










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.106 no.5-6 Pretoria Mai./Jun. 2010
RESEARCH LETTER
Creation of a high-yielding recombinant maize hybrid for the production of a microbicide to prevent HIV-1 transmission
Eugenia BarrosI; Sydney W. NelsonII
ICSIR Biosciences, Pretoria, South Africa
IINelson Genetics CC, Bryanston, South Africa
ABSTRACT
The aim of this study was to use conventional breeding to increase the production in maize of the human monoclonal antibody 2G12, known to have potential therapeutic properties in the prevention of HIV-1 transmission. The recombinant antibody, together with a fluorescent marker, was introduced into two South African high-performing maize elite inbred lines by crossing them with a transgenic maize line that had been transformed with the monoclonal antibody 2G12. The effect of breeding to produce high-expressing recombinant hybrid seed was evaluated by comparing 2G12 production in the different breeding lines with the original maize line. 'Good production practice standards' were followed throughout the breeding programme. 'Conventional drug regulations' adapted to plant-made pharmaceuticals were also followed, with the seeds being stored in a 'master seed bank'. The maize hybrid expressed a higher level of the antibody than the recombinant maize elite lines. This plant-derived antibody provides a means of producing a microbicide component that could be used with other HIV-neutralising antibodies as an additional approach to prevent HIV infection.
Keywords: 2G12; biopharming; HIV-1; microbicide; monoclonal antibody
INTRODUCTION
HIV- and AIDS-related infections are possibly the primary cause of death in sub-Saharan Africa and are also a leading cause of mortality worldwide, with an estimated two million deaths in 2008 alone. In that year, the number of people living with HIV/AIDS worldwide was estimated to be 33.4 million, of which 2.7 million were newly infected with HIV.1 Although various vaccination strategies have been pursued in an attempt to halt the HIV pandemic, the complex biology of the virus makes it a difficult target for vaccine protection and no effective vaccine has been developed as yet. According to Letvin2, HIV is probably transmitted both as a cell-free virus and as a cell-associated virus. Cell-free viruses can only be eliminated through binding to neutralising antibodies, whereas cell-associated viruses are mainly eliminated by cell-mediated immune responses. For a vaccine to be effective, it must elicit both types of immune responses accordingly. Microbicides, on the other hand, can bypass many of the immunological challenges associated with HIV vaccine development and can thus be used in the form of topical gels that offer an additional strategy for the prevention of HIV transmission.
Monoclonal antibodies able to neutralise primary isolates of HIV type 1 (HIV-1) are important candidates for passive immunisation regimens that aim to reduce the infectivity of the virus. HIV-1 neutralisation has been shown to be mediated by antibodies that bind to certain epitopes found on the virus envelope. The envelope is made up of a complex of two glycoproteins, gp120 and gp41.3 The human monoclonal antibody 2G12 is a broadly neutralising antibody that recognises a unique epitope on the surface of gp120 and binds to a cluster of oligomannose-type N-glycans. When antibody cocktails made up of 2G12 and two other neutralising antibodies, 4E10 and 2F5, were tested parenterally on humans who had had their antiretroviral treatment stopped, a delay in viral rebound was observed.4 Similar results were obtained in monkeys, where passive administration of antibodies prevented infection by simian HIV. This illustrates the role that antibodies can play in containing the spread of the virus. Very high levels of the antibodies need to be administered for effective protection, however, and their large-scale production using mammalian cell systems is very costly and not feasible for poorer countries.
Plant-based expression systems have been developed as alternative vehicles for the safe and economic production of pharmaceutical proteins including therapeutic recombinant monoclonal antibodies.5 The suitability of maize as a production host for a fully functional 2G12 antibody was demonstrated by Rademacher et al.6 The antibody was expressed in the maize endosperm and a lysine-aspartate-glutamate-leucine (KDEL) tetrapeptide signal was attached to the C-terminus of the glycoprotein to retain it in the endoplasmic reticulum so that the antibody would be devoid of plant-specific complex-type N-glycans. This antibody-producing maize has been crossed with other maize lines, including two South African elite maize inbred lines, NSP5120A and SSG62B. In this study we demonstrate the breeding of these two transgenic maize lines containing 2G12, aimed at the increased production of the 2G12 antibody under greenhouse containment, and confirm the expression of the antibody during all stages of breeding. Good agricultural practice and good manufacturing practice guidelines were followed and a 'master seed bank' was created for the long-term storage of the transgenic seed.
METHODS
Plant material
The transgenic South African elite lines, SSG62B and NSP5120A, containing the stable mAb 2G12 derived from crosses made with primary Hi-II maize transformants that contained 2G12,6 were used in this study for breeding and increased production of 2G12. The genes for the two 2G12 antibody chains were obtained from Polymum (Vienna, Austria) and the recombinant antibody 2G12 was shown efficiently to neutralise HIV-1.6
This antibody was expressed in the endosperm, together with the visual fluorescent marker DsRed, to facilitate selection during breeding of the transgenic maize and to serve as an additional biosafety precaution in facilitating identification of the transgenic maize from non-transgenic maize.
Breeding and selection of 2G12 transgenic maize plants for the increased production of 2G12 under greenhouse containment
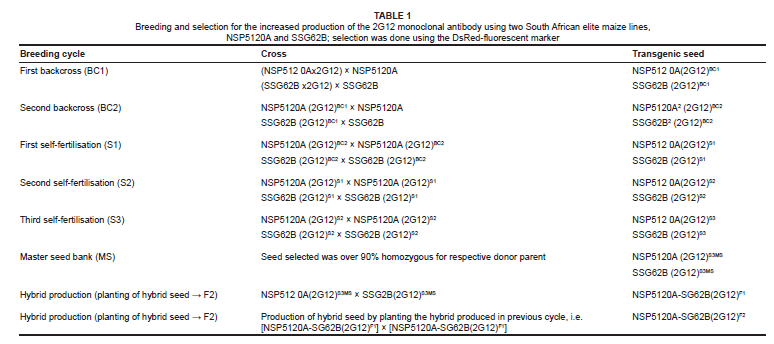
The two South African transgenic maize lines containing the monoclonal antibody 2G12 were backcrossed twice to their respective original elite line (donor/recurrent parent) followed by three generations of self-fertilisation (S1 to S3) and selection. Selection was done using the DsRed-fluorescent marker, where a green light and a red filter were used to detect DsRed-containing maize kernels, while selection on agronomic characteristics among the backcross progenies was performed simultaneously. At the end of S3 generation, over 90% homozygosity of the recurrent parent phenotype was obtained for each of the parent backgrounds, NPS5120A and SSG62B, and this seed was stored at the master seed bank, from which seed was also used for hybrid production (F1 and F2). Hybrids were produced by crossing homozygous inbred seeds of SSG62B-2G12 and NSP5120A-2G12, constituting the F1 generation. The harvested seed was then planted to produce F2 hybrid seed, constituting the F2 generation. F1 hybrid plants and S3 inbred plants were grown separately in adjacent compartments in the greenhouse under identical conditions. After each breeding cycle the seeds were harvested, screened for the presence of the fluorescent marker, tested for the expression of 2G12 using an enzyme-linked immunosorbent sandwich assay (ELISA) and then stored in heat-sealed packets. The packets were properly labelled with a batch code and the information entered in a detailed database.
The cultivation of both transgenic maize and control maize plants took place under greenhouse containment level 2 according to the biosafety regulations for growing genetically modified organisms in South Africa.7 Standard greenhouse procedures were followed to produce a uniform maize crop. As the monoclonal antibody 2G12 is aimed for human application, additional guidelines were followed which are similar to those prescribed by the European Agency for the Evaluation of Medicinal Products.8 These guidelines were followed during all the stages of plant cultivation, collection, harvesting, packaging and storage, as well as during the purification of the monoclonal antibody.
Quantification of 2G12 expression in maize seeds
Transgenic maize kernels from three generations of self-fertilisation and from two generations of hybrid production were milled to a fine powder using a small coffee grinder. The total soluble protein was extracted overnight using three volumes of phosphate-buffered saline (PBS). Insoluble material was removed by centrifugation at 8000 g for 15 min at 4 ºC. This was done twice and the supernatant saved each time. The expression and quantification of 2G12 present in the final supernatant was determined by an ELISA. Total protein was also extracted from maize kernels from non-transformed maize plants. Flat-bottom microtiter plates (NUNC, Roskilde, Denmark) were coated overnight at 4 ºC with goat anti-human light chain (K3502) and goat anti-human heavy chain (I2136) antibodies (Sigma, St. Louis, United States of America) at a concentration of 20 µg/mL. Subsequent incubation steps were performed at 37 ºC. Coated plates were washed twice with PBS supplemented with 0.5% Tween (PBST) and blocked for 2 h with PBS supplemented with 5% skim milk. Plates were washed again six times with PBS and subsequently incubated for 2 h with a standard, as well as transgenic and non-transformed maize samples diluted 1:2 in PBS. The standard was a human IgG (BP078) mAb (The Binding Site, Birmingham, United Kingdom) serially diluted in PBS (starting at a concentration of 100 ng/mL). Plates were washed six times with PBST and then incubated with diluted (1:100 in PBS) alkaline phosphatase-conjugated goat anti-human IgG A3813 or A9544 (Sigma) for 2 h. Plates were washed again six times with PBST, followed by incubation with pNPP substrate (Sigma). The optical density was measured at 405 nm after 20 min. Apparent affinities were calculated using Microsoft Excel.
Purification of 2G12 (from hybrid seed) using Protein-A agarose affinity purification
Total soluble protein (3 mL) containing 2G12 at a concentration of 41.12 µg/mL (crude extract) was diluted with 3 mL of binding buffer (Affi-Gel Protein A MAPS II Kit, Bio-Rad Laboratories, Hercules, USA). The diluted sample was applied to a 300µL Affi-Gel protein agarose column that was equilibrated with binding buffer. Once the first eluate (flow-through fraction) was passed through, the column was washed with 15 bed volumes of binding buffer (1st wash fraction). The antibody was then eluted with 5 bed volumes of elution buffer (purified extract) and the column eluted again with 10 bed volumes elution buffer to ensure total removal of 2G12 (2nd wash fraction). In both cases, the eluted antibody was neutralised with 1 mol Tris HCl, pH 9 as specified in the protocol. The concentration of 2G12 was quantified in the different fractions using ELISA (described below).
SDS-PAGE, ELISA and western blot
Crude plant extracts from untransformed maize were used as negative controls to test for interference of maize plant extracts with the ELISA protocols. Purified and crude extracts of 2G12 mAb and eluates generated during the purification of the 2G12 monoclonal antibody were analysed by SDS-polyacrylamide gel electrophoresis (SDS-PAGE). Electrophoresis was performed in a Mini-Protean electrophoresis unit (Bio-Rad Laboratories) using a 12% acrylamide separating gel and a 4% acrylamide stacking gel. The proteins were transferred electrophoretically from the gel to a methanol pre-treated Sequi-Blot PVDF membrane (Bio-Rad Laboratories) in a Trans-Blot semi-dry transfer cell module (Bio-Rad Laboratories). After the transblotting, the membranes were incubated in PBST supplemented with 5% skim milk powder for 1 h, followed by incubation with diluted (1:5000 in PBST) alkaline phosphatase-conjugated anti-human IgG (Bio-Rad Laboratories) supplemented with 2.5% skim milk powder overnight. The membranes were washed three times for 10 min with PBST buffer. Luminometric detection was performed using the BCIP/NBT Chemiluminescent Reagent (Roche, Indianapolis, USA) according to manufacturer's instructions. All the steps were carried out at room temperature.
RESULTS
Expression and quantification of 2G12 in transgenic maize seeds Expression levels of 2G12 extracted from whole maize seeds were quantified using ELISA. The evaluation experiments showed that the maize seed extract did not influence the ELISA performance. The extracted yield of 2G12 antibody varied from 42 µg/g dry seed weight to 55 µg/g dry seed weight in the first self-fertilising generation of breeding (S1). A lower amount of extractable 2G12 was found in the SSG parental background in comparison with the NSP background. The seeds that were part of the maize cobs with higher antibody expression levels were selected for the following cycle of breeding. The co-segregation of DsRed with both antibody chains made the selection of seeds from each breeding cycle easier and more accurate (Figure 1). Some of the white kernels were also tested by ELISA together with the DsRed kernels; no 2G12 expression was observed in the case of the white kernels (results not shown). The intensity of the red fluorescence in the DsRed-containing kernels had no correlation with the level of 2G12 in the kernel. The 2G12 antibody levels were determined at the end of each breeding cycle (see next section). The antigen-binding properties of 2G12 produced in maize were compared with 2G12 produced in Chinese hamster ovary cells using Biacore analysis; this, and the antigen-binding assays, confirmed that the monoclonal antibody was fully assembled and had proper antigen-binding activity (Rademacher T 2006, personal communication, June 6).

Increased production of monoclonal antibody 2G12 by breeding, under greenhouse containment
The expression of 2G12 in maize seeds at the end of each breeding cycle was quantified using ELISA. In both transgenic lines (NSP and SSG origin), the antibody content was slightly lower in the S1 generation (about 42 µg/g - 55 µg/g dry seed weight) than in the following generations, probably due to a higher gene dose expected when homozygosity was reached. The antibody level was stabilised throughout S2 and S3 for both lines, varying between 60 µg/g and 67 µg/g dry seed weight for NSP background and between 56 µg/g and 58 µg/g dry seed weight for SSG background. In order to have S3 seed expressing higher levels of the antibody, higher S1 expressers could have been selected through breeding in future generations; this was not done in this study. The seed was stored in the cold room as the master seed bank, defined as containing homozygous seed (over 90% homozygosity) expressing the trait of interest from which all future increased productions can be initiated to ensure uniformity and consistency of the product. This is the basis of any plant-made pharmaceutical production platform.
Hybrids (F1) were produced by crossing seeds of these two transgenic lines with the hybrid seed. These were then planted to produce hybrid progeny (F2). The rationale for using hybrids was to increase crop yields because hybrids produce higher yields than 'selfed' inbred lines. The amount of 2G12 produced by the hybrid seed averaged 68 µg/g dry seed weight (Figure 2). Although the amount of 2G12 in the hybrid seed was not substantially higher than that obtained for the two individual lines, the advantage of hybrid production from crosses of two elite inbred lines is that this combines important agronomic traits and advantageous alleles that allow a constant and stable high yield of maize cobs even when grown under greenhouse containment. The F1 hybrid plants were uniformly more vigorous than the S3 inbred plants in all phenotypically observable traits, including plant height, stalk diameter, crop size and tassel size. Table 1 shows the crosses made at each breeding cycle. Hybrid seed progeny (F1, 38 kg) were also stored at the master seed bank in heat-sealed packets at 4 ºC.
Purification of 2G12 from maize kernels using Protein-A agarose affinity purification
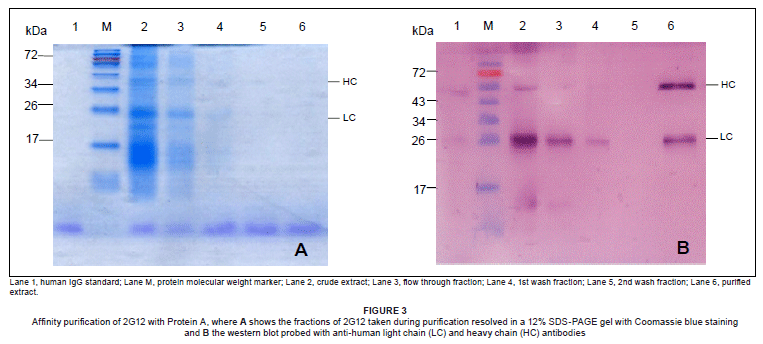
The crude extract of 2G12 monoclonal antibody (3 mL) was purified using the Affi-Gel Protein A MAPS II Kit (Bio-Rad Laboratories). Fractions from each purification step were quantified by ELISA. To determine if there was any antibody loss during the purification steps, the fractions were analysed by SDS-PAGE gel (Figure 3A) and by western blot (Figure 3B) to confirm the presence of light and heavy antibody chains, especially in the crude extract and in the last fraction containing most of the purified monoclonal antibody. The concentration of the purified monoclonal antibody was estimated to be 13.98 µg/mL (13.98 µg/g of dry seed weight) which represented 30% of total antibody present in the crude extract (42.12 µg/mL).
DISCUSSION
The conventional technology for producing large quantities of a given monoclonal antibody is expensive and researchers have investigated plants as possible 'factories' for lower-cost production of antibodies. Seed-based production platforms have also been investigated for plant-made pharmaceuticals, with the best yields being reported for maize.6,9,10 One of the benefits of expressing a plant-made pharmaceutical in maize is the ability to use production strategies to increase the quantities of the desired pharmaceutical protein through higher yields while keeping the cost low in comparison with processes involving fermenters.
The use of the monoclonal antibody 2G12 as a microbicide for the prevention of HIV infection could be an alternative means of mitigating the HIV pandemic while the search for a vaccine continues. Although only small amounts of this monoclonal antibody would need to be produced for the initial preclinical and clinical studies, large amounts would eventually have to be produced if efficacy was demonstrated. In order to keep the costs of production of this microbicide low, genes encoding the 2G12 monoclonal antibody were introduced into maize6 constituting the functional recombinant 2G12 and crossed with two high-yielding South African elite inbred lines as the first step in the increased breeding programme. Uniform expression of the functional antibody was observed after the second cycle of self-fertilisation and throughout the third self-fertilisation cycle. The importance of producing hybrids is to increase the crop yield per hectare in field production where the number of grains per cob and grain size is greater than in selfed plants. This also impacts on the cost and space required to produce a certain yield of maize seeds.
The expression of 2G12 was evaluated at each stage of the breeding cycle by testing a representative seed sample using ELISA. In addition, a sample of the hybrid progeny was also purified using a Protein-A agarose affinity column and the purified recombinant antibody was characterised by SDS-PAGE and western blot analysis. These analyses confirmed the structural integrity of the 2G12 antibody, with the heavy chain having a molecular mass of 50 kDa and the light chain a molecular mass of 25 kDa. Furthermore, the antibody was shown to have proper antigen-binding activity (Rademacher T 2006, personal communication, June 6). There are different subtypes of HIV-1 that are prevalent in different geographic areas, with subtype C dominant in South Africa. Gray et al.11 reported that a 2G12 monoclonal antibody failed to neutralise different isolates of HIV-1 subtype C. They found that their isolates lacked glycan 295, rendering them resistant to 2G12. The 2G12 produced in this study will be evaluated, along with other monoclonal antibodies that target different epitopes of different subtypes of HIV-1, as a microbicide ingredient to be used as a cocktail for the passive immunisation of people worldwide.
CONCLUSION
The use of conventional breeding for the introduction of the transgenic monoclonal antibody 2G12 into two well-adapted South African maize lines, followed by hybrid production, has highlighted the potential of using an efficient breeding programme for the future production of plant-made pharmaceuticals. The co-segregation of the fluorescent DsRed marker with the antibody not only facilitated the selection of the transgenic seeds during breeding, but also secured the identity preservation of the transgenic seed. Selection of seeds expressing higher levels of 2G12 should be identified at the stage of self-fertilisation of transformed lines at the commencement of the breeding programme. The high yield that can be generated by open-field cultivation of maize hybrids would also represent a reduction in the acreage required, making it an improved technological solution towards the goal of realising the full potential of biopharming.
ACKNOWLEDGEMENTS
This work was supported by the South African Department of Science and Technology and the European Union project Pharma-Planta, Frame Work 6 Programme. The authors would also like to acknowledge the technical assistance of Dr Sabine Lezar, Mr Moses Mokoena and Mr Taola Shai.
REFERENCES
1. UNAIDS: Aids epidemic update 2009 [document on the Internet]. c2009 [cited 2010 May 15]. Available from: http://data.unaids.org/pub/Report/2009/JC1700_Epi_Update_2009_en.pdf [ Links ]
2. Letvin NL. Progress towards HIV vaccine. Annu Rev Med. 2005;56:213-223. [ Links ]
3. Fouts TR, Binley JM, Trkola A, Robinson JE, Moore JP. Neutralization of the human immunodeficiency virus type 1 primary isolate JR-FL by human monoclonal antibodies correlates with antibody binding to the oligomeric form of the envelope glycoprotein complex. J Virol. 1997;71:2779-2785. [ Links ]
4. Trkola A, Kuster H, Rusert P, et al. Delay of HIV-1 rebound after cessation of antiretroviral therapy though passive transfer of human neutralizing antibodies. Nat Med. 2005;11:615-622. [ Links ]
5. Ko K, Koprowski H. Plant biopharming of monoclonal antibodies. Virus Res. 2005;111:93-100. [ Links ]
6. Rademacher T, Sack M, Arcalis E, et al. Recombinant antibody 2G12 produced in maize endosperm efficiently neutralizes HIV-1 and contains predominantly single-GlcNAc N-glycans. Plant Biotechnol. 2008;6:189-201. [ Links ]
7. Genetically Modified Organisms Act No. 15 of 1997, South Africa [statute on the Internet]. c1997 [cited 2010 March 15]. Available from: http://www.nda.agric.za/docs/geneticresources/act15.htm [ Links ]
8. European Agency for the Evaluation of Medicinal Products. Good agricultural and collection practice for starting materials of herbal origin. EMEA/HMPC/246816/05; 2005. [ Links ]
9. Stöger E, Ma JK-C, Fischer R, Christou P. Sowing the seeds of success: Pharmaceutical proteins. Curr Opin Biotechnol. 2005;16:167-173. [ Links ]
10. Ramessar K, Rademacher T, Sack M, et al. Cost-effective production of a vaginal protein microbicide to prevent HIV transmission. Proc Natl Acad Sci USA. 2008;104:3727-3732. [ Links ]
11. Gray ES, Meyers T, Gray G, Montefiori DC, Morris L. Insensitivity of paediatric HIV-1 subtype C viruses to broadly neutralizing antibodies raised against subtype B. PLoS Med. 2006;3:1023-1031. [ Links ]
 Correspondence to:
Correspondence to:
Eugenia Barros
Postal address:
CSIR Biosciences
PO Box 395, Pretoria
0001, South Africa
email: ebarros@csir.co.za
Received: 25 Nov. 2009
Accepted: 16 Mar. 2010
Published: 26 May 2010
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}
{kind=link}