










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.106 n.3-4 Pretoria Mar./Apr. 2010
RESEARCH ARTICLE
Characterisation of South African isolates of Fusarium oxysporum f.sp. cubense from Cavendish bananas
Marinda VisserI; Tom GordonII; Gerda FourieI; Altus ViljoenI,III
IDepartment of Microbiology and Plant Pathology, Forestry and Agricultural Biotechnology Institute, University of Pretoria, South Africa
IIDepartment of Plant Pathology, University of California, Davis CA 95616, USA
IIIDepartment of Plant Pathology, Stellenbosch University, South Africa
ABSTRACT
Fusarium wilt, caused bythe soil-borne fungusFusarium oxysporum f.sp. cubense (Foc), is a serious vascular disease of bananas in most subtropical and tropical regions of the world. Twenty-four vegetative compatibility groups (VCGs) and three pathogenic races have been identified in Foc, reflecting a relatively high genetic diversity for an asexual fungus. To characterise a South African population of Foc, a collection of 128 isolates from diverse geographic origins were isolated from diseased Cavendish bananas and subjected to VCG analysis and sequencing of the translation elongation factor 1-α (TEF) gene region. The presence of mating type genes was also determined using MAT-1 and MAT-2 specific primers. VCG 0120 was established as the only VCG of Foc present in the South African population studied. Only the MAT-2 idiomorph was present in all the local isolates of Foc. A phylogenetic analysis of DNA sequences of the TEF gene region revealed that the South African isolates grouped closely with VCG 0120 isolates from Australia and Asia. These results suggest that the South African population of Foc was most likely introduced in a limited number of events and that it had spread with infected planting material within the country. The presence of only one mating type and the limited diversity in this pathogen render it unlikely to rapidly overcome disease management strategies involving host resistance.
Keywords: bananas; Fusarium wilt; mating types; phylogenetics; vegetative compatibility groups
INTRODUCTION
Fusarium wilt of bananas is caused by the fungus Fusarium oxysporum f.sp. cubense (E.F. Smith) Snyder and Hansen (Foc), a pathogen that is generally considered to be one of the most destructive formae speciales of F.oxysporum.1,2 The disease seriously hampers banana production once Foc is introduced into fields and it is difficult to manage. Because the pathogen persists in infested soils for long periods,3 control strategies involve the use of tissue culture-derived plantlets to prevent the introduction of Foc into disease-free fields, as well as the implementation of sanitation practices to prevent spread.4,5 The most effective means of controlling Fusarium wilt, however, is the replacement of susceptible banana cultivars with resistant ones.
Fusarium wilt became notorious because it almost destroyed the Gros Michel-based banana export industry in Central America during the mid-1900s. The disease was eventually managed by replacing Gros Michel bananas with highly resistant Cavendish cultivars.1 Since then, Cavendish bananas have been found to succumb to a new race of Foc, called Foc race 4, in other banana-producing areas of the world.6 Cavendish cultivars are the only banana varieties produced commercially in South Africa. Since the 1970s, Fusarium wilt has destroyed almost 40% of all Cavendish bananas grown in the Kiepersol and southern KwaZulu-Natal areas of the country.5 The disease has also been discovered in two further production areas since the turn of the century.7 New replacement cultivars for Cavendish bananas have not been readily accepted by producers and markets, primarily because of a slight difference in taste. Owing to the parthenocarpic nature of Cavendish bananas, unconventional improvement, rather than classical breeding, now offers the most feasible option to develop Fusarium-wilt-resistant Cavendish banana cultivars.
Knowledge of the genetic diversity of fungal populations and their mode of reproduction is important for implementing management strategies to reduce disease impact.8,9 Foc has a relatively diverse population structure for an apparently asexual fungus that consists of three races3,4,10 and 24 vegetative compatibility groups (VCGs).6,11,12 A teleomorph for F. oxysporum has never been observed and the pathogen appears to rely on mutations and parasexuality as the main basis for genetic variation.13,14,15,16 Polymerase chain reaction (PCR) amplification experiments have demonstrated the presence of both MAT idiomorphs in at least two formae speciales of F. oxysporum,17,18,19 but MAT idiomorphs in Foc have not yet been reported.
Studies to determine diversity in Foc have included both phenotypic and genotypic markers. The phenotypic characters most commonly used are pathogenic race and vegetative compatibility.20,21,22 Foc races 1, 2 and 4 are distinguished from one another based on their virulence to a defined group of banana cultivars under field conditions.3,4,23 Foc race 1 attacks Gros Michel, Silk, Apple, Lady Finger and Latundan cultivars, while race 2 attacks Bluggoe bananas and race 4 attacks all cultivars susceptible to Foc races 1 and 2, as well as Cavendish bananas. Although predisposing factors, such as cold temperatures, are associated with damage caused by Foc race 4 in the subtropics,5 these factors are not involved in Fusarium wilt of Cavendish bananas in the tropics. To recognise the effect of environmental factors and differences that exist between populations of Foc causing disease to Cavendish bananas in the subtropics and tropics, the pathogens are referred to as 'subtropical' and 'tropical' race 4, respectively.
Vegetative compatibility is a useful means of subdividing Foc into genetically isolated groups, but does not measure genetic relatedness among isolates. In addition, VCGs are phenotypic markers that may be subjected to selection pressures.24,25 Therefore, neutral DNA-based techniques would be more suitable for analysing genetic variation within and between Foc populations. DNA sequence analyses of several formae speciales of F.oxysporum, including isolates of Foc, have shown that the Fusarium wilt fungus represents two genetically distinct lineages.26 Concordant evidence from the gene genealogies further revealed that Foc harbours at least five clonal lineages.26
Foc is believed to have spread worldwide through infected planting material.1,2,10 The route of entry of the pathogen into South Africa is unknown due to incomplete records of banana production in the country. It is thought that Indian labourers, who worked on sugar cane plantations in KwaZulu-Natal during colonial times, could have introduced infected rhizomes into South Africa.27 In this study, the identity of Foc VCGs in a collection of isolates from South Africa was determined. The phylogenetic relationship of the South African isolates in relation to those from other banana-producing countries was assessed by DNA sequence data for the translation elongation factor 1-α (TEF) gene regions. We further considered whether both mating type genes were present in the South African Foc population to determine if sexual reproduction might occur in this fungus.
METHODS
Fungal isolates
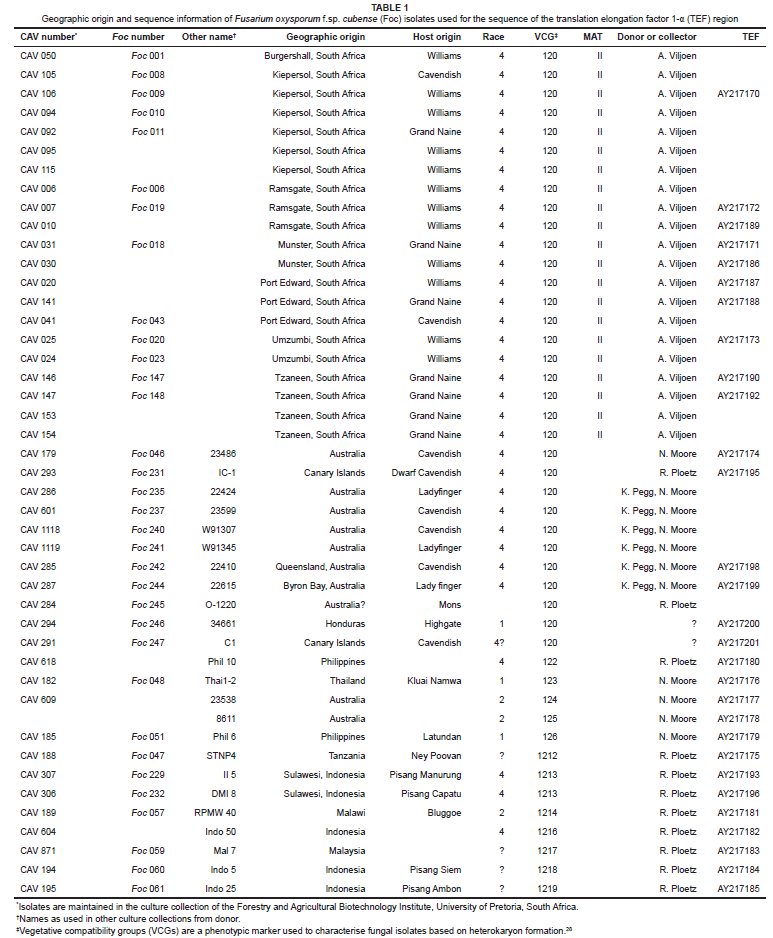
Foc isolates (n = 152) from different banana genotypes and geographic origins were selected for this study. These included 128 isolates collected from Kiepersol and Komatipoort (Mpumalanga Province), Tzaneen (Limpopo Province) and the south-coast region of the KwaZulu-Natal Province in South Africa, and 24 VCG tester isolates from the collections of Dr N. Moore of the Queensland Department of Primary Industries (Australia), Dr S. Bentley of the University of Queensland (Australia) and Dr R. Ploetz of the University of Florida (USA). The VCG tester isolates represented Foc races 1 and 2, as well as Foc 'tropical' and 'subtropical' race 4. All cultures were single-spored and were maintained in the culture collection of the Forestry and Agricultural Biotechnology Institute, University of Pretoria, South Africa.
Generation of nitrate non-utilising mutants and VCG testing
Vegetative compatibility of the 128 South African Foc isolates was determined using the technique described by Puhalla.28 In this technique, isolates are assigned to VCGs based on heterokaryon formation between complementary nitrate non-utilising (nit) mutants produced on media supplemented with chlorate. Nit mutants were produced for all South African isolates as well as for the known VCG tester strains. The nit-1 and nit-3 mutants were then paired with each of the nit-M tester strains on minimal medium (MM) at least twice.29 Nit-M, nit-3 and nit-1 mutants of the same isolate were also paired to test for self-compatibility. Complementary nit mutants that formed dense, wild-type growth on MM were assigned to the same VCG. Vegetatively incompatible isolates were detected by their inability to form a heterokaryon when paired on MM.
DNA extraction
After VCG identification, 45 Foc isolates were selected for sequence analysis and mating type identification (Table 1). These isolates consisted of 21 Foc isolates representative of the different geographic areas, the Cavendish cultivars grown in South Africa and the 24 VCG tester isolates. For mating type identification, only the 21 South African Foc isolates were considered. All 45 Foc isolates were grown in 100 mL potato dextrose broth (Biolab Diagnostics, Wadeville, South Africa) in 250-mL flasks, without shaking, at room temperature for 7-10 days, after which the mycelium was harvested and freeze dried. For extraction of total DNA, freeze-dried mycelia were ground to a fine powder in liquid nitrogen and added to DNA extraction buffer (200 mM Tris-HCl, pH 8; 150 mM NaCl; 25 mM EDTA, pH 8; 0.5% SDS),30 followed by phenol-chloroform extraction.31 The DNA concentration was estimated by comparing the intensity of ethidium bromide fluorescence of the DNA samples to known concentrations of lambda DNA marker (marker III) (Roche Molecular Biochemicals, Mannheim, Germany) following gel electrophoresis.
Identification of mating type genes
To determine if MAT-1 or MAT-2 idiomorphs were present in the South African population of Foc, DNA of the 21 representative Foc isolates were subjected to PCR analysis with primers designed by Steenkamp et al.32 Additional primers were designed for MAT-1 (FO-MAT-1-For 5'ACC GCC AGC CGT CGT GCA GTG 3'and FO-MAT-1-Rev 5'CTT GCG GGG GTA TGA GAA CGC 3') based on the MAT-1idiomorph sequences in GenBank, while a MAT-2 reverse primer was designed specifically for the high mobility group (HMG) box (FF1 Foc 5' GTA TCT TCT GTC CAC CAC AG 3') and used with the forward primer Gfmat2c.32
For each isolate, a 25-µL PCR reaction mix was prepared that contained 0.4 mM of each deoxynucleoside triphosphate (dNTPs), 1 x PCR buffer, 1.0 pmol of each primer, 0.25 units Expand High Fidelity Taq polymerase (Roche Molecular Biochemicals, Germany), 2 ng DNA, and sterile deionised water. PCR reactions were performed on a Hybaid TouchDown PCR machine (Hybaid Limited, Middlesex, United Kingdom) and reaction conditions were as follows: initial denaturation at 95 ºC for 2 min, followed by 35 cycles of denaturation at 92 ºC for 30 s, primer annealing at 62 ºC (MAT 1) or 54 ºC (MAT 2) for 40 s, elongation at 72 ºC for 2 min, and a final extension at 72 ºC for 7 min. The amplified product was resolved by electrophoresis in a 1.5% (w/v) agarose gel in TBE buffer (Tris boric acid EDTA; pH 8.0), stained with ethidium bromide and visualised under UV illumination.31 Size estimates of the PCR fragments were determined using a molecular weight standard (100-bp ladder; Promega, Madison, Wisconsin, USA).
DNA sequence analyses
A standard 25-µL PCR reaction mixture for TEF was prepared as described above and reaction conditions set as follows: denaturation at 95 ºC for 2 min, followed by 30 cycles of 30 s at 95 ºC, 40 s at 60 ºC,26 1 min at 72 ºC, and a final extension of 7 min at 72 ºC. The amplified products were verified using electrophoresis in 1.5% agarose gels in Tris borate EDTA (TBE, pH 8.0) buffer. The resulting PCR amplicons were purified using a QIAquick PCR Purification kit (QIAGEN, Straβe, Germany), according to the specifications of the manufacturer.
Purified PCR products were sequenced in both directions using the PCR primers listed above. DNA sequences were determined using the ABI PRISMTM Dye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems, Foster City, California, USA) and an ABI PRISMTM 377 automated sequencer. For comparative purposes, five TEF sequences,26 which represent the two clades and clonal lineages, were obtained from GenBank and included in the analyses. NRRL 22903, previously used by O'Donnell et al.26 as an out group, was also included in the current study as an out group. All sequences from the current study were deposited in GenBank (http://www.ncbi.nlm.nih.gov/) (Table1).
Datasets were aligned with MAFFT (http://mafft.cbrc.jp/alignment/software/) software.33 Bayesian inference was accomplished using MrBayes version 3.b.4 34 and maximum likelihood methods were employed by using PhyML version 2.4.3 35 Bootstrap confidence levels were assessed by 1000 parsimony replications.
RESULTS
Generation of nit mutants and VCG testing
Nit mutants were successfully generated for all the South African isolates and the known VCG testers of Foc. Almost all isolates produced at least one nit-M mutant and several nit-1 and/or nit-3 mutants. Pairings of nit-M mutants with nit-1 or nit-3 mutants of the same isolate produced a zone of wild-type growth where the two nit-mutants formed a heterokaryon. When paired with the different VCG tester isolates, nit-1 mutants from the South African Foc population formed heterokaryons only with the nit-M tester isolate of VCG 0120 (Table 1). No complementary reactions resulted from pairings of the South African Foc isolates with testers representing any other VCG.
Identification of mating type genes
Amplification of genomic DNA with MAT-2 specific primers (Gfmat2c and FF1 Foc; Gfmat2c and Gfmat2d) produced an amplicon for all the South African Foc isolates tested (Table 1). A PCR reaction with the primer pair Gfmat2c and FF1 amplified a 700-bp fragment and the primer pair Gfmat2c and Gfmat2d resulted in a 200-bp PCR product. PCR using the MAT-1 primers with genomic DNA as a template consistently failed to produce an amplicon.
DNA sequence analysis
Amplification of the TEF region yielded a fragment of 700 bp. Alignment of the DNA sequences resulted in a data set of 598 characters. When aligned by TEF sequences, Foc isolates in the current study were broadly separated into two clades (Clades A and B; Figure 1). The South African population of Foc VCG 0120 'subtropical' race 4 grouped within Clade A with Foc isolates representing VCGs 0122, 0126, 0120/01215, 01212, 01213, 01216 and 01218. This grouping, as well as the relationships within it, however, was not supported by high bootstrap values. Isolates from the Indo-Malaysian region representing VCGs 01213 and 01216 appeared to form a subclade within Clade A. Clade B included isolates representing VCGs 0123, 0124, 0125, 01214, 01217 and 01219.
DISCUSSION
All isolates of Foc from commercial banana plantations in South Africa tested in this study belong to VCG 0120. This VCG is best known for its ability to cause disease of Cavendish bananas following incidents of environmental stress2,5 and has been reported from many banana-producing areas worldwide.2,6 The occurrence of a single VCG in South Africa indicates that the genetic diversity within the South African population is low and reconfirms the idea that the Fusarium wilt fungus was introduced into the country, most likely in a single or only a few events.
The phylogenetic tree generated in this study shows that the South African population of Foc harbours isolates that are closely related to Foc isolates from Australia, the Canary Islands and Central America. It seems likely, therefore, that VCG 0120 was originally introduced into subtropical and tropical Cavendish-producing areas - such as South Africa, Australia, the Canary Islands and Central America - with infected banana planting material from Southeast Asia.21,22 Planting material has also been moved between southern KwaZulu-Natal and Mpumalanga, which could have contributed to the movement of the Fusarium wilt pathogen between production areas.
Isolates of Foc globally are clearly heterogeneous. Sequencing results of this study showed that Foc can be divided into two phylogenetic clades with potentially separate evolutionary origins and five genetically distinct clonal lineages, as described by Koenig et al.36 and O'Donnell et al.26 Clade A can be divided into at least two lineages and Clade B into three lineages. The first clonal lineage in Clade A consisted almost entirely of isolates representing VCG 0120, while the second clonal lineage included isolates representing Foc 'tropical' race 4 (VCGs 1213 and 1216). Clade B consisted of three clonal lineages, all made up of isolates belonging to Foc races 1 and 2. Since the different pathogenic lineages may be capable of causing disease to different host genotypes,37 banana improvement programmes must consider different pathogen lineages when developing plants with Fusarium wilt resistance. The race structure in Foc is not well defined and the genotypic groups defined above can, in future, be used to redefine pathotypes in Foc.
The occurrence of only the MAT-2 idiomorph in a representative population of isolates of Foc from South Africa provides strong evidence that sexual reproduction is absent in this fungus in the country. This finding is of great importance to the development of future management strategies for Fusarium wilt of bananas, since phytopathogenic fungi with an ability to reproduce sexually may overcome disease resistance in plants more rapidly than asexual forms. This has been true in bananas where the sexually reproducing fungus responsible for black Sigatoka, Mycosphaerella fijiensis Morelet, rapidly developed resistance to fungicides.38 Both mating types have previously been reported for F. oxysporum17,18 but, to date, no studies have revealed clear evidence of sexual reproduction in contemporary populations.19 The stability of resistance to Fusarium wilt in Cavendish bananas in Central America may be further testimony to the absence of sexual reproduction and the consequent inability of the pathogen to generate new pathotypes.
ACKNOWLEDGEMENTS
We acknowledge financial support from the National Research Foundation, South Africa, the THRIP initiative of the South African Department of Trade and Industry, the US Department of Agriculture, the University of Pretoria and the Banana Growers Association of South Africa. We also thank Dr Natalie Moore of Queensland, Australia and Prof Randy Ploetz of the University of Florida, USA, for supplying Foc cultures, as well as Sharon Kirkpatrick of the University of California, Davis CA, USA, for useful advice regarding pairings made between isolates.
REFERENCES
1. Stover RH. Fusarium wilt (Panama disease) of banana and other Musa species. Surrey: Commonwealth Mycological Institute; 1962. [ Links ]
2. Ploetz RC. Population biology of Fusarium oxysporum f.sp. cubense. In: Ploetz RC, editor. Fusarium wilt of banana. St Paul: APS Press, 1990; p. 63-67. [ Links ]
3. Stover RH, Buddenhagen IW. Banana breeding: Polyploidy, disease resistance and productivity. Fruits. 1986;41:175-191. [ Links ]
4. Stover RH, Simmonds NW. Bananas. 3rd ed. London: Longmans, 1987; p. 366-378. [ Links ]
5. Viljoen A. The status of Fusarium wilt (Panama disease) of banana in South Africa. S Afr J Sci. 2002;98:341-344. [ Links ]
6. Ploetz RC, Pegg KG. Fungal disease of the root, corm, and pseudostem: Fusarium wilt. In: Jones DR, editor. Diseases of banana, abaca and enset. London: CAB International, 2000; p. 143-157. [ Links ]
7. Grimbeek EJ, Viljoen A, Bentley S. First occurrence of Panama disease in two banana- growing areas of South Africa. Plant Dis. 2001;85:1211. [ Links ]
8. McDonald BA, McDermott JM. Population genetics of plant pathogenic fungi. Bioscience. 1993;43:311-319. [ Links ]
9. Taylor JW, Geiser DM, Burt A, Koufopanou V.The evolutionary biology and population genetics underlying strain typing. Clin Microbiol Rev.1999;12:126-146. [ Links ]
10. Pegg KG, Langdon PW. Fusarium wilt (Panama disease). In: Persley GJ, DeLanghe EA, editors. Banana and plantain breeding strategies, ACIAR Proc. No. 21. Cairns: ACIAR, 1987; p. 119-123. [ Links ]
11. Katan T. Current status of vegetative compatibility groups in Fusarium oxysporum. Phytoparasitica. 1999;27:51-64. [ Links ]
12. Katan T, Di Primo P. Current status of vegetative compatibility groups in Fusarium oxysporum supplement. Phytoparasitica. 1999;27:1-5. [ Links ]
13. Buxton EW. Heterokaryosis and recombination in pathogenic strain of Fusarium oxysporum. J Gen Microbiol.1956;15:133-139. [ Links ]
14. Buxton EW. Parasexual recombination in the banana-wilt Fusarium. Trans Br Mycol Soc. 1962;45:274-279. [ Links ]
15. Parameter JR, Snyder WC, Reichle RE. Heterokaryosis and variability in plant-pathogenic fungi. Annu Rev Phytopathol.1963;1:51-76. [ Links ]
16. Tinline RD, Macneil BH.Parasexuality in plant pathogenic fungi. Annu Rev Phytopathol.1969;7:147-166. [ Links ]
17. Yoshida T, Arie T, Nomura Y, Yoder OC, Turgeon BG, Yamaguchi I.Cloning mating type genes of the asexual ascomycete Fusarium oxysporum. Poster presented at: The VIIth International Congress on Plant Pathology; 1998 Aug 9-15; Edinburgh, United Kingdom. [ Links ]
18. Kerényi Z, Zeller KA, Hornok L, Leslie JF. Molecular standardization of mating type terminology in the Gibberella fujikuroi species complex. Appl Environ Microbiol. 1999;65:4071-4076. [ Links ]
19. Arie T, Kaneko I, Yoshida T, Noguchi M, Nomura Y, Yamaguchi I. Mating-type genes from asexual phytopathogenic ascomycete, Fusarium oxysporum and Alternaria alternata. Mol Plant Microbe Interact. 2000;13:1330-1339. [ Links ]
20. Ploetz RC, Correll JC. Vegetative compatibility among races of Fusarium oxysporum f.sp. cubense. Plant Dis. 1988;72:325-328. [ Links ]
21. Brake VM, Pegg KG, Irwin JAG, Langdon PW. Vegetative compatibility groups within Australian populations of Fusarium oxysporum f.sp. cubense, the cause of Fusarium wilt of bananas. Aust J Agric Res. 1990;41:863-870. [ Links ]
22. Moore NY, Pegg KG, Allen RN, Irwin JAG. Vegetative compatibility and distribution of Fusarium oxysporum f.sp. cubense in Australia. Aust J Exp Agric. 1993;33:797-802. [ Links ]
23. Stover RH. Fusarium wilt of banana: Some history and current status of the disease. In: Ploetz RC, editor. Fusarium wilt of banana. St Paul: APS Press, 1990; p. 1-7. [ Links ]
24. Glass NL, Kuldau GA. Mating type and vegetative incompatibility in filamentous ascomycete. Annu Rev Phytopathol. 1992;30:201-224. [ Links ]
25. Leslie JF. Vegetative compatibility in fungi. Annu Rev Phytopathol. 1993;31:127-151. [ Links ]
26. O'Donnell K, Kistler HC, Cigelnik E, Ploetz RC. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc Natl Acad Sci USA. 1998;95:2044-2049. [ Links ]
27. Ploetz RC, Herbert J, Sebasigari K, et al. Importance of Fusarium wilt in different banana- growing regions. In: Ploetz RC, editor. Fusarium wilt of banana. St Paul: APS Press, 1990; p. 9-26. [ Links ]
28. Puhalla JE. Classification of strains of Fusarium oxysporum on the basis of vegetative compatibility. Can J Bot. 1985;63:179-183. [ Links ]
29. Correll JC, Klittich CJR, Leslie JF. Nitrate nonutilizing mutants of Fusarium oxysporum and their use in vegetative compatibility tests. Phytopathology. 1987;77:1640-1646. [ Links ]
30. Raeder U, Broda P. Rapid preparation of DNA from filamentous fungi. Lett Appl Microbiol. 1985;1:17-20. [ Links ]
31. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: A laboratory manual. 2nd ed. New York: Cold Spring Harbour Press, 1989; p. E3-E4. [ Links ]
32. Steenkamp ET, Wingfield BD, Coutinho TA, et al. PCR-based identification of MAT-1 and MAT-2 in the Gibberella fujikuroi species complex. Appl Environ Microbiol. 2000;66:4378-4382. [ Links ]
33. Kazutaka K, Kazuharu M, Kei-ichi K, Takashi M. MAFFT: A novel method for rapid multiple sequence alignment based on fast fourier transform. Nucleic Acid Res. 2002;30:3059-3066. [ Links ]
34. Heulsenbeck JP, Ronquist F, Nielsen R, Bollack JP. Bayesian inference of phylogeny and its impact on evolutionary biology. Science. 2001;294:2310-2314. [ Links ]
35. Guidon S, Gascuel O. A simple, fast and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol. 2003;52:696-704. [ Links ]
36. Koenig R, Ploetz RC, Kistler HC. Fusarium oxysporum f.sp. cubense consists of a small number of divergent and globally distributed lineages. Phytopathology. 1997;87:915-923. [ Links ]
37. Woudt LP, Neuvel A, Sikkema A, de Milliano WAJ, Campbell CL, Leslie JF. Fusarium from cyclamen. Phytopathology. 1995;85:1348-1355. [ Links ]
38. Carlier J, Fouré E, Gauhl F, et al. Black leaf streak. In: Jones DR, editor. Diseases of banana, abaca and enset. Wallingford: CABI Publishing, 2000; p. 37-79. [ Links ]
 Correspondence to:
Correspondence to:
Altus Viljoen
Postal address:
Department of Plant Pathology, Stellenbosch University
Private Bag X1, Matieland 7601, South Africa
email: altus@sun.ac.za
Received: 9 Apr. 2009
Accepted: 23 Nov. 2009
Published: [To be released]
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}