










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.106 no.5-6 Pretoria Mai./Jun. 2010
RESEARCH ARTICLE
PI3K-dependent ERK is involved in GM-CSF-mediated activation of progenitor cells in a neovascularisation model
Wei Zhao; Xiang-Hong Yang
Department of Pathology, Affiliated Shengjing Hospital, China Medical University, Shenyang, Liaoning 110004, China
ABSTRACT
The current study investigated the effects of the granulocyte-macrophage colony-stimulating factor (GM-CSF) on rat endothelial progenitor cell (EPC) biology. EPCs were isolated from rat bone marrow and experiments were carried out to determine the dose- and time-dependent effects of GM-CSF on angiogenesis, proliferation, viability, migration and vasculogenesis. EPCs were co-cultured with rat cardiac microvascular endothelial cells (CMECs) to determine the effect of GM-CSF on EPC angiogenesis in vitro. EPC proliferation and viability, migration, and vasculogenesis were measured by the respective application of the method of transcription and translation (MTT) assay, the Transwell chamber assay and the Matrigel assay. GM-CSF treatment stimulated EPC angiogenesis, proliferation, viability, migration and tubule formation in a dose- and time-dependent manner. Furthermore, GM-CSF treatment led to the upregulation of integrinβ2, vascular endothelial growth factor (VEGF) and p-extracellular signal-regulated kinase (p-ERK) 1/2 levels in EPCs. Moreover, two different phosphatidylinositol 3-kinase (PI3K) inhibitors (LY294002 and Wortmannin) partially inhibited the GM-CSF-mediated increase in p-ERK1/2 levels in EPCs. The ERK inhibitor, FR180204, dramatically inhibited GM-CSF-mediated stimulation of EPC proliferation, migration and vasculogenesis. Taken together, the results of the current study provide evidence that a PI3K-dependent ERK signalling pathway is, at least, partially responsible for the GM-CSF-mediated effects on EPC function.
Keywords: co-culture model; endothelial progenitor cell; extracellular signal-regulated kinase; granulocyte-macrophage colony-stimulating factor; neovascularisation; phosphatidylinositol 3-kinase
INTRODUCTION
Increasing evidence that endothelial progenitor cells (EPCs) play an important role in vasculogenesis has resulted in a paradigm shift in vascular biology, whereby angiogenesis (the formation of new blood vessels from local endothelial cell proliferation, migration, and remodelling) has been redefined to include the contribution of bone-marrow-derived EPCs.1,2,3,4,5 EPCs are recruited to sites of ischaemia, where they differentiate into mature endothelial cells and participate in the establishment of new blood vessels for collateral circulation. EPCs have been reported to contribute to as much as 25% of the number of endothelial cells in newly formed blood vessels6 and EPC transplantation in patients has been demonstrated to induce blood flow recovery in ischaemic limbs and increase myocardial viability after infarction.7,8,9,10,11
Granulocyte-macrophage colony-stimulating factor (GM-CSF), a well-known stimulator of haematopoietic progenitor cells and myeloid lineage cells, has been shown to exert a potent stimulatory effect on EPC kinetics. GM-CSF has been shown specifically to mobilise EPCs that contribute to both improved vascularisation of ischaemic hindlimbs in mice12 and myocardial collateral flow in patients with coronary artery disease.13 In rabbits, EPCs mobilised by GM-CSF accelerated re-endothelialisation after intravascular radiation.14
The underlying molecular mechanisms associated with GM-CSF-mediated effects on EPC function remain largely undefined, however. The current study was designed to investigate the effects of GM-CSF on EPC function and signal transduction through the establishment of a co-culture model with EPCs and cardiac microvascular endothelial cells (CMECs), which provides an in vitro, three-dimensional model for the assembly of sprouting endothelium and incorporated progenitor cells in vessel formation. The elucidation of the mechanisms underlying GM-CSF-mediated effects may provide valuable insights into EPC-mediated neovascularisation and new therapies for ischaemic diseases, such as atherosclerosis.
MATERIALS AND METHODS
Cell culture
All animal procedures were approved by the Care of Experimental Animals Committee of China Medical University. EPCs were isolated by density gradient centrifugation.15 Briefly, hollow leg bones of Sprague-Dawley (SD) rats (100 g - 120 g) were prepared by standard surgical procedures, and whole bone marrow was harvested by flushing the marrow with phosphate-buffered saline (PBS, pH 7.4) containing 20 u/mL heparin, using a syringe with a 20-gauge needle. The total mononuclear cell fraction was isolated by 1.083 g/L Ficoll-Paque (Becton Dickinson, San Jose, United States of America) density gradient centrifugation and washed three times with PBS. The cells were then plated at a density of 106 mononuclear cells/cm2 on culture plates (Becton Dickinson) containing endothelial cell basal medium-2 (EBM-2, Clonetics, San Diego, USA) supplemented with 10% foetal bovine serum (FBS), 5% fibroblast growth factor-2 (FGF-2), 5% epidermal growth factor (EGF), 5% insulin-like growth factor-1 (IGF-1), and 0.5 mmol/L ascorbic acid. Twenty-four hours later, nonadherent cells were aspirated and transferred to fresh fibronectin-coated plates to remove rapidly adherent haematopoietic cells and mature endothelial cells. After another 4 days in culture, nonadherent cells were removed by washing with PBS and the colony-forming units (CFUs) were determined. Rat CMECs were purchased from Cascade Biologics (Portland, USA).
Characterisation of EPCs
After 7 days in culture, the cells were stained with phycoerythrin-labelled anti-AC133 antibody (PE-AC133, Santa Cruz Biotechnology, Santa Cruz, USA). After 5 days, 7 days and 9 days in culture, AC133 and vascular endothelial growth factor receptor-2 (VEGFR-2) mRNA levels were also determined by semi-quantitative reverse transcriptase-polymerase chain reaction (RT-PCR); AC133 mRNA levels in CMECs cultured at the same time points were also determined by RT-PCR. After Trizol (Invitrogen, Carlsbad, USA) extraction, total RNA was reverse transcribed in 20-µL volumes using an RT-PCR kit (TaKaRa, Tokyo, Japan). For each RT product, aliquots (2 µL - 10 µL) of the final reaction volume were amplified in parallel PCR reactions using AC133-specific (315 bp product, sense 5'-GCAACAGGTACATCCACGAG-3', antisense 5'-TTTCATAAGGTTCAGAGCAA-3'), VEGFR-2-specific (250 bp product, sense 5'-GAGGCTCCGCTCCTTGTC-3', antisense 5'-AATGCTTTCTCCGCTTCGA-3'), and β-actin-specific (471 bp product, sense 5'-TGAAGTACCCCATTGAACACG-3', antisense 5'-CGAAGTCTAGGGCACATAGCA-3') primer sets. PCR cycles were as follows: 94 ºC for 40 s, 57 ºC for 40 s and
72 ºC for 60 s (35 cycles) for AC133 and VEGFR-2, and 94 ºC for 40 s, 53 ºC for 40 s and 72 ºC for 40 s (35 cycles) for β-actin, which served as an internal control. RT-PCR products were analysed by 2% agarose gel electrophoresis and were visualised with ethidium bromide under ultraviolet light.
In vitro co-culture model of neovascularisation
Growth-factor-reduced Matrigel (Becton Dickinson) was used for tubule-formation assays in vitro. Matrigel was thawed overnight at 4 ºC on ice and placed in a 24-well plate at room temperature for 30 min to allow solidification. EPCs were either pre-treated with 0 ng/mL (control), 10 ng/mL, 25 ng/mL or 50 ng/mL GM-CSF (Peprotech, London, United Kingdom) for 48 h, or with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h or 48 h. DAPI-labelled EPCs (105 cells) were co-plated with rat CMECs (4 × 105 cells) and incubated at 37 ºC. The number of EPCs contributing to in vitro tubule formation was assessed by counting the number of labelled cells in capillary-like structures that appeared in three random high-power fields for each group.
Proliferation and viability assay
After 7 days in culture, the EPCs were detached with 0.25% trypsin, harvested by centrifugation and counted. A total of 1 × 104 cells per well were cultured in 200 µL growth-factor-free medium in 96-well plates, and GM-CSF (0 ng/mL [control], 10 ng/mL, 25 ng/mL or 50 ng/mL) was added to each well for 48 h, or 50 ng/mL GM-CSF to each well for 12 h, 24 h, 36 h or 48 h; each group included five wells. EPC proliferation and viabilitywere determined by performing the method of transcription and translation (MTT) colorimetric assay. Briefly, the medium was removed and replaced with medium containing 5 mg/mL MTT (Sigma, St Louis, USA), followed by a 4-h incubation at 37 ºC. The supernatant was then aspirated and formazan crystal was dissolved in 150 mL dimethyl sulphoxide (DMSO, Sigma). The absorbance at 490 nm was measured for each well.
Migration assay
EPC migration was evaluated by using a Transwell chamber (6.5 mm, 8.0 µm pore size, Corning Life Sciences, New York, USA) assay. EPCs were harvested as described above. A total of 5 × 104 cells per well in 600 µL growth-factor-free medium were placed in the upper compartment of the chamber, and were then challenged by the addition of a chemoattractant solution (100 µL per well) composed of GM-CSF or no additional compounds (control) to the lower compartment. Each dose and time point was the same as described above. Non-transmigrated cells were scraped and the transmigrated cells were washed with PBS and fixed with 2% paraformaldehyde. For quantification, cells were stained with haematoxylin and counted manually in five random high-power fields per group.
Vasculogenesis assay
EPCs were harvested as described above and placed on top of Matrigel. A total of 105 cells per well of a 24-well plate were grown in the presence of GM-CSF or with no additional compound added (control). Each dose and time point was the same as described above. The number of tubules that formed was counted manually in three random high-power fields per group.
Immunoblot analysis of integrinβ2 and endogenous VEGF expression
EPCs were harvested after treatment with GM-CSF, as described above, and cellular proteins were isolated on ice in lysis buffer (20 mmol/L Tris-HCl, pH 8.0, 150 mmol/L NaCl, 1% Triton X-100, 2 mmol/L EDTA, 1 mmol/L phenylmethylsulfonylfluoride). Protein concentrations were determined using the bicinchoninic acid protein assay. An equal amount of protein (50 µg per lane) was separated by 12% SDS-polyacrylamide gel electrophoresis. Proteins from the analytical gel were blotted onto nitrocellulose membranes. Thereafter, the membranes were blocked in TBS-T buffer (5% non-fat milk in 25 mmol/L Tris-HCl, pH 8.0, 3 mmol/L KCl, 150 mmol/L NaCl, 0.05% Tween) and subsequently incubated with primary antibodies against rat integrinβ2 and VEGF (1:400, Santa Cruz Biotechnology) for 2 h at room temperature, followed by incubations with secondary horseradish peroxidase-conjugated antibodies for 1 h at room temperature. Beta-actin served as an internal loading control. The intensities of visualised signals were analysed densitometrically.16
Phospho-ERK1/2 expression
After 7 days in culture, EPCs were exposed to either no additional compound (control), GM-CSF (50 ng/mL), GM-CSF (50 ng/mL) plus the PI3K inhibitor LY294002 (50 µmol/L, Cell Signaling, Boston, USA), or GM-CSF (50 ng/mL) plus the PI3K inhibitor Wortmannin (50 µmol/L, Cell Signaling) for 15 min each. Immunoblotting was used to detect the expression of p-ERK1/2 (1:400, Santa Cruz Biotechnology), as described above.
Effects of ERK inhibitor
Experiments were carried out to determine whether ERK is required for GM-CSF-induced EPC function. After 7 days in culture, EPCs were exposed either to no additional compound (control), GM-CSF (50 ng/mL), or GM-CSF (50 ng/mL) plus the ERK inhibitor FR180204 (50 µmol/L, Calbiochem, San Diego, USA) for 48 h. EPC proliferation, viability, migration and vasculogenesis were then measured, as described above.
Statistical analyses
All data are presented as mean ± standard deviation (s.d.). Differences between group means were determined by one-way ANOVA followed by Student-Newman-Keuls test for post-hoc comparisons. Probability values of p < 0.05 were interpreted to denote statistical significance.
RESULTS
Characteristics of EPCs
Morphological characteristics of EPCs were observed under an inverted phase-contrast microscope. After 5 days in culture, EPCs formed island-like colony-forming units that consisted of a central core of round cells with elongated sprouting cells at the periphery (Figure 1A). AC133 is a novel haematopoietic stem cell marker that is expressed on EPCs, but is absent from mature endothelial cells. AC133-positive, VEGFR-2-positive cells are likely to correspond to EPCs. After 7 days in culture, EPCs stained positive for PE-AC133 (Figure 1B). When cultured for between 5 days and 9 days, EPCs maintained certain levels of AC133 and VEGFR-2 mRNA expression (Figure 1C). In contrast, AC133 mRNA expression was barely detectable in mature CMECs (Figure 1D).
GM-CSF improves EPC-mediated angiogenesis in vitro
The co-culture model of progenitor cells and endothelial cells mimics a three-dimensional assembly of sprouting endothelium and incorporated progenitor cells in vessel formation. In order to observe EPC-associated blood vessel formation in the current study, we developed a co-culture model using EPCs and CMECs to analyse the effect of GM-CSF on EPC-mediated angiogenesis in vitro. EPCs were pre-treated with GM-CSF and labelled with DAPI. After they were co-cultured with CMECs on Matrigel, the number of labelled EPCs (as determined by DAPI-stained nuclei) incorporated into capillary-like structures of CMECs was counted in three random high-power fields per group. The results showed that GM-CSF significantly increased the number of EPCs that were incorporated into capillary-like structures of CMECs in a dose- and time-dependent manner (Figures 2A and 2B). The dose-dependent results were as follows: 61.33 ± 4.16, 85.33 ± 6.43, 105.67 ± 6.03, and 145.33 ± 6.51 for EPCs treated with 0 ng/mL, 10 ng/mL, 25 ng/mL and 50 ng/mL GM-CSF for 48 h, respectively (n = 9 for each group, *p < 0.001 versus control [0 ng/mL]). The following were the time-dependent results: 64.33 ± 3.06, 82.67 ± 3.51, 111.67 ± 5.51, and 151.67 ± 6.51 for EPCs incubated with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h and 48 h, respectively (n = 9 for each group, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL] for 12 h).
GM-CSF promotes EPC function
Investigation as to whether GM-CSF improves EPC-mediated angiogenesis in vitro is connected with the effects of GM-CSF on EPC function. We assessed the influence of GM-CSF on EPC proliferation and viability, migration, and vasculogenesis using the MTT, Transwell chamber and Matrigel assays, respectively. The results showed that GM-CSF significantly promoted EPC proliferation and viability in a dose- and time-dependent manner (Tables 1A and 1B). Furthermore, GM-CSF promoted a dose- and time-dependent increase in EPC migration (Figure 3A and 3B) and vasculogenesis (Figures 4A and 4B); the number of EPCs that migrated through the chamber membrane and formed tubules increased significantly. The dose-dependent cell migration was as follows: 44.07 ± 4.98, 67.93 ± 1.03, 90.53 ± 2.17, and 150.67 ± 5.05 for EPCs grown next to lower chambers with 0 ng/mL, 10 ng/mL, 25 ng/mL and 50 ng/mL GM-CSF for 48 h, respectively (n = 15 for each group, *p < 0.001 versus control [0 ng/mL]). The time-dependent cell migration was the following: 51.67 ± 1.68, 80.40 ± 2.97, 103.67 ± 1.95, and 152.13 ± 5.99 for EPCs grown next to lower chambers with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h and 48 h, respectively (n = 15 for each group, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL] for 12 h). The dose-dependent tubule formation was: 3.33 ± 0.58, 8.33 ± 0.58, 23.00 ± 1.00, and 41.33 ± 1.16 for EPCs treated with 0 ng/mL, 10 ng/mL, 25 ng/mL and 50 ng/mL GM-CSF for 48 h, respectively (n = 9 for each group, *p < 0.001 versus control [0 ng/mL]). The time-dependent tubule formation was: 3.33 ± 0.58, 10.00 ± 1.00, 18.00 ± 1.00, and 41.33 ± 0.58 for EPCs incubated with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h and 48 h, respectively (n = 9 for each group, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL] for 12 h).
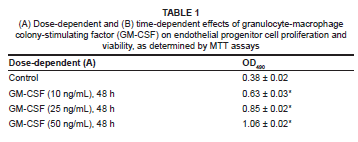

GM-CSF upregulates integrinβ2 and endogenous VEGF protein expression
Adhesion and paracrine signalling are both necessary for EPC-mediated angiogenesis. In the present study, we examined integrinβ2 and endogenous VEGF protein expression levels by immunoblot analysis. Figures 5A and 5B show that GM-CSF treatment led to a significant increase in the expression of integrinβ2 and endogenous VEGF in a dose- and time-dependent manner. The dose-dependent changes in integrinβ2 expression were: 0.31 ± 0.01, 0.41 ± 0.02, 0.49 ± 0.02, and 0.69 ± 0.02 for EPCs treated with 0 ng/mL, 10 ng/mL, 25 ng/mL and 50 ng/mL GM-CSF for 48 h, respectively (n = 3 for each group, p < 0.001 versus control [0 ng/mL]). The dose-dependent changes in VEGF expression were: 0.21 ± 0.01, 0.36 ± 0.02, 0.48 ± 0.02, and 0.60 ± 0.02 for EPCs treated with 0 ng/mL, 10 ng/mL, 25 ng/mL and 50 ng/mL GM-CSF for 48 h, respectively (n = 3 for each group, p < 0.001 versus control [0 ng/mL]). The time-dependent changes in Integrinβ2 expression were: 0.23 ± 0.01, 0.34 ± 0.01, 0.43 ± 0.02, and 0.58 ± 0.02 for EPCs treated with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h and 48 h, respectively (n = 3 for each group, p < 0.001 versus EPCs treated with GM-CSF [50 ng/mL] for 12h). The time-dependent changes in VEGF expression were: 0.03 ± 0.01, 0.31 ± 0.02, 0.39 ± 0.02, and 0.56 ± 0.01 for EPCs treated with 50 ng/mL GM-CSF for 12 h, 24 h, 36 h and 48 h, respectively (n = 3 for each group, p < 0.001 versus EPCs treated with GM-CSF [50 ng/mL] for 12 h).
GM-CSF activates ERK via PI3K pathway
To determine the possible molecular mechanisms underlying the effect of GM-CSF on EPC function, we determined the expression levels of p-ERK1/2 by immunoblot analysis. Figure 6 shows that GM-CSF treatment led to a significant increase in the expression of p-ERK1/2. The addition of two different PI3K inhibitors (LY294002 or Wortmannin) partially inhibited the GM-CSF-mediated increase in p-ERK1/2 expression. Therefore, GM-CSF might activate ERK via the PI3K pathway. The levels of p-ERK1/2 expression were: 0.25 ± 0.01, 0.58 ± 0.02*, 0.23 ± 0.01Δ and 0.28 ± 0.01Δ for control, GM-CSF, GM-CSF plus LY294002, and GM-CSF plus Wortmannin (n = 3 for each group, *p < 0.001 versus control, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL]).
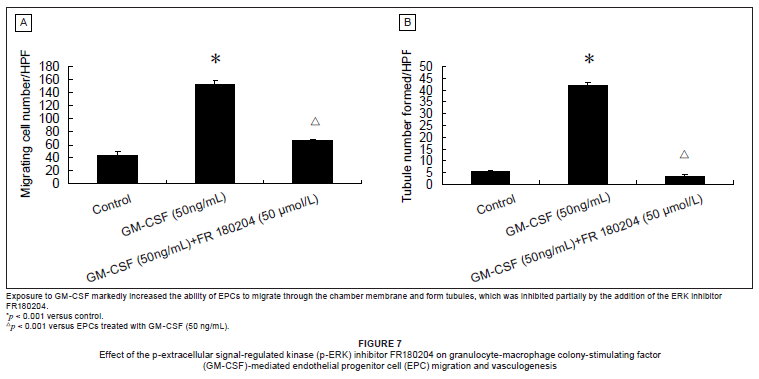
The effect of an ERK inhibitor on GM-CSF-mediated activation of EPC function
To determine the effect of the ERK inhibitor FR180204 on GM-CSF-mediated activation of EPC function, we assayed EPC proliferation and viability, migration, and vasculogenesis using the MTT, Transwell chamber and Matrigel assays, respectively. Table 2 and Figures 7A and 7B show that GM-CSF significantly increased the proliferation and viability of EPCs, the number of EPCs migrating through the membrane and EPC tubule formation. The addition of the ERK inhibitor FR180204 led to a marked decrease in GM-CSF-mediated EPC proliferation, viability, migration and tubule formation. Collectively, the results of these experiments suggest that GM-CSF modulates EPC function via a PI3K-dependent ERK signalling pathway. The average number of migrating cells per high-power field were: 44.33 ± 5.51, 152.47 ± 6.76, and 66.20 ± 1.52 for control, GM-CSF, and GM-CSF plus FR180204, respectively (n = 15 for each group, *p < 0.001 versus control, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL]). The average number of tubules formed included 5.33 ± 0.58, 42.00 ± 1.00, and 3.33 ± 0.58 for control, GM-CSF, and GM-CSF plus FR180204, respectively (n = 9 for each analysis, *p < 0.001 versus control, Δp < 0.001 versus EPCs treated with GM-CSF [50 ng/mL]).

DISCUSSION
In the current study, we successfully established a co-culture system using EPCs and CMECs to analyse the effects of GM-CSF on EPC-mediated angiogenesis in vitro. Our results have shown that GM-CSF treatment leads to a dose- and time-dependent increase in the number of EPCs incorporated into capillary-like structures of CMECs. Based on these findings, we conclude that GM-CSF enhances EPC-mediated angiogenesis in vitro.
It is conceivable that a complex pathway may recruit EPCs to sites of ischaemia and incorporate them into newly formed blood vessels. This would involve a coordinated sequence of events, including proliferation, migration, adhesion, paracrine signalling and, finally, the differentiation of EPCs into endothelial cells. To determine whether GM-CSF-regulated EPC angiogenesis may be linked to such a coordinated sequence of events, we assessed the effect of GM-CSF on additional EPC functions; collectively, the results of the current study reveal that GM-CSF exerts several beneficial effects on EPCs.
Although originally considered a key regulator of haematopoiesis, the major role of GM-CSF may lie in its ability to induce a functional angiogenic programme in cultured endothelial cells. In the current study, we found that EPCs incubated with GM-CSF exhibited increased proliferation, viability, migration and vasculogenesis. Previous studies have shown that GM-CSF triggers vessel sprouting and angiogenesis17,18 and that treatment with a GM-CSF antagonist can lead to a marked reduction in neovascularisation.19 Furthermore, GM-CSF was recently shown to induce the proliferation and rate of S-phase entry of EPCs from murine bone marrow mononuclear cells in vitro.20
Cell adhesion is involved in the homing of stem cells to tissues with active angiogenesis. In the current study, we examined integrinβ2 expression, which is required for EPC adhesion to endothelial cells.21 Ex vivo-expanded EPCs express integrinβ2, whichhas been shown markedly to enhance both the incorporation of transplanted EPCsinto endothelial cell monolayers in vitro and the neovascularisation of ischaemic limbs in vivo.22 Previous studies demonstrated that several cell adhesion molecules, including intercellular adhesion molecule-1 (ICAM-1) and CD44, were upregulated by GM-CSF.23 The results of the current study showed that GM-CSF significantly upregulated integrinβ2 expression in EPCs in a dose- and time-dependent manner. Further studies are needed to elucidate the potential role of other adhesion molecules during the recruitment of EPCs to ischaemic tissue.
The efficiency of neovascularisation may be attributable not only to the incorporation of EPCs into newly formed vessels, but also to the release of pro-angiogenic factors by EPCs in a paracrine manner.24,25 Early outgrowth of EPCs (after 4-7 days in culture) is associated with the secretion of additional pro-angiogenic cytokines, including VEGF.26 Previous studies have shown that inflammatory cell-derived VEGF may act as an angiogenic mediator of the healing effect of GM-CSF in chronic ulcers.27 The results of the current study demonstrated that GM-CSF significantly upregulates endogenous VEGF expression in EPCs in a dose- and time-dependent manner, which, in turn, may influence the classical process of EPC-mediated angiogenesis. Additional studies will be required to determine the extent of EPC incorporation, paracrine effects and other possible effects.
The current study included experiments to determine the possible mechanisms underlying the effects of GM-CSF on EPC function. GM-CSF is known to activate several classical cellular signalling pathways, including the PI3K/Akt, JAK/STAT and Ras-Raf-MEK-ERK kinase cascades, in both haematopoietic and endothelial cells. A previous study in neutrophils showed that GM-CSF-dependent ERK phosphorylation was decreased by PI3K inhibitors, whereas ERK inhibitors did not affect GM-CSF-induced PI3K phosphorylation,28 suggesting that PI3K acts upstream of ERK. The current study extended these findings by showing that the PI3K inhibitors LY294002 and Wortmannin partially inhibited GM-CSF-induced upregulation of p-ERK1/2 in EPCs. These data suggested that GM-CSF may activate ERK via PI3K in EPCs. Moreover, the results of the current study show that FR180204, a selective and cell-permeable inhibitor of ERK, inhibits the ability of GM-CSF to induce EPC proliferation, migration, and vasculogenesis. Taken together, these results suggest that PI3K-dependent ERK activation is required for GM-CSF-mediated effects on EPC function and that GM-CSF may regulate EPC function via a PI3K-dependent ERK signalling pathway.
CONCLUSION
The haematopoietic and inflammatory factor GM-CSF exerts several beneficial effects on EPC function, including proliferation, migration, adhesion, paracrine signalling, and differentiation. This coordinated set of events is predicted to be involved in the recruitment of EPCs to sites of ischaemia and in the incorporation of EPCs into newly formed vessels. These events may occur partly via a PI3K-dependent ERK signal transduction pathway. Taken together, the results of the current study suggest that EPCs may serve as a useful therapeutic tool for increasing neovascularisation, rescuing ischaemic tissue and improving cardiovascular disease prognosis.
REFERENCES
1. Asahara T, Masuda H, Takahashi T, et al. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ Res. 1999;85(3):221-228. [ Links ]
2. Asahara T, Murohara T, Sullivan A, et al. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275(5302):964-967. [ Links ]
3. Grant MB, May WS, Caballero S, et al. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med. 2002;8(6):607-612. [ Links ]
4. Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for organ vascularization and regeneration. Nat Med. 2003;9(6):702-712. [ Links ]
5. Aicher A, Heeschen C, Mildner-Rihm C, et al. Essential role of endothelial nitric oxide synthase for mobilization of stem and progenitor cells. Nat Med. 2003;9(11):1370-1376. [ Links ]
6. Murayama T, Tepper OM, Silver M, et al. Determination of bone marrow-derived endothelial progenitor cell significance in angiogenic growth factor-induced neovascularization in vivo. Exp Hematol. 2002;30(8):967-972. [ Links ]
7. Kalka C, Masuda H, Takahashi T, et al. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc Natl Acad Sci USA. 2000;97(7):3422-3427. [ Links ]
8. Kocher AA, Schuster MD, Szabolcs MJ, et al. Neovascularization of ischaemic myocardium by human bone-marrow-derived angioblasts prevents cardiomyocyte apoptosis, reduces remodeling and improves cardiac function. Nat Med. 2001;7(4):430-436. [ Links ]
9. Kawamoto A, Gwon HC, Iwaguro H, et al. Therapeutic potential of ex vivo expanded endothelial progenitor cells for myocardial ischaemia. Circulation. 2001;103(5):634-637. [ Links ]
10. Assmus B, Schächinger V, Teupe C, et al. Transplantation of progenitor cells and regeneration enhancement in acute myocardial infarction (TOPCARE-AMI). Circulation. 2002;106(24):3009-3017. [ Links ]
11. Tateishi-Yuyama E, Matsubara H, Murohara T, et al. Therapeutic angiogenesis for patients with limb ischaemia by autologous transplantation of bone-marrow cells: A pilot study and a randomised controlled trial. Lancet. 2002;360(9331):427-435. [ Links ]
12. Takahashi T, Kalka C, Masuda H, et al. Ischaemia and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nat Med. 1999;5(4):434-438. [ Links ]
13. Seiler C, Pohl T, Wustmann K, et al. Promotion of collateral growth by granulocyte-macrophage colony-stimulating factor in patients with coronary artery disease: A randomized, double-blind, placebo-controlled study. Circulation. 2001;104(17):2012-2017. [ Links ]
14. Cho HJ, Kim HS, Lee MM, et al. Mobilized endothelial progenitor cells by granulocyte-macrophage colony-stimulating factor accelerate reendothelialization and reduce vascular inflammation after intravascular radiation. Circulation. 2003;108(23):2918-2925. [ Links ]
15. Thum T, Fraccarollo D, Galuppo P, et al. Bone marrow molecular alterations after myocardial infarction: Impact on endothelial progenitor cells. Cardiovasc Res. 2006;70(1):50-60. [ Links ]
16. Scheubel RJ, Kahrstedt S, Weber H, et al. Depression of progenitor cell function by advanced glycation endproducts (AGEs): Potential relevance for impaired angiogenesis in advanced age and diabetes. Exp Gerontol. 2006;41(5):540-548. [ Links ]
17. Valdembri D, Serini G, Vacca A, et al. In vivo activation of JAK2/STAT-3 pathway during angiogenesis induced by GM-CSF. FASEB J. 2002;16(2):225-227. [ Links ]
18. Mann A, Breuhahn K, Schirmacher P, et al. Keratinocyte-derived granulocyte-macrophage colony stimulating factor accelerates wound healing: Stimulation of keratinocyte proliferation, granulation tissue formation, and vascularization. J Invest Dermatol. 2001;117(6):1382-1390. [ Links ]
19. Mann A, Niekisch K, Schirmacher P, et al. Granulocyte-macrophage colony-stimulating factor is essential for normal wound healing. J Investig Dermatol Symp Proc. 2006;11(1):87-92. [ Links ]
20. Wang QR, Wang F, Zhu WB, et al. GM-CSF accelerates proliferation of endothelial progenitor cells from murine bone marrow mononuclear cells in vitro.Cytokine. 2009;45(3):174-178. [ Links ]
21. Wu Y, Ip JE, Huang J, et al. Essential role of ICAM-1/CD18 in mediating EPC recruitment, angiogenesis, and repair to the infarcted myocardium. Circ Res. 2006;99(3):315-322. [ Links ]
22. Chavakis E, Aicher A, Heeschen C, et al. Role of beta2-integrins for homing and neovascularization capacity of endothelial progenitor cells. J Exp Med. 2005;201(1):63-72. [ Links ]
23. Ip WK, Wong CK, Wang CB, et al. Interleukin-3, -5, and granulocyte macrophage colony-stimulating factor induce adhesion and chemotaxis of human eosinophils via p38 mitogen-activated protein kinase and nuclear factor kappaB. Immunopharmacol Immunotoxicol. 2005;27(3):371-393. [ Links ]
24. Urbich C, Dimmeler S. Endothelial progenitor cells: Characterization and role in vascular biology. Circ Res. 2004;95(4):343-353. [ Links ]
25. Rehman J, Li J, Orschell CM, et al. Peripheral blood 'endothelial progenitor cells' are derived from monocyte/macrophages and secrete angiogenic growth factors. Circulation. 2003;107(8):1164-1169. [ Links ]
26. Hur J, Yoon CH, Kim HS, et al. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 2004;24(2):288-293. [ Links ]
27. Cianfarani F, Tommasi R, Failla CM, et al. Granulocyte/macrophage colony-stimulating factor treatment of human chronic ulcers promotes angiogenesis associated with de novo vascular endothelial growth factor transcription in the ulcer bed. Br J Dermatol. 2006;154(1):34-41. [ Links ]
28. Tortorella C, Simone O, Piazzolla G, et al. Role of phosphoinositide 3-kinase and extracellular signal-regulated kinase pathways in granulocyte macrophage-colony-stimulating factor failure to delay fas-induced neutrophil apoptosis in elderly humans. J Gerontol A Biol Sci Med Sci. 2006;61(11):1111-1118. [ Links ]
 Correspondence to:
Correspondence to:
Xiang-Hong Yang
Postal address:
Department of Pathology, Affiliated Shengjing Hospital, China Medical University
Sanhao Street 36, Heping District, Shenyang
Liaoning 110004, China
email: xhyang4933@vip.sina.com.cn
Received: 8 Jan. 2010
Accepted: 12 Apr. 2010
Published: 10 June 2010
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}
{kind=link}