










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.106 no.1-2 Pretoria Jan./Fev. 2010
RESEARCH LETTER
Characterisation of the arsenic resistance genes in Bacillus sp. UWC isolated from maturing fly ash acid mine drainage neutralised solids
Wicleffe Musingarimi; Marla Tuffin; Donald Cowan
Institute for Microbial Biotechnology and Metagenomics, University of the Western Cape, Cape Town, South Africa
ABSTRACT
An arsenic resistant Bacillus sp. UWC was isolated from fly ash acid mine drainage (FA-AMD) neutralised solids. A genomic library was prepared and screened in an arsenic sensitive mutant Escherichia coli strain for the presence of arsenic resistance (ars) genes. Sequence analysis of a clone conferring resistance to both sodium arsenite and sodium arsenate revealed homologues to the arsR (regulatory repressor), arsB (membrane located arsenite pump), arsC (arsenate reductase), arsD (second regulatory repressor and a metallochaperone) and arsA (ATPase) genes from known arsenic resistance operons. The Bacillus sp. UWC arsRBCDA genes were shown to be arranged in an unusual manner with the arsDA genes immediately downstream of arsC.
Keywords: fly ash; acid mine drainage; arsenic resistance genes; Bacillus; groundwater contamination
INTRODUCTION
Coal mining in South Africa is the second biggest mining sector after gold, with 16% export revenue and 4% of the gross domestic product. Acid mine drainage (AMD) and fly ash (FA) are produced through coal mining and, through the subsequent use of coal in power generation, contribute towards environmental pollution. AMD (pH 2.3 - pH 6.5) is formed in coal mining by the oxidation of metal sulphides found in the coal, rock and/or clay that is on top or below coal seams and dissolves heavy metals and metalloids (including arsenic) which pollute environmental waters. FA (pH 12) is a silicaceous particulate matter typically disposed of in surface 'ash dumps'. It consists mainly of silicon, aluminium, iron and calcium oxides. Arsenic can also be associated with FA, predominantly as arsenate,1 usually complexed with carbonates or iron-manganese oxides.2 As with AMD, natural hydrological processes can mobilise potentially toxic elements in ash dumps into groundwater. There is current interest in utilising FA for the neutralisation of AMD, thereby potentially simultaneously resolving two environmental problems.3 The 'neutralisation solids' generated from the co-disposal process are generally rich in insoluble metals and metalloids (including potentially toxic elements such as arsenic, lead and chromium). Such solids have been considered for use in soil remediation, for back-filling and for other municipal and agricultural purposes.4,5
Current research in our laboratory has shown that 'neutralisation solids', which may be sterile at the point of preparation, develop microbial communities during exposure and maturation (Mutondo and Cowan, unpublished results). These communities may include microbial species whose activities could result, potentially, in environmental hazards. These transformations include methanogenesis, sulphate reduction (sulphidogenesis) or metal/metalloid redox processes, resulting in mobilisation or enhanced toxicity of metals and metalloids.
Arsenic is the most environmentally abundant toxic metal and all living organisms are exposed to low levels of arsenic naturally occurring in both water and soil.6 In soils, arsenic is present in two oxidation states: As(III) (arsenite) and As(V) (arsenate).7 Under oxidising conditions arsenate predominates and strongly adsorbs onto or forms minerals with iron, manganese and aluminium, thus reducing its solubility.8 Generally, As(V) is the major species in arsenic-contaminated soils.9 Under reducing or anaerobic conditions, the more toxic and soluble form of arsenic, arsenite, occurs. Due to the abundance of arsenic in the environment, microorganisms are under competitive pressure to maintain arsenic detoxification systems. These microbial activities are also responsible for arsenic cycling. It is possible that arsenic present in both AMD and FA could be mobilised into environments where neutralisation solids have been applied, increasing the risk of groundwater contamination.
The most common form of arsenic resistance in prokaryotes is through detoxification operons encoded on plasmids and/or genomes. These operons are diverse, but most often contain three genes: arsR (transcriptional repressor), arsC (arsenate reductase) and arsB (arsenite efflux membrane protein).10 Some have two additional genes, arsD (second regulator and a metallochaperone) and arsA (ATPase), which confer higher resistance than the three-gene operon.11 Arsenate entering the cell is reduced to the more toxic arsenite by ArsC, and is then transported out of the cell via ArsB (complexed with ArsA if it is present).10 In this way, prokaryotes contribute to arsenite cycling in the environment.
In this study, we describe the isolation of an arsenic-resistant Bacillus strain from matured FA-AMD neutralised solids, together with the identification, cloning and characterisation of an ars operon.
MATERIALS AND METHODS
Bacterial strains, plasmids and media
Escherichia coli (E. coli) and Bacillus sp. UWC were grown at 37 ºC in Luria-Bertani (LB) broth or agar medium,12 with appropriate supplements of either ampicillin (100 µg/mL) or chloramphenicol (30 µg/mL) as required.
Isolation and identification of arsenic-resistant organisms
FA-AMD neutralised solids were obtained from Eskom3 and dried. The solids were transferred into a pot (with a height of 25 cm, diameter of 20 cm, and a volume of 7 L) and mixed with moist garden soil (50% w/w) and exposed to the environment for six weeks prior to sampling. Samples (5 g) were suspended in 10 mL sterile water containing 2% (v/v) Tween 20,13 then decanted into 2 mL sterile tubes before centrifugation at 5000 rpm for 90 s. Aliquots (200 µL) of the supernatant were aseptically spread in duplicate onto LB agar supplemented with 160 mM sodium arsenate, and incubated aerobically at 37 ºC for 3-7 days. Colony morphology was observed under a stereomicroscope and Gram staining was carried out according to the method described by Cowan and Steel14. For identification of the arsenic-resistant isolate described in this study, a single colony was suspended in 50 µL water and treated for 5 min at 100 ºC. This was used as a template for amplification using the universal bacterial 16S rRNA gene primers, E9F 9 and U1510R 15. The reactions were carried out in an Applied Biosystems thermocycler in a 50 µL volume containing: 1x PCR Buffer; 100 ng DNA template; 0.5 µM of each primer; 200 µM of each dNTP and 1.25 U Taq DNA polymerase. The cycling conditions were: initial denaturation at 94 ºC for 4 min; followed by 30 cycles of denaturation (94 ºC for 30 s), annealing (52 ºC for 30 s), elongation (72 ºC for 2 min); and a final elongation step at 72 ºC for 10 min. A partial (first 700 bp) 16S rRNA sequence was determined directly from the amplicon.
DNA isolation and manipulations
For the construction of the Bacillus sp. UWC genomic library in E. coli DH10B (Invitrogen), DNA was isolated from cells in late-stationary growth phase using the cetyltrimethylammonium bromide (CTAB) method12. Partially MboI-digested DNA was fractionated by electrophoresis on an agarose gel and those between 5 kb and 10 kb were ligated into the BglII restriction site of plasmid pEcoR252.16 For routine cloning and manipulation procedures, standard procedures were used.12 DNA fragments were purified from agarose gels using the PCR and Gel Band Purification kit (GE Healthcare, Cape Town, South Africa). Plasmid DNA was extracted using the Invisorb Spin Plasmid Mini Kit (Invitek, Berlin, Germany).
Screening for arsenic resistance genes
The Bacillus sp. UWC genomic library was screened in the ars deletion mutant E. coli ACSH50Iq17 and clones were selected for their ability to complement the mutant on LB plates containing 1 mM sodium arsenate or 0.5 mM sodium arsenite. Plasmid DNA was extracted from positive clones and re-transformed to confirm arsenic resistance. Clones were analysed by restriction enzyme digestion and subcloned for sequencing.
Arsenic resistance assays
To test for the level of resistance conferred by the Bacillus sp. UWC ars genes, assays in E. coli ACSH50Iq containing plasmids pArs8, pUM3 (as positive control)18 and pEcoBlunt (as negative control)19 were carried out in LB media containing chloramphenicol (30 µg/mL) and increasing concentrations of sodium arsenite or sodium arsenate. Overnight cultures were diluted 100-fold into fresh media and incubated at 37 ºC for 5 h and absorbance was determined at 600 nm. The incubation period corresponded to the middle of the log phase of growth of a control culture grown under the same conditions. The resistance was expressed as a percentage of the optical density (OD600) compared with the control culture with no added arsenic.
Sequencing analysis
Sequencing was by the dideoxy chain-termination method using an ABI 3130XL Genetic Analyser. Sequences were analysed using a variety of software program, including the DNAMAN package from Lynnon BioSoft. Comparison searches were performed using the gapped-BLAST program.20 The Bacillus sp. UWC 16S rRNA and arsRBCDA gene cluster sequences have been deposited in the GenBank database with the following accession numbers: FJ554669 and FJ542199, respectively.
RESULTS
Isolation of an arsenic-resistant organism from maturing FA-AMD neutralisation solids
Several bacterial strains with different colony morphologies were isolated from FA-AMD samples on media containing 160 mM sodium arsenate. One of these, visualised as a Gram-positive rod, was analysed further. The 16S rRNA gene was amplified and partial sequencing (700 bp) indicated that it belonged to the genus Bacillus (accession number FJ554669), and was named Bacillus sp. UWC. This isolate was also able to grow in media containing 0.5 mM sodium arsenite.
Isolation of arsenic resistance genes
To identify and isolate ars genes, a genomic library of the Bacillus sp. UWC isolate was prepared and screened in the E. coli arsenic-sensitive mutant ACSH50Iq. Approximately 6 000 transformed clones were plated on media containing 0.5 mM sodium arsenite, and 10 of these (pArs1-10) were arsenite resistant. The plasmids were analysed by restriction digestion (data not shown) and pArs8 was selected for further analysis. In order to identify the arsenic resistance genes in pArs8, several subclones were prepared and the plasmid inserts sequenced.
Sequence analysis of pArs8
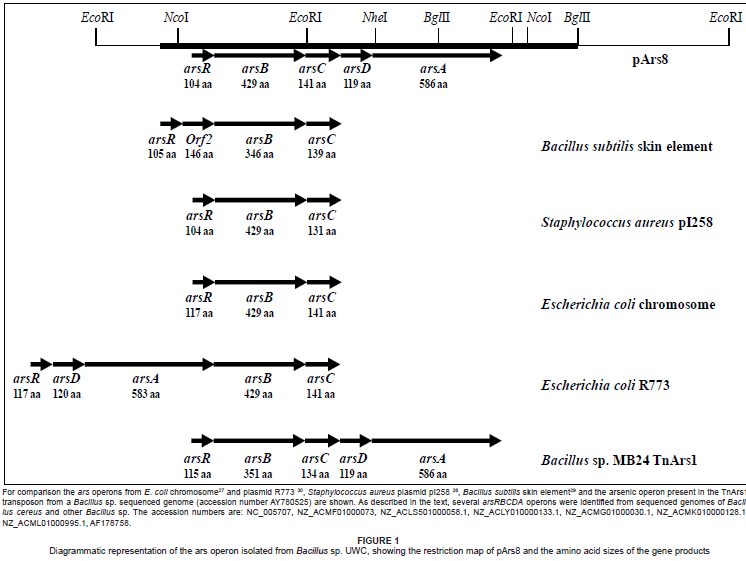
The insert of plasmid pArs8 contained five open reading frames (ORFs) with high amino acid identity to genes associated with arsenic resistance genes in Gram-positive organisms (Table 1). The ORFs were encoded for ArsR (negative regulator)21, ArsB (membrane pump for arsenite efflux),22,23 ArsC (arsenate reductase),24 ArsD (a second repressor and chaperone to deliver As(III) to ArsA)25,26 and ArsA (an ATPase that associates with ArsB and links arsenite export to ATP hydrolysis)22. The highest sequence identities obtained were with Bacillus sp genes. BLAST analyses of the Bacillus sp. UWC ars gene products are shown in Table 1. The ars operon genes were positioned in the order arsRBCDA and transcribed in the same direction. While the arsRBC genes were ordered, as is commonly found in typical operons from the E. coli chromosome,27 Staphylococcus aureus plasmid pI258 29 and the Bacillus skin element29 (Figure 1), the arsD and arsA genes were identified together, unusually situated downstream of the arsRBC genes. The arsDA genes are always associated and their linkage suggests that they have dependent functions in arsenic detoxification.25 Operons containing arsDA confer high level of resistance to arsenic and have been described in E. coli plasmid R773,30 Acidithiobacillus caldus transposon TnAtcArs 20 and Leptospirillum ferriphilum transposon TnLfArs.31 In none of these operons were the arsDA genes directly downstream of the arsRBC genes. A BLAST search using the Bacillus sp. UWC ars gene sequences identified the presence of a similar arsRBCDA operon in several sequenced genomes from Bacillus cereus strains and other Bacillus species (Figure 1), suggesting a common organisation in this genus. However, the individual ars genes from Bacillus sp. UWC did not necessarily share the highest sequence identity with those identified from the sequenced genomes (Table 1). This suggests the possibility of a common ancestor operon, but with substantial evolutionary changes in the individual ars genes. The placement of the arsDA genes directly downstream of the arsRBC genes in this genus makes it tempting to speculate that the ancestor contained the typical arsRBC operon and later acquired the arsDA genes. In one B. cereus strain (ATCC 10987), the ars genes were identified on a plasmid (accession number NC_005707), while in the Bacillus sp. MB24 they occurred on a transposon. Typically, ars operons containing arsDA genes are found on plasmids and transposons19,30,31 and are therefore easily transferred by horizontal gene transfer.31 However, since total genomic DNA was used to prepare the genebank from which the Bacillus sp. UWC ars genes were isolated, it is not possible to determine their location without further experimentation.
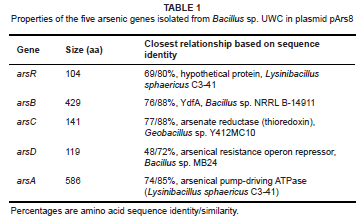
Analysis of the arsRBCDA gene products revealed the presence of motifs typical for these proteins. ArsR contained the typical ELCVCDL metal-binding6,32 and the helix-turn-helix DNA binding21,33 domains. Bacterial ArsCs have been divided into two families: those requiring thioredoxin34,35 and those requiring glutathione/glutaredoxin.36 Alignment of the Bacillus sp. UWC ArsC with ArsC proteins from both groups suggests that it groups with the Trx clade and is therefore likely to be a thioredoxin type ArsC. Furthermore, it has the four conserved cysteine residues (Cys10, 15, 82, 89, numbering for the pI258 ArsC) identified as being required for activity in this family.34,35 ArsD was originally reported to be a second, weak transcriptional regulator,26 but has recently been shown to function as a chaperone, transferring arsenite to the ArsA for export by the ArsB and thereby conferring higher resistance.25 The E. coli R773 plasmid ArsD has three vicinal cysteine pairs (Cys12-13, Cys112-113, and Cys119-120) and an additional cysteine, Cys18.17,26 It has been proposed that only Cys12-13 and Cys18 are required for the metallochaperone activity,37 while the Cys112-113 pair is associated with the weak transcriptional activity.38 The Bacillus sp. UWC ArsD only contains Cys12-13 and Cys18, which might suggest that it functions only as the metallochaperone. The Bacillus sp. UWC ArsA contained the nucleotide binding domain and the conserved 12 residue motif (DTAPTGHTIRLL), called the DTAP motif,39 in both the A1 and A2 halves. The three cysteine residues required for As(III)/Sb(III) binding and activation40 are conserved in the Bacillus sp. UWC ArsA (Cys113, 172 and 422), suggesting that it too is involved in binding arsenite and possibly antimony.
Arsenic resistance in E. coli
When expressed in the E. coli ars mutant ACSH50Iq, construct pArs8 conferred an almost equal level of resistance to arsenate but a lower level of resistance to arsenite, compared to the positive control pUM3 (Figure 2). Although the experiment was not done in a low phosphate medium (phosphate competes with arsenate uptake thereby reducing the 'toxicity' of arsenate ions),41 the comparative data with the arsenic-sensitive E. coli strain ACSH50Iq is strongly suggestive that the ars genes in pArs8 confer in vivo arsenic resistance.
DISCUSSION
To date, many arsenic-resistant microorganisms have been isolated from various environmental sources. Resistant isolates are found in both arsenic-uncontaminated1 and arsenic-contaminated environmental samples.42 Here, we report the isolation of a Bacillus isolated from FA-AMD neutralised solids which showed resistance to 160 mM sodium arsenate. Given the six week exposure of the FA-AMD sample to the environment, the chances of isolating common aerobic soil organisms were high43 and several Bacillus species have been isolated from heavy metal-contaminated environments.43,44,45
General mechanisms of toxic metal resistance have been well studied and documented.6,46 The most common resistance mechanism is the efflux of arsenite from the cell, encoded by either a three- or five-gene operon. The Bacillus sp. UWC isolate contains a five-gene operon, arsRBCDA, exhibiting a unique arrangement. While mRNA expression studies of the genes were not performed, the resistance conferred in E. coli, the presence of arsDA genes and the conserved active site amino acids identified in all the genes, together suggest that this operon is involved in resistance to high levels of arsenic in this Bacillus isolate.
These findings have some relevance to the possible in situ function of the Bacillus sp. UWC isolate (and other arsenic resistant organisms) in the FA-AMD neutralised solids. Although other factors such as pH and redox potential would also have an influence, the natural functioning of this operon may have toxigenic capacity: reducing low toxicity As(V) (arsenate) to high toxicity As(III) (arsenite) and exporting the reduced metalloid to pore water where it could, potentially, be leached to groundwater. To further evaluate this, analysis of the distribution and frequency of occurrence of homologous ars resistance genes in the FA-AMD neutralised solids metagenome should be assessed. This should be coordinated with an analysis of microbial function (demonstrating the capacity to reduce As(V) in situ) and chemical determination of arsenic speciation.
ACKNOWLEDGEMENTS
The authors thank D. Surender and K. Reynolds of Eskom (SA) for provision of the FA-AMD neutralised solids and with the development of the FA-AMD project; and L. Petrik, Department of Chemistry, University of the Western Cape, for assistance with chemical analyses. This work was funded by The Water Research Commission, South Africa.
REFERENCES
1. Huggins FE, Senior CL, Chu P, Ladwig K, Huffman GP. Selenium and arsenic speciation fly ash from full-scale coal-burning utility plants. Environ Sci Technol. 2007;41:3284-3289. [ Links ]
2. Narukawa T, Takatsu A, Chiba K, Riley KW, French DH. Investigation on chemical species of arsenic, selenium and antimony in fly ash from coal fuel thermal power stations. J Environ Monit. 2005;7:1342-1348. [ Links ]
3. Gitari MW, Petrik LF, Etchebers O, Key DL, Iwuoha E, Okujeni C. Treatment of acid mine drainage with fly ash: removal of major contaminants and trace elements. J Environ Sci Heal A. 2006;41:1729-1747. [ Links ]
4. Benzaazoua M, Belem T, Bussiere B. Chemical factors that influence the performance of mine sulphidic paste backfill. Cement Concrete Res. 2002;32:1133-1144. [ Links ]
5. Gitari WM, Petrik LF, Etchebers O, Key DL, Iwuoha E, Okujeni C. Passive neutralisation of acid mine drainage by fly ash and its derivatives: A column leaching study. Fuel. 2008;87:1637-1650. [ Links ]
6. Silver S, Pung LT. Genes and enzymes involved in bacterial oxidation and reduction of inorganic Arsenic. Appl Environ Microb. 2005;71:599-608. [ Links ]
7. Cullen WR, Reimer KJ. Arsenic speciation in the environment. Chem Rev. 1989;89:713-764. [ Links ]
8. Mok MW, Wa CM. Mobilization of arsenic in contaminated river waters. In: Nriagu JO, editor. Arsenic in the environment. Part I: cycling and characterization. New York: Willy-Interscience, 1994; p. 99-117. [ Links ]
9. Garcia-Manyes S, Jiménez G, Padró A, Rubio R, Rauret G. Arsenic speciation in contaminated soils. Talanta. 2002;58:97-109. [ Links ]
10. Rosen BP. Families of arsenic transporters. Trends Microbiol. 1999;7:207-212. [ Links ]
11. Chen CM, Msira TK, Silver S, Rosen BP. Nucleotide sequence of the structural genes for an anion pump. The plasmid-encoded arsenical resistance operon. J Biol Chem 1986;261:15030-15038. [ Links ]
12. Sambrook J, Fritsch EF, Manniatis T. Molecular cloning: a laboratory manual. 2nd ed. Cold Spring Harbour: Cold Spring Harbour Laboratory Press; 1989. [ Links ]
13. Williams WL, Broquist HP and Snell EE. Oleic acid and related compounds as growth factors for lactic acid bacteria. J Biol Chem 1947;170:619-630. [ Links ]
14. Cowan ST, Steel KJ. Manual for the identification of medical bacteria. 3rd ed. Barrow GI, Feltham RKA, editors. Cambridge: Cambridge University Press, 1993. [ Links ]
15. Reysenbach AL, Pace NR. Reliable amplification of hyperthermophilic archaeal 16S rRNA genes by Polymerase Chain Reaction. In: Rob FT, Place AR, editors. Archaea: a laboratory manual - thermophiles. Cold spring Harbour: Cold Spring Harbour Laboratory Press, 1995; p. 101-107. [ Links ]
16. Zabeau M, Stanley KK. Enhanced expression of cro-beta-galactosidase fusion proteins under the control of the PR promoter of bacteriophage lambda. EMBO J. 1982;1:1217-1224. [ Links ]
17. Butcher BG, Rawlings DE. The divergent chromosomal ars operon of Acidithiobacillus ferrooxidans is regulated by an atypical ArsR protein. Microbiol. 2002;148:3983-3992. [ Links ]
18. Mobley HL, Chen CM, Silver S, Rosen BP. Cloning and expression of R-factor mediated arsenate resistance in Escherichia coli. Mol Gen Genet 183;191:421-426. [ Links ]
19. Tuffin IM, de Groot P, Deane SM, Rawlings DE. An unusual Tn21-like transposon containing an ars operon is present in highly arsenic-resistant strains of the biomining bacterium Acidithiobacillus caldus. Microbiol. 2005;151:3027-3039. [ Links ]
20. Altschul SF, Madden TL, Schäffer AA, et al. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389-3402. [ Links ]
21. Wu J, Rosen BP. The ArsR protein is a trans-acting regulatory protein. Mol Microbiol. 1991;5(6):1331-1336. [ Links ]
22. Tisa LS,Rosen BP. Molecular characterisation of an anion pump. The ArsB protein is the membrane anchor for the ArsA protein. J Biol Chem 1990;265:190-194. [ Links ]
23. Wu J., Tisa LS, Rosen BP. Membrane topology of the ArsB protein, the membrane subunit of an anion-translocating ATPase. J Biol Chem 21992;67:12570-12576. [ Links ]
24. Ji G, Silver S. Reduction of arsenate to arsenite by the ArsC protein of the arsenic resistance operon of Staphylococcus aureus plasmid pI258. Proc Natl Acad Sci. Sci U S A. 1992;89: 7974-7978. [ Links ]
25. Lin Y, Walmsley AR, Rosen BP. An arsenic metallochaperone for an arsenic detoxification pump. P Natl Acad Sci Sci U S A. 2006;103:15617-15622. [ Links ]
26. Wu J, Rosen BP. The arsD gene encodes a second trans-acting regulatory protein of the plasmid-encoded arsenical resistance operon. Mol Microbiol. 1993;8:615-623. [ Links ]
27. Carlin A, Shi W, Dey S, Rosen BP. The ars operon of Escherichia coli confers arsenical and antimonial resistance. J Bacteriol. 1995;177(4):981-986. [ Links ]
28. Ji G, Silver S. Regulation and expression of the arsenic resistance operon from Staphylococcus aureus plasmid pI258. J Bacteriol. 1992;174(11):3684-3694. [ Links ]
29. Sato T, Kobayashi Y. The ars operon in the skin element of Bacillus subtilis confers resistance to arsenate and arsenite. J Bacteriol. 1998;180(7):1655-1661. [ Links ]
30. Hedges RW, Baumberg SA. Resistance to arsenic compounds conferred by a plasmid transmissible between strains of Escherichia coli. J Bacteriol. 1973;115(1):459-460. [ Links ]
31. Tuffin IM, Hector SB, Deane SM, Rawlings DE. Resistance determinants of a highly arsenic-resistant strain of Leptospirillum ferriphilum isolated from a commercial biooxidation tank. Appl Environ Microb. 2006;72(3):2247-2253. [ Links ]
32. Shi W, Wu J, Rosen BP. Identification of a putative metal binding site in a new family of metalloregulatory proteins. J Biol Chem. 1994;269:19826-19829. [ Links ]
33. Wu J, Rosen BP. Metalloregulated expression of the ars operon. J Biol Chem. 1993;268(1):52-58. [ Links ]
34. Bennett MS, Guan Z, Laurberg M, Su XD. Bacillus subtilis arsenate reductase is structurally and functionally similar to low molecular weight protein tyrosine phosphatases. P Natl Acad Sci U S A. 2001;98:13577-138582. [ Links ]
35. Messens J, Hayburn G, Desmyter A, Laus G, Wyns L. The essential catalytic redox couple in arsenate reductase from Staphylococcus aureus. Biochemistry. 1999;38:16857-16865. [ Links ]
36. Martin P, DeMel S, Shi J, et al. Insights into the structure, solvation and mechanism of ArsC arsenate reductase, a novel arsenic detoxification enzyme. Structure. 2001;9:1071-1081. [ Links ]
37. Lin Y, Yang J, Rosen BP. ArsD residues Cys12 , Cys13, and Cys18 form an As(III)-binding site required for arsenic metallochaperone activity. J Biol Chem. 2007;282:16783-16791. [ Links ]
38. Li S, Chen Y, Rosen BP. Role of vicinal cysteine pairs in metalloid sensing by the ArsD As(III)-responsive repressor. Mol Microbiol. 2001;41:687-696. [ Links ]
39. Zhou T, Radaev S, Rosen BP, Gatti DL. Structure of the ArsA ATPase: the catalytic subunit of a heavy metal resistance pump. EMBO J. 2000;19:4838-4845. [ Links ]
40. Bhattacharjee H, Li J, Ksenzonko MY, Rosen BP. Role of cysteinyl reidues in metalloactivation of the oxyanion-translocating ArsA ATPase. J Biol Chem. 1995;270:11245-11250. [ Links ]
41. Oden KL, Gladysheva TB, Rosen BP. Arsenate reduction mediated by the plasmid-encoded ArsC protein is coupled to glutathione. Mol Microbiol. 1994;12:301-306. [ Links ]
42. Chopra BK, Bhat S, Mikheenko IP, et al. The characteristics of rhizosphere microbes associated with plants in arsenic-contaminated soils from cattle dip sites. Sci Total Environ. 2007;378:331-342. [ Links ]
43. Achour AR, Bauda P, Billard P. Diversity of arsenite transporter genes from arsenic-resistant soil bacteria. Res Microbiol. 2007;158:128-137. [ Links ]
44. Anderson CR, Cook GM. Isolation and characterization of arsenate-reducing bacteria from arsenic-contaminated sites in New Zealand. Curr Microbiol. 2004;48:341-347. [ Links ]
45. Joshi DN, Florab SJS, Kaliaa K. Bacillus sp. strain DJ-1, potent arsenic hypertolerant bacterium isolated from the industrial effluent of India. J Hazard Mater. 2009;166(2-3):1500-1505. [ Links ]
46. Rosen BP. Transport and detoxification systems for transition metals, heavy metals and metalloids in eukaryotic and prokaryotic microbes. Comp Biochem Physiol A Mol Integr Physiol. 2002;133(3):689-93. [ Links ]
 Correspondence to:
Correspondence to:
Marla Tuffin
Institute for Microbial Biotechnology and Metagenomics
University of the Western Cape, Private Bag X17
Bellville 7535, Cape Town, South Africa
email: ituffin@uwc.ac.za
Received: 19 Jan. 2009
Accepted: 07 Dec. 2009
Published: 11 Feb. 2010
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}