as the free N-trityl substituted DL-phenylalanyl amino acids, were tested for their effect on the DNA polymerase activity of the Moloney murine leukaemia virus reverse transcriptase (M-MuLV RT). A number of these compounds inhibited the enzyme with IC50 values (the concentration at which 50% inhibition of activity occurs) of 60 µM - 100 µM. If one particular compound, N-trityl-m-fluoro-DL-phenylalanine, was attached to the nucleotide 8-(6-aminohexyl) amino-adenosine-5'-monophosphate (Figure 1), the inhibitory activity increased, giving an IC50 value of 5 µM.
M-MuLV RT is a cloned enzyme expressed in Escherichia coli as a single polypeptide (71 kDa) 2 and, similarly to HIV type 1 reverse transcriptase (HIV-1 RT), exhibits both DNA-polymerase and ribonuclease H (RNase H) activities3; the common properties of M-MuLV RT and HIV-1 RT suggest that the two enzymes are able to carry out identical biological functions as far as the mechanism of DNA synthesis is concerned. Furthermore, the amino acid sequences of several drug-resistant mutations of HIV-1 RT have revealed similarities to the native sequences found in the M-MuLV RT catalytic fragment,4 which makes M-MuLV RT an attractive enzyme to help understand mutations in HIV-1 RT and its use in inhibition studies can help facilitate new drug designs.
In our previous paper,1 we made the assumption that the nucleotide component (5'-AMP) of the conjugate N-trityl-m-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-5'-monophosphate could possibly bind to the polymerase site (area) of M-MuLV RT, while the N-trityl-m-fluoro-DL-phenylalanine grouping was likely to interact with a hydrophobic pocket or area situated 10 Å - 15 Å away. The binding of the conjugate therefore resembled the interaction of certain chimeras with HIV-1 RT. We now realise that the suggested binding of the N-trityl conjugates mentioned above may have been entirely wrong and that other areas or sites on the RT enzyme may possibly have been involved (see Discussion).
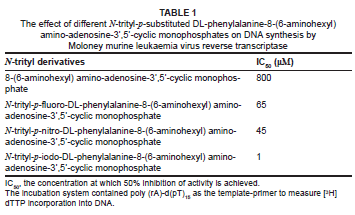
It was therefore of particular interest to see if replacement of the nucleotide adenosine-5'-phosphate component of N-trityl-p-substituted-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-5'-monophosphate by adenosine-3',5'-cyclic monophosphate would result in inhibitory derivatives against the reverse transcriptase. This paper outlines the synthesis of three compounds containing 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate that are linked to different N-trityl-p-substituted-DL-phenylalanines (p-fluoro, p-nitro and p-iodo; Figure 2) and their effects on the DNA polymerase activity of M-MuLV RT.
It would have been logical to use m-derivatives (m-fluoro, m-nitro and m-iodo) of DL-phenylalanine in these new conjugates with 3',5'-cyclic-AMP. Our reason for using the N-tritylated-p-substituted DL-phenylalanine derivatives was that we had adequate amounts of crystalline N-hydroxysuccinimide-activated esters of these N-tritylated-p-substituted amino acids, while the m-substituted derivatives were not available.
MATERIALS AND METHODS
Poly (rA) and 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate were supplied by Sigma (St Louis, United States of America). Oligo d(pT)15 and M-MuLV RT were supplied by Roche (Mannheim, Germany). The [3H] dTTP (65.5 Ci/mmol) used as the tetrasodium salt was obtained from American Radiolabeled Chemicals (St Louis, USA). All other chemical reagents were of analytical grade.
We synthesised three new conjugates, as detailed below. The N-tritylated amino acids used were N-trityl-p-fluoro-DL-phenylalanine, N-trityl-p-nitro-DL-phenylalanine and N-trityl-p-iodo-DL-phenylalanine. The N-tritylated phenylalanines were activated as their N-hydroxysuccinimide derivatives, which were then coupled to 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate (Figure 2). N-trityl-p-fluoro-DL-phenylalanine, N-trityl-p-nitro-DL-phenylalanine and N-trityl-p-iodo-DL-phenylalanine were prepared as described previously by Nel and Ariatti5, using the method of Zervas and Theodoropoulos6. The N-hydroxysuccinimide derivatives of the above compounds were prepared according to the procedure of Nel and Ariatti5.
Assay for RNA-dependent DNA polymerase activity of M-MuLV RT
The incubation system contained 50 mM Tris-HCl (pH 8.3), 20 mM KCl, 10 mM dithiothreitol, 5 mM MgCl2, reverse transcriptase (10 units), poly rA (1.0 µg), d(pT)15 primer (0.1 µg), [3H] dTTP (1.25 µCi) and dTTP (0.5 µg), in a final volume of 20 µL. Reactions were carried out at 37 ºC. Aliquots (4 µL) were removed at regular 1 min intervals after commencing the run and mixed into 20 µL 0.1 M EDTA (pH 7.0). From these solutions, aliquots (7 µL), were removed and streaked onto discs of DE-81 paper, dried at 37 ºC and then placed in 0.5 M sodium phosphate (pH 6.8) for 5 min. This procedure was repeated with fresh 0.5 M phosphate. Discs were dried at 60 ºC and radioactivity was assessed by liquid scintillation. Test compounds were dissolved in ethanol or 0.05 M Tris-HCl (pH 7.6). The final concentration of ethanol in test runs and controls was 10% (v/v).
N-trityl-p-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate
To a solution of 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate (4.7 mg, 0.01 mmol) in 0.8 mL pyridine:H2O (7:3, v/v) was added to the N-hydroxysuccinimide ester of N-trityl-p-fluoro-DL-phenylalanine (5.3 mg, 0.01 mmol) in 0.4 mL pyridine:H2O (7:3, v/v). The reaction mixture was left in the dark at room temperature (20 ºC) for 20 h. It was then taken to dryness under a stream of nitrogen at 30 ºC. Traces of pyridine were removed by co-evaporation with toluene using nitrogen. The crude product was dissolved in 0.8 mL dioxane:ethanol:H2O (2:2:1, v/v) at 30 ºC and chromatographed on silica gel 60 F254 plates (20 cm × 20 cm) using CHCl3:CH3OH (4:1, v/v). The strongly absorbing UV band in the area around the origin was extracted with ethanol (7.0 mL) at 30 ºC and filtered through Whatman No. 1 paper.
Chromatography of an aliquot of the filtered extract on silica gel 60 F254 using ethanol:H2O (5:1, v/v) produced a new single strongly absorbing UV spot (retardation factor [RF] = 0.6), which was positive for the trityl group (2% aqueous HClO4, 100 ºC). The starting material 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate produced an RF value of 0.26, while N-hydroxysuccinimide had an RF value of 0.49.
On UV examination, with ethanol as a solvent, the new compound, N-trityl-p-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate, was found to have a maximum wavelength (λmax) of 274 nm and a 274/243 ratio of 2.98. The mass spectrum of the compound was measured in methanol containing 0.1% formic acid. The precursor ion of the new compound had a molecular mass of 869.5 Da and product ions with mass-to-charge (m/z) ratios of 243.1 (trityl), 299.1, 353.3, 381.4, 409.3, 537.4 and 627.1 were found.
N-trityl-p-nitro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate
This compound was prepared similarly to the method described above for the p-fluoro derivative. The synthesis used 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate (4.7 mg, 0.01 mmol) and the N-hydroxysuccinimide ester of N-trityl-p-nitro-DL-phenylalanine (5.6 mg, 0.01 mmol). Following isolation by thin layer chromatography (TLC), the final product was dissolved in ethanol, giving a single spot on TLC of RF = 0.71 using silica gel 60F254 and ethanol:H2O (5:1, v/v). With ethanol as a solvent, the UV had a λmax of 274 nm, a 274/244 ratio of 2.54 and a mass spectrum m/z of 245.1 (trityl), 299.2, 333.3, 413.3, 671.5, and 879.6 (molecular mass = 879.6).
N-trityl-p-iodo-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate
This compound was also similarly prepared as described above for the p-fluoro derivative. The synthesis used 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate (4.7 mg, 0.01 mmol) and the N-hydroxysuccinimide ester of N-trityl-p-iodo-DL-phenylalanine (6.4 mg, 0.01 mmol). Following isolation by TLC, the pure derivative was dissolved in ethanol and gave a single spot on TLC (silica gel 60F254), with an RF of 0.78 using ethanol:H2O (5:1, v/v). With ethanol as a solvent, the UV had a λmax of 273 nm and a 273/244 ratio of 2.13. When run in negative mode, the mass spectrum m/z ratios were 243.1 (trityl), 444.2, 717.1, 734.6 and 960.3 (molecular mass = 960.5). In positive mode, the mass spectrum results showed m/z ratios of 442.1, 510.1, 957.1 and 958.1 (molecular mass = 960.5).
RESULTS AND DISCUSSION
Previous work from our laboratory1 described the effects of various N-trityl amino acids, as well as the conjugate compound N-trityl-m-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-5'-monophosphate, on the activity of enzyme-MuLV RT (Figure 1). The N-tritylated amino acids all inhibited the reverse transcriptase with IC50 values varying between 95 µM and 65 µM. When N-trityl-m-fluoro-DL-phenylalanine was chemically coupled to the 6-amino-side chain of 8-(6-aminohexyl) amino-adenosine-5'-monophosphate, however, the resulting conjugate showed a dramatic increase in inhibitory activity, with an IC50 value of 5 µM; we have previously discussed possible reasons for this and also suggested how the conjugate might bind to the reverse transcriptase enzyme.1
In this study, we wished to examine whether the replacement of the adenosine-5'-monophosphate component of N-trityl-m-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-5'-monophosphate with 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate would result in a conjugate with an inhibitory effect on the reverse transcriptase enzyme. We therefore synthesised three new conjugates: N-trityl-p-fluoro-DL-phenylalanine, N-trityl-p-nitro-DL-phenylalanine and N-trityl-p-iodo-DL-phenylalanine.
Assays for reverse transcriptase activity (reported as IC50 values) were carried out using poly (rA)-d(pT)15 as the template-primer and measuring incorporation of [3H]dTTP into newly synthesised DNA (poly dT)7 (Table 1). The p-iodo derivative was clearly a good inhibitor (IC50 = 1 µM), while the p-nitro (IC50 = 45 µM) and p-fluoro (IC50 = 65 µM) derivatives were weak inhibitors. The molecular/kinetic reasons for these differences are not apparent at present. It is worth noting that the 8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate that was run as a control resulted in a weak inhibition of the reverse transcriptase enzyme (IC50 = 800 µM).
The strongly inhibitory activity of the p-iodo derivative is interesting; the iodine atom has a large radius (covalent radius = 1.33 Å) and is lipophilic, whilst fluorine and bromine have covalent radii of 0.72 Å and 1.14 Å, respectively. The large covalent radius of the iodine atom may have had some bearing on the activity of the nucleotide conjugate.
Cushman and colleagues8 synthesised a number of HIV-1 reverse transcriptase inhibitors, of which one of the most active was 3',3''-dibromo-4',4''-dimethoxy-5',5''-bis(methoxycarbonyl)-1,1-diphenyl-1-heptene. They reported that replacement of the two bromine atoms in the above compound with two chlorine atoms resulted in a compound with decreased HIV-1 RT inhibitory activity.
In this study, the compounds synthesised and evaluated as inhibitors of the M-MuLV RT were originally visualised as chimeras. Several other researchers9,10,11,12 originally suggested the idea of cross-linking a non-nucleoside compound, specifically a non-nucleoside reverse transcriptase inhibitor (NNRTI), to substrates (i.e. the formation of a chimera). As reported in our previous paper,1 the design features of the nucleotide-N-trityl-m-fluoro-DL-phenylalanine appear to be consistent with these ideas. The concept of chimera structures binding to M-MuLV RT was prominent in our thinking when we began to carry out the work reported in the present paper. We now believe that the evidence for a chimera-type inhibitor and the mechanism we proposed, that is, of conjugates binding to M-MuLV RT, is incorrect, for the reasons outlined below.
Binding of N-trityl-p-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-3',5'-cyclic monophosphate and the p-nitro and p-iodo substituted DL-phenylalanine conjugates to M-MuLV RT
HIV-1 RT is possibly the most extensively studied enzyme and a multitude of studies on its crystal structure have been reported.9,12 Although there are distinct structural differences between the monomeric M-MuLV RT and the heterodimeric HIV-1 RT, it is important to keep in mind that the respective polymerase domains of M-MuLV RT and HIV-1 RT are approximately 85% structurally similar.13,14 This similarity allows the use of M-MuLV-RT as a tool in order better to understand the reverse transcription process and the design of novel inhibitors for HIV-1 RT. We do not know how the M-MuLV RT binds NNRTIs, whether there is a hydrophobic pocket present, or whether the inhibitor can induce allosteric changes in the enzyme structure. It is also unlikely that the nucleotide (5'-AMP or 3',5'-cyclic-AMP) component of the conjugate is binding near, or at, the polymerase site, as these are not deoxyribose nucleoside triphosphates but ribose-containing nucleotides. Also, inhibition, when it occurs, should be non-competitive. When using N-trityl-m-fluoro-DL-phenylalanine-8-(6-aminohexyl) amino-adenosine-5'-monophosphate in our original paper,1 we performed competition experiments using varying concentrations of poly rA, d(pT)16 at fixed concentrations of N-trityl-m-fluoro-DL-phenylalanine-nucleotide, which showed the compound acted as a non-competitive inhibitor (results not published).
It is thus unlikely that the DL-phenyl conjugates would fit into both the polymerase binding area and the nearby allosteric pocket site. Based on our results, we propose that other areas of the M-MuLV RT enzyme are being used for the attachment and functioning of the N-tritylated conjugates.
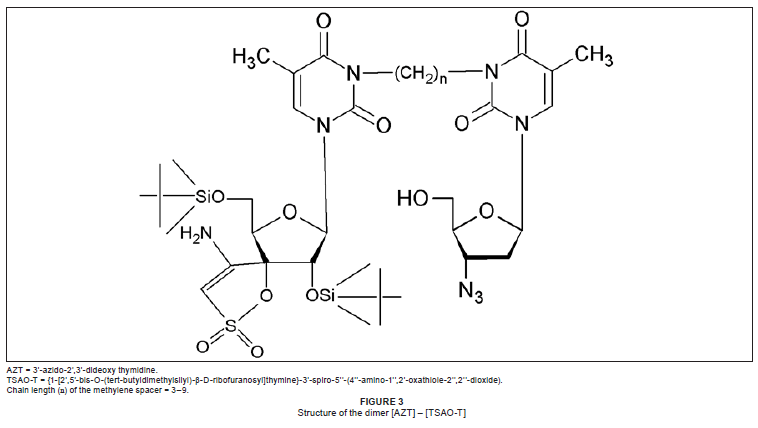
A number of research groups have reported on the synthesis and biological evaluation of dimers, some of which behave as true chimeras.15,16,17,18,19 Velázquez et al.15,16 have prepared a number of dimers consisting of 3'-azido-2',3'-dideoxy thymidine (AZT) linked to the non-nucleoside (NNRTI) TSAO or HEPT. These can be written as [AZT]-[TSAO-T] and [AZT]-[HEPT]. They can also be placed in a general formula: [ddN]-(CH2)n-[NNRTI], where n = 3-9.
The TSAO-T molecule does not bind in the hydrophobic pocket of the HIV-1 RT, but rather interacts with certain amino acids at the interface of the p51 and p66 subunits. Velázquez et al.15 believe that the [AZT]-[TSAO-T] dimer works only through the NNRTI portion of the molecule and not through the AZT (ddN) segment. It should be noted that TSAO is also a ribose derivative. The [AZT]-(CH2)n-[TSAO-T] structure is shown in Figure 3.
A recent paper by Tomassi et al.20 deals with the synthesis of new TSAO-T derivatives, particularly TSAO-Boc3T. The introduction of a Boc grouping to the N-3 position of the thymidine ring creates a good inhibitor of HIV-1 RT activity. Besides details of the synthesis and biological evaluation of this inhibitor, they report on computational analysis regarding the binding site of the enzyme.20
Our further work will be aimed at determining the mode of binding of the N-tritylated conjugates and also of various substituted amino acids and short peptides to M-MuLV RT, in order to resolve the abovementioned uncertainties.
ACKNOWLEDGEMENTS
We gratefully acknowledge the financial support given by the Harry Crossley Foundation. We also thank Dr H. Seifart for the mass-spectra and the anonymous reviewers for their constructive criticism and interesting concepts.
REFERENCES
1. Hawtrey A, Pieterse A, Van Zyl J, et al. Studies on the inhibition of Moloney murine leukemia virus reverse transcriptase by N-tritylamino acids and N-tritylamino acid-nucleotide compounds. Nucleosides Nucleotides Nucleic Acids. 2008;27:1011-1023. [ Links ]
2. Roth MJ, Tanese N, Goff SP. Purification and characterization of murine retroviral reverse transcriptase expressed in Escherichia coli. J Biol Chem. 1985;260:9326-9335. [ Links ]
3. Grandgenett DP, Gerard GF, Green M. A single subunit from avian myeloblastosis virus with both RNA-directed DNA polymerase and ribonuclease H activity. Proc Natl Acad Sci USA. 1973;70:230-234. [ Links ]
4. Georgiadis M, Jessen SM, Ogata CM, Telesnitsky A, Goff SP, Hendrickson W. Mechanistic implications from the structure of a catalytic fragment of Moloney murine leukemia virus reverse transcriptase. Structure. 1995;15:879-892. [ Links ]
5. Nel W, Ariatti M. Peptidyltransferase substrate specificity in E. coli and rat liver ribosomes with aromatic aminoacyl analogues of puromycin. S Afr J Sci. 1982;78:202-204. [ Links ]
6. Zervas L, Theodoropoulos DM. N-Tritylamino acids and peptides: A new method of peptide synthesis. J Am Chem Soc. 1956;78:1359-1363. [ Links ]
7. Maniatis T, Fritsch EF, Sambrook J. Molecular cloning. A laboratory manual. Cold Spring Harbor: Cold Spring Harbor Laboratory Press, 1989; p. 473. [ Links ]
8. Cushman M, Golebiewski WM, Graham L, et al. Synthesis and biological evaluation of certain alkenyl-diarylmethanes as anti-HIV-I agents which act as non-nucleoside reverse transcriptase inhibitors. J Med Chem. 1996;39:3217-3227. [ Links ]
9. Smerdon SJ, Jager J, Wang J, et al. Structure of the binding site for non-nucleoside inhibitors of the reverse transcriptase of human immuno-deficiency virus type I. Proc Natl Acad Sci USA. 1994;91:3911-3915. [ Links ]
10. Pata JD, Stirtan WG, Goldstein SW, Steitz TA. Structure of HIV-1 reverse transcriptase bound to an inhibitor active against mutant reverse transcriptases resistant to other non-nucleoside inhibitors. Proc Natl Acad Sci USA. 2004;101:10548-10553. [ Links ]
11. Nanni RG, Ding J, Jacobo-Molina A, Hughes SH, Arnold E. Review of HIV-1 reverse transcriptase three-dimensional structure: Implications for drug design. Perspect Drug Discov Des. 1993;1:129-150. [ Links ]
12. Das K, Bauman JD, Clark AD Jr, et al. High-resolution structures of HIV-1 reverse transcriptase/TMC278 complexes: Strategic flexibility explains potency against resistance mutations. Proc Natl Acad Sci USA. 2008;105:1466-1471. [ Links ]
13. Pandey PK, Kaushik N, Talele TT, Yadav PNS, Pandey N. The β7-β8 loop of the p51 subunit in the heterodimeric (p66/p51) human immunodeficiency virus type 1 reverse transcriptase is essential for the catalytic function of the p66 subunit. Biochemistry. 2001;40:9505-9512. [ Links ]
14. Coté ML, Roth MJ. Murine leukemia virus reverse transcriptase: Structural comparison with HIV-1 reverse transcriptase. Virus Res. 2008;134:186-202. [ Links ]
15. Velázquez S, Alvarez R, San Felix A, et al. Synthesis and anti-HIV activity of [AZT]-[TSAO-T] and [AZT]-[HEPT] dimers as potential multifunctional inhibitors of HIV-1 reverse transcriptase. J Med Chem. 1995;38:1641-1649. [ Links ]
16. Velázquez S, Tuñón V, Jimeno ML, et al. Novel series of [ddN]-[TSAO-T] heterodimers as potential bi-functional inhibitors of HIV-1 RT: Studies in the linker and ddN region. Nucleosides Nucleotides. 1999;4-5:1029-1030. [ Links ]
17. Pontikis R, Dollé V, Guillaumel J, et al. Synthesis and evaluation of 'AZT-HEPT', 'AZT-pyridinone' and 'ddC-HEPT' conjugates as inhibitors of HIV reverse transcriptase. J Med Chem. 2000;43:1927-1939. [ Links ]
18. Mavromoustakos T, Calogeropoulou T, Koufaki M, et al. Ether phospholipid-AZT conjugates possessing anti-HIV and antitumor cell activity. Synthesis, conformational analysis, and study of their thermal effects on membrane bilayers. J Med Chem. 2001;44:1702-1709. [ Links ]
19. Kucera LS, Morris-Natschke SL, Ishaq KS, et al. Synthesis and evaluation of a novel synthetic phosphocholine lipid-AZT conjugate that double-targets wild-type and drug resistant variants of HIV. Nucleosides Nucleotides Nucleic Acids. 2004;23:385-399. [ Links ]
20. Tomassi C, Van Nhien AN, Marco-Contelles J, et al. Synthesis, anti-HIV-1 activity, and modeling studies of N-3 Boc TSAO compound. Bioorg Med Chem Lett. 2008;18:2277-2281. [ Links ]
 Correspondence to:
Correspondence to:
Johann van Zyl
Postal address: Pharmacology Division, Department of Medicine, Stellenbosch University
PO Box 19063
Tygerberg 7505, South Africa
email: jmvzyl@sun.ac.za
Received: 28 Jan. 2010
Accepted: 6 Apr. 2010
Published: 07 July 2010
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}