










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.106 no.1-2 Pretoria ene./feb. 2010
RESEARCH LETTER
One or two species? A morphometric comparison between robust australopithecines from Kromdraai and Swartkrans
Zachary CofranI; J. Francis ThackerayII
IDepartment of Anthropology, University of Michigan, United States of America
IIInstitute for Human Evolution, University of the Witwatersrand, Johannesburg, South Africa
ABSTRACT
The type specimen of Paranthropus robustus (TM 1517, including a partial cranium) was discovered at Kromdraai near Sterkfontein in 1938 and described by Robert Broom as a new species. Subsequently, more robust australopithecines were discovered at the nearby site of Swartkrans. These Swartkrans hominins were described by Broom as Paranthropus crassidens. Many palaeoanthropologists currently regard the robust australopithecines from Kromdraai and Swartkrans as one species, but consensus has not been reached on this issue. A morphometric analysis has been undertaken to assess the probability that specimens attributed to P. crassidens represent the same species as that which is represented by TM 1517, the holotype of P. robustus. Our results failed to reject the null hypothesis that both sites sample the same, single species of robust australopithecine.
Keywords: Kromdraai; Swartkrans; Paranthropus robustus; Paranthropus crassidens; STET
INTRODUCTION
The type specimen of Paranthropus robustus (TM 1517, see Figure 1) was discovered at Kromdraai B in the Cradle of Humankind World Heritage Site in South Africa and described by Broom1 as a new genus and species. Subsequently, additional specimens of 'robust' australopithecines were discovered at Swartkrans, within four kilometers of Kromdraai B, and described as a distinct species, P. crassidens.2 Many palaeoanthropologists such as Tobias3, Brain4, Fuller5, and Kaszycka6, regard the Kromdraai B and Swartkrans 'robust' australopithecine specimens as representing a single species. Others such as Grine7,8,9,10 and Howell11, and more recently, Schwartz and Tattersall12, regard the two as distinct at a species level, P. robustus and P. crassidens, respectively. To date, this issue has not been addressed by a morphometric analysis of both cranial and dental material. The objective of this study is to test the hypothesis that the robust australopithecine crania from Swartkrans and Kromdraai are morphometrically similar enough to belong to the same species, P. robustus.

MATERIALS
Craniodental measurements were taken on original specimens: TM 1517a from Kromdraai, and SK 12, SK 46, SK 48, SK 79, SK 83, and SKW 11 from Swartkrans. The set of cranial measurements was adapted from that of Lockwood13, and is listed in Table 1. Dental measurements included only the maximum labiolingual/buccolingual breadth, because heavy interproximal wear obscures maximum mesiodistal lengths in most specimens.5,14 Where a trait was present on both the right and left sides, the average of the two values was taken.
METHODS
Thackeray and colleagues15,16 described a method whereby measurements of pairs of specimens were compared to one another, in order to assess probabilities of conspecificity. Measurements obtained from extant, conspecific male-female pairs were plotted against each other, with the female on the x-axis and male on the y-axis. Morphometrically similar pairs tended to exhibit a relatively low degree of scatter about a least-squares regression line associated with the equation y = mx + c. This degree of scatter, or degree of dissimilarity, was quantified by calculating the standard error of the slope, m (here, "sem"). Using modern conspecifics as a frame of reference, Thackeray and colleagues15 found that this sem statistic displayed a normal distribution when log-transformed (base 10). Thus, pairs of specimens that were morphometrically very different from one another - specimens of different species - can be expected to have a relatively high degree of scatter about the regression line, and so a relatively high sem. Conversely, pairs of conspecifics tended to have a lower degree of scatter and hence a relatively low sem. Using an extant reference sample of over 1 400 specimens of vertebrates and invertebrates, Thackeray16 presented a log sem value of -1.61 ± 0.23 as a statistical definition of a species expressed in terms of probabilities.
The log sem method described by Thackeray and colleagues15 was developed further by Wolpoff and Lee17,18, who used a statistic which they referred to as the 'standard error test of the null hypothesis' (STET). Like log sem, STET is based on the standard error of the slope of the least-squares regression line in bispecimen comparisons. However, Wolpoff and Lee18 argued against the dampening effects of logging the sem statistic. Additionally, they noted that regressing specimen x on specimen y produced different slopes and therefore different standard errors, as compared to regressions of y on x. Thus, STET utilises the standard error of the slopes of both possible pair-wise regressions: STET = [(s.e.mx)2 + (s.e.my)2]1/2, where mx and my are the least-squares slopes of each possible regression. In this way, the STET statistic is invariant of how specimens are regressed on one another, and makes no assumptions about sex, which is often difficult to determine in fossil specimens. Low STET values indicate very similar overall shape (low scatter about the regression lines).
For this analysis, we used the STET statistic to assess the likelihood that TM 1517 and the Swartkrans crania represent the same species. If TM 1517 represents a different species from that which is represented by the Swartkrans robust australopithecines, STET values computed for Swartkrans specimens (compared against each other) should be significantly lower than the STETs derived from TM 1517-Swartkrans comparisons. A ranked-sums test19 was used to determine whether the Swartkrans-only STET values were significantly different from the values computed for the TM 1517-Swartkrans comparisons. This non-parametric test was appropriate in this case because the two samples were not independent: The same Swartkrans crania were used to compute STET values for within-Swartkrans and TM 1517-Swartkrans comparisons.
Finally, because the STET statistic is more reliable with increasing numbers of variables,18 the above procedure was repeated in a secondary analysis, omitting pair-wise comparisons that share less than ten variables, as in the case of the following pairs: TM 1517-SK 12, TM 1517-SK 79, SK 79-SK 12, and SK 12-SK 83. An advantage of the methods developed by Thackeray and colleagues15,16 and Lee and Wolpoff17,18 is that they facilitate assessment of fossil assemblages when certain specimens do not preserve all of the traits under study. Additionally, these methods provide a means of testing hypotheses about taxonomy, based on metric similarity. It should be noted that these methods only test null hypotheses - hypotheses of no significant difference.
RESULTS
The mean STET value for Swartkrans-only was 0.156 ± 0.118, and that of TM 1517-Swartkrans was 0.119 ± 0.104 (see Table 2). The Wilcoxon test showed the distributions are not significantly different (p = 0.3) The TM 1517-Swartkrans distribution was within the range of the Swartkrans distribution, with the exception of the TM 1517-SKW 11 comparison, which had a lower STET value than any comparison within Swartkrans alone (see Figure 2). That the TM 1517-Swartkrans distribution had a lower mean STET value than the Swartkrans-only distribution, highlights the morphological similarities of TM 1517 with many specimens from the Swartkrans sample.


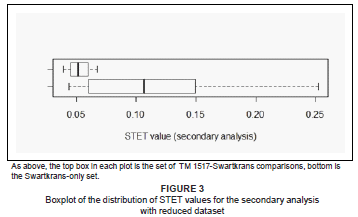
Figure 2 shows that within the Swartkrans-only distribution there is a major outlier, the comparison of SK 12-SK79; this is one of the comparisons preserving fewer than 10 variables in common. The secondary analysis, omitting the comparisons with less than 10 variables in common, produced similar results to the whole-sample analysis (see Figure 3). The mean STET for Swartkrans-only was 0.120 ± 0.070, and that of TM 1517-Swartkrans was 0.053 ± 0.012. In the reduced dataset, the TM 1517-Swartkrans STET values span an even smaller part of the lower range of the within-Swartkrans variation, further underscoring the similarity between TM 1517 and the Swartkrans fossils. A Wilcoxon ranked-sums test on this reduced dataset showed the TM 1517-Swartkrans and Swartkrans-only STET distributions to be significantly different (p = 0.02) at the p < 0.05 level. Because the TM 1517-Swartkrans STET values were generally lower than those within Swartkrans, this significant result is surprising, and further emphasises the morphological similarity between the crania from these two sites, as well as the great variation within Swartkrans itself.
DISCUSSION
The results show that one cannot reject the null hypothesis: That TM 1517 from Kromdraai and crania from Swartkrans sample the same species, Paranthropus (or Australopithecus) robustus. The TM 1517-Swartkrans STET values were generally lower than the within-Swartkrans values, indicating that not only is TM 1517 within the Swartkrans-only range of variation, but also that TM 1517 is generally more similar to Swartkrans crania than the Swartkrans crania are to each other. In fact, the lowest STET value from the analysis, indicating the two most similar specimens, is that of TM 1517 from Kromdraai and SKW 11 from Swartkrans Member 1's Hanging Remnant. In light of the fairly large variation displayed by the Swartkrans fossils, our results point to a relatively stable lineage of robust australopithecine in the Sterkfontein valley in the lower Pleistocene.
Our results indicate that the Kromdraai and Swartkrans robust australopithecines represent the same species. This is contrary to findings of Howell11, Grine7,9,10, and Schwartz and Tattersall12, whose conclusions were based largely on differences in dental sizes, proportions, and gross morphology between the two sites. Two issues arose with these authors' conclusions. Firstly, the decisions to make specific distinctions appear to be have been made from gross inspection of each site's summary statistics for given dental traits (e.g. M3 mesiodistal length), rather than from statistical tests. Along these lines, Fuller5 used a resampling procedure to compare the pooled Swartkrans-Kromdraai dental coefficients of variation (CV) to the CVs of modern humans, African apes, and fossil hominins. Her results showed that variation within the pooled Swartkrans-Kromdraai sample was statistically no greater than in most of the modern and fossil referents, leading her not to reject the hypothesis of a single species represented at the two sites.
Secondly, some Kromdraai specimens contributing to the seemingly specific differences between the two sites may actually relate to the presence of early Homo. For example, Grine9, 10 suggested that the teeth of a sub-adult from Kromdraai, KB 5223, represented a robust australopithecine species different from that which was represented at Swartkrans. However, Braga and Thackeray20 have shown, using qualitative and quantitative criteria, that KB 5223 is probably attributable to early Homo. Placing all other Kromdraai B dental specimens in the P. robustus sample along with Swartkrans specimens, these authors found it improbable that the KB 5223 postcanine dentition came from the same P. robustus sample, but rather more likely indicated the presence of Homo at Kromdraai In some non-metric traits, this specimen is similar to the TM 1536 M1 and dm1, also from Kromdraai. However, the authors noted that TM 1536 lacked the key diagnostic features of Homo molars, such as the absence of the hypoconulid on the dm1 and C6 on the dm2, present in KB 5223. Thus, the results of Braga and Thackeray20 corroborated those of Fuller5, indicating a high probability that the 'robust' dental specimens from Kromdraai and Swartkrans, with the exception of the KB 5223 dentition, sample a single species. Our results further demonstrate the craniodental affinities of the Kromdraai and Swartkrans robust australopithecine fossils.
CONCLUSION
From our morphometric analysis, we failed to reject the null hypothesis, therefore there is a very high probability that the type specimen of P. robustus, TM 1517, is the same species of robust australopithecine that is present at Swartkrans. Moreover, TM 1517 fit comfortably within the range of craniodental variation seen in Swartkrans, and is indeed morphometrically very similar to many of the Swartkrans fossils. Hence, the Swartkrans 'robust' australopithecines and TM 1517 can be considered conspecific, representing a single species, Paranthropus (Australopithecus) robustus.
ACKNOWLEDGEMENTS
Support for this research was provided by the Rackham Graduate School and the University of Michigan Department of Anthropology (ZC), and by the French Embassy in South Africa, the National Research Foundation (South Africa) and the Andrew Mellon foundation (JFT). We thank Dana Begun and Milford Wolpoff for helpful comments on earlier manuscripts, as well as two anonymous reviewers for helpful and insightful comments on an earlier draft. Our gratitude also goes to Stephany Potze, Lazarus Kgasi and Tersia Perregil at the Transvaal Museum for helpful access to the fossil material.
REFERENCES
1. Broom R. The Pleistocene anthropoid apes of South Africa. Nature. 1938;142:377-379. [ Links ]
2. Broom R. Another new type of fossil ape-man. Nature. 1949;163:57. [ Links ]
3. Tobias PV. The cranium and maxillary dentition of Australopithecus (Zinjanthropus) boisei, Olduvai Gorge. Cambridge University Press, Cambridge; 1967. [ Links ]
4. Brain CK. The hunters or the hunted? An introduction to African cave taphonomy, University of Chicago Press, Chicago; 1981. [ Links ]
5. Fuller K. An analysis of the probability of multiple taxa in a combined sample of Swartkrans and Kromdraai dental material. Am J of Phys. Anthropol. 1996;101:429-439. [ Links ]
6. Kaszycka KA. Status of Kromdraai: cranial, mandibular, and dental morphology, systematic relationships, and significance of the Kromdraai hominids, CNRS Editions, Paris; 2002. [ Links ]
7. Grine FE. New craniodental fossils of Paranthropus from the Swartkrans formation and their significance in 'robust' Australopithecine evolution. In: Grine FE. Evolutionary history of the 'robust' Australopithecines. New York: Aldine, 1988; p. 223-246. [ Links ]
8. Grine FE. New hominid fossils from the Swartkrans formation (1979-1986 excavations): Craniodental specimens. Am J Phys Anthropol. 1989;79:409-449. [ Links ]
9. Grine FE. Description and preliminary analysis of new hominid craniodental fossils from the Swartkrans formation. In: Brain CK. Swartkrans: A cave's chronicle of early man. Pretoria: Transvaal Museum, 1993; p. 75-116. [ Links ]
10. Grine FE. A new juvenile hominid (Mammalia: Primates) from Member 3, Kromdraai Formation, Transvaal, South Africa. Ann Transvaal Mus. 1982;33:165-239. [ Links ]
11. Howell FC. Hominidae. In: Maglio VJ, Cooke HBS, editors. Evolution of African Mammals. Cambridge: Harvard University Press, 1978; p. 154-248. [ Links ]
12. Schwartz JH, Tattersall I. The human fossil record vol. 4. craniodental morphology of early hominids (Genera Australopithecus, Paranthropus, Orrorin), and overview. Hoboken: John Wiley & Sons Inc.; 2005. [ Links ]
13. Lockwood CA. Sexual dimorphism in the face of Australopithecus africanus. Am J Phys Anthropol. 1999;108:97-127. [ Links ]
14. Wolpoff MH. Interstitial wear. Am J Phys Anthropol. 1971;34:205-228. [ Links ]
15. Thackeray JF, Bellamy CL, Bellars D, et al. Probabilities of conspecificity: application of a morphometric technique to modern taxa and fossil specimens attributed to Australopithecus and Homo. S Afr J Sci. 1997;93:195-196. [ Links ]
16. Thackeray JF. Probabilities of conspecificity. S Afr J Sci. 2007;103:1. [ Links ]
17. Wolpoff MH, Lee SH. The Late Pleistocene human species of Israel. Bull Mem Soc Anthropol Paris. 2001;13:291-310. [ Links ]
18. Lee SH, Wolpoff MH. Habiline variation: A new approach using STET. Theory Biosci. 2005;124:25-40. [ Links ]
19. Siegel S. Nonparametric statistics for the behavioral sciences. New York: McGraw-Hill; 1956. [ Links ]
20. Braga J, Thackeray JF. Early homo at Kromdraai B: probabilistic and morphological analysis of the lower dentition. C R Palevo. 2003;2:269-279. [ Links ]
 Correspondence to:
Correspondence to:
Zachary Cofran
Department of Anthropology
University of Michigan, 101 West Hall 1085 S. University Ave
Ann Arbor, MI 48109, United States of America
email: zacharoo@umich.edu
Received: 15 July 2009
Accepted: 05 Nov. 2009
Published: 11 Mar. 2010
This article is available at: http://www.sajs.co.za


{kind=link}