










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.106 n.1-2 Pretoria Jan./Feb. 2010
RESEARCH ARTICLE
Historical mammal distribution data: how reliable are written records?
Andre F. Boshoff; Graham I.H. Kerley
Centre for African Conservation Ecology, Nelson Mandela Metropolitan University, Port Elizabeth, South Africa
ABSTRACT
Written historical records are widely used to estimate the previous distributions of the larger mammals in southern Africa. However, such records have some limitations and the use of those older than 100 years has been questioned. Written historical records, from the broader Eastern Cape, South Africa, were investigated to examine this contention critically. They were classified according to record quality (acceptability of identification and precision of locality) and analysed according to two levels: 'all' species and 'noticeable' versus 'non-noticeable' species. Records that comprise acceptable identification and precise locality information are the most suitable for mapping historical distributions; they form 33% of the records for the 27 mammal species analysed. A further 49% of the records have acceptable identification but imprecise locality information; they can fulfil a useful function when supported by records where both parameters are of good quality. Thus, the majority (82%) of written historical records from the study area are useful for compiling historical distribution maps and the quality of these records is consistent back to 1750 for this data set. The number and quality of written historical records varies between species. Historical distribution data should be evaluated for reliability (quality) and degree of usefulness, rather than simply discarded a priori.
Keywords: mammals; historical distribution; written records; record reliability; translocation policy; South Africa
INTRODUCTION
Distributional data form the basis of the science of zoogeography, which has applications in, inter alia, ecology and conservation.1,2,3,4 These data are essential for establishing the link between the occurrence of a species on a landscape and its typical habitat.5 They are also important for investigating the presence, absence or nature of spatial and temporal patterns and trends of a species across a landscape, as well as guiding the restoration of species to areas from which they have been extirpated.6,7 The benefits of combining the (usually) parallel disciplines of environmental history (being primarily an interpretation of the past) and conservation biology (which primarily seeks to shape the future) are largely the increased confidence in the understanding of distribution information, and hence implementation of any management actions.8
Historical distribution data refer to written records found in the diaries, journals and correspondence of people, most notably explorers, settlers, hunters, missionaries and naturalists during historical times, namely that period for which such records are available. The value of historical distribution data has been widely recognised for aiding the reconstruction of animal assemblages for a region, over a certain time period, and for understanding the structure, function and processes of ecosystems by ecologists and conservation biologists.5,6,7,8,9,10,11
There are many examples in the literature of the use of published historical distribution records, based on, inter alia, sightings, to estimate historical ranges of mammals (especially the larger ones) for a range of applications. For example, published historical information was used to describe changes in the distribution of the caribou Rangifer tarandus in Quebec Province, Canada12, to measure range contractions and expansions of North American carnivores and ungulates13, to determine historical mammal species richness in Canadian national parks14, to establish mammal distribution in the United States in the early 19th century, based on the well-known Lewis and Clark journals15, to study global range contraction dynamics16 and to identify areas where the establishment of protected areas is required for effective wildlife conservation in Africa.17 On a continental scale, for example in North America, oral narratives and actual field sightings have provided insight into historical mammal distribution.17,18
In South Africa, historical distribution data have been used to inform the spatial distribution component of the conservation plan for the globally critically endangered black rhinoceros Diceros bicornis.19 The value placed on such data is exemplified by one of the principles that govern the re-establishment or removal of species into South Africa's national parks, namely this 'should only take place where there is good evidence that the species occurred in the area in historical times'.20 Notably, these data have provided the basis for determining national and regional policies and principles for the translocation of indigenous and alien mammals, as embodied in the National Environmental Management: Biodiversity Act (Act 10 of 2004). Historical distribution data have also been successfully used for landscape-level conservation planning purposes for the larger mammals in the biodiversity-rich Cape Floristic Region.7,21
Notwithstanding the wide use of written historical records for depicting historical distributions of, inter alia, mammals, caution has been urged in the interpretation and use of some of these data.10 For example, the contents of historical documents, such as the journals kept by Lewis and Clark, must be 'evaluated for their accuracy and thoroughness'22, especially as far as ecological information is concerned.23 Identifications can be problematic, with either no, short or inadequate descriptions of the species provided with the record. While large and charismatic species are relatively easy to identify from written accounts, this is not the case for taxa with many similar-appearing species.24 There were significant discrepancies in a series of 'independent' range maps of the gray wolf Canis lupus in North America, which were compiled largely from museum specimens and the diaries and journals of early trappers, hunters, settlers and explorers; problems with identification and record location description are suggested as two important reasons for these discrepancies.25 The pitfalls of written records, ranging over a variety of sources from explorer's accounts to local histories to census records, have been highlighted.26 Here, the principal problems in using written evidence relate to nomenclature (compounded by the lack of widely accepted names), geographical location (exacerbated by a lack of geographical knowledge) and the personal biases of authors.
In the South African context, certain weaknesses in early (historical) distribution records have been highlighted.11,24,27,28,29 Recently, it has been postulated that while there is a high level of certainty attached to historical distribution records from the past 100 years, this decreases rapidly between 100 and 500 years ago and therefore records from more than 100 years ago may have limited value.30 In this discourse, however, it is important to distinguish between the different types of historical distribution records, which comprise sight records, rock art and rock engraving records, and archaeological and ethnographic records. There are pitfalls associated with the interpretation and use of all of these types of records. The present study, however, deals only with written historical records, which have been widely used to reconstruct early distribution patterns of mammals on southern African landscapes.24,29,30,31,32,33,34
It is important, though, to recognise the limitations associated with written historical records. Most notably, early observers did not keep systematic records (i.e. they did not record every animal that they saw, or attempt to achieve equal spatial coverage) and they did not always document and/or publish their observations. They also tended to confuse the identification of similar looking species, keeping to flat open areas that could be traversed by horse, ox and wagon (resulting in few records from mountainous areas, secluded valleys and densely vegetated habitats) and they did not normally travel at night (resulting in very few records of nocturnal species).29,33 In North America, early descriptions of fauna and flora are usually restricted to species that were easily visible from major transportation routes, such as rivers and frequently travelled roads.26 Consequently, early written records should be interpreted with circumspection.
The recent publication of Historical incidence of the larger mammals in the broader Eastern Cape, provides an extensive collection of historical records of these mammals in this region.29 We employed this data set, which was initially used to create detailed distribution maps, to assess and comment critically upon the reliability of historical distributional data and specifically those emanating from written records.
METHODS
Distributional data pertaining to selected medium- to large-sized (= larger) mammals in the Eastern Cape (Table 1) were extracted from the distributional information documented in Skead29 and geo-referenced (at 1' x 1' resolution). For each record of each species, both the year in which the record was made (if available) and the record type, were listed. 'Record type' refers to the scheme used in Skead29 to classify each distribution record according to one of five categories, based on record quality:
• Type 1 - accurate identification; precise locality (specimen)
• Type 2 - accurate identification; precise locality (sighting)
• Type 3 - accurate identification; imprecise locality
• Type 4 - questionable identification; precise locality
• Type 5 - questionable identification; imprecise locality.
To achieve consistency, all record interpretation, categorisation and geo-referencing was done by the same person, who is highly familiar with the mammal species and the geography and topography of the Eastern Cape. The following criteria guided the process: (1) how well the species was described, (2) the accuracy of the description of the behaviour of the species (where available), (3) the amount of locality detail provided, and (4) the accuracy of the description of the habitat (where available). With regard to the first criteria, the identity of a species was accepted only if the description (sometimes accompanied by an illustration) was sufficient to establish its identity beyond reasonable doubt.29
Given that the time (year) of death of animals that provided specimens (e.g. bones, teeth, tusks) is not known, Type 1 records cannot be used to indicate the period when the species was living on that particular landscape and they were therefore excluded from the analyses. In the definitions used above, a 'precise' locality is one where the sighting falls within an area with a diameter of < c. 5 km, whereas an 'imprecise' locality is one where the sighting falls within an area with a diameter of < c. 50 km, but > 5 km.
Only Type 2 and Type 3 records, for the period 1750-1924 (see below), were used to calculate the number of historical records per species. The common and widespread species (such as chacma baboon Papio hamadryas, vervet monkey Cercopithecus pygerythrus, black-backed jackal Canis mesomelas, caracal Caracal caracal, common duiker Sylvicapra grimmia, steenbok Raphicerus campestris, bushbuck Tragelaphus scriptus) and a range of shy and unobtrusive species (such as the otters, the foxes, African wild cat Felis silvestris, blue duiker Cephalopus monticola), all of which did not receive comprehensive documentation in Skead29, were excluded from this analysis.
Analysis and interpretation refers to (1) all 27 larger mammal species, and (2) a subset of seven 'noticeable' and seven 'non-noticeable' larger mammal species; the subset was selected according to a combination of the number of records per species and information in Skead29.
A 'noticeable' species is one that appears to have been easily recognised (identified) and, as a result of its behaviour, habitat and abundance, was probably relatively easily seen or noticed by observers in historical times (18th, 19th and early 20th century).29 A 'non-noticeable' species is one that was easily confused with other, similar, species or was not easily seen or noticed by observers in historical times due to its behaviour, habitat or rarity. 29 Thus, 'noticeable' is a catchword that combines two broad parameters, namely the relative ease with which a species could be identified and the relatively ease with which it could be observed (i.e. seen) in the field.
For this study we have taken 'historical' to mean as far back as 250 years (i.e. about 1750) because it was only in the latter half of the 18th century that European missionaries, hunters, travellers and settlers started becoming prominent in the region now known as the Eastern Cape, with written accounts of a number of the animals being made by some of them.29 Prior to 1750, records in South Africa are too sparse to enable meaningful analysis. Each historical record was allocated to a 25-year time period, as some years had too few records for analysis. The following time periods apply:
• 1750-1774
• 1775-1799
• 1800-1824
• 1825-1849
• 1850-1874
• 1875-1899
• 1900-1924.
Furthermore, although there are some distribution records in Skead29 for the post-1924 period, these have not been included in the analysis, mainly because Skead did not focus on this period for his literature search for historical records, and because the 1920s are considered to have witnessed the start of 'organised' zoology in southern Africa, demonstrated by an increase in field-collecting expeditions, the growth of natural museum collections, the advent of handbooks, and the development of modern taxonomy, as exemplified by the work of, notably, Vivian Fitzsimons, John Hewitt, Austin Roberts and GC Shortridge.35,36,37
The results of the analyses are expressed as, (1) the number of records within each time period, for each record type, and (2) the proportion (%) of each such type, within each time period. The latter reflects the relative importance of the various types of data. Differences in the numbers of records, per type, and the proportion of record types, between the time periods, were explored using χ2 analyses.
A distribution map (Figure 1) for a representative species (the true quagga Equus quagga) illustrates the contribution made by (mainly) Type 2 and Type 3 records. The years in which certain species were last recorded in the Eastern Cape, or by when their numbers had been drastically reduced in this region, were taken from Skead29.
RESULTS
The number of records per species varies from 3 (klipspringer) to 120 (buffalo) (Table 1). Type 2 and 3 records, together, dominate the record types for the 'all' species and 'noticeable' species data sets, whereas the proportions of record types are evenly spread among the types for the 'non-noticeable species' data set (Table 2).
The map of Type 2 and Type 3 records for the true quagga (Figure 1) illustrates the congruence of the former with the latter. Thus, even though Type 3 records do not have precise locality attributes, when plotted in conjunction with Type 2 records, which do have these attributes, they are supported. Inspection of such maps can also be used to identify outliers among Type 3 data records.
'All' species
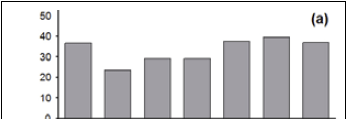
There are few Type 2 and Type 3 records from the 1750-1774 period, constraining the analysis of the data for this period (Figure 2, a and b). The number of Type 2 records varies between the remaining six 25-year time periods (1775-1924) (χ 2 = 39.8; p < 0.001; df = 5), as it does for Type 3 records (χ 2 = 297; p < 0.001; df = 5). The number of Type 2 and Type 3 records per time period decreases, overall, from the 1775-1799 period to the 1900-1924 period, inclusive (Figure 2, a and b).




The number of Type 4 records varies between the six 25-year time periods (1775-1924) (χ 2 = 28.6; p < 0.001; df = 5), as it does for Type 5 records (χ 2 = 32.1; p < 0.001; df = 5) (Figure 2, c and d). The numbers of Type 4 and Type 5 records increase, overall, between the 1750-1774 period and the 1825-1849 period, after which they show a marked decrease (Figure 2, c and d).
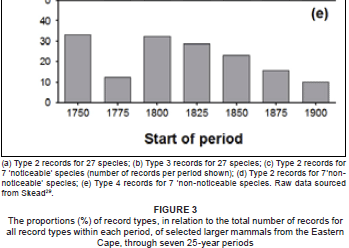
The proportions of Type 2 (χ2= 6.2; p > 0.05; df = 5) and Type 3 (χ 2 = 11.8; p = 0.038; df = 5) records are relatively stable over time, being consistently around 33% and 49% of the records, respectively (Figure 3, a and b). There is thus no indication of the predicted decline in these proportions for older records. The proportions of Type 4 and Type 5 records show much temporal variation, and there are no clearly discernible patterns or trends; consequently, these data are not shown here.

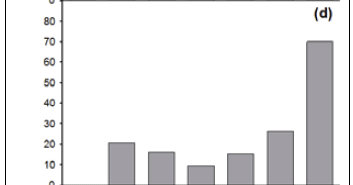
'Noticeable' versus 'non-noticeable' species
When the 27 species are ranked according to the number of records (Figure 4), it is evident that there is a positive relationship between the independently derived level of 'noticeability' and the number of records per species. The relatively limited number of records for the black rhinoceros may reflect its scarcity rather than its noticeability.
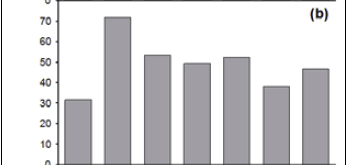
The number of Type 2 records for the selected 'noticeable' species varies between the last six 25-year time periods (1775-1924) (χ2= 65.9; p < 0.001; df = 5) (Figure 3c). After low initial reporting for the period 1750-1774, there is, overall, a temporal decline in the number of Type 2 records of the selected 'noticeable species'. However, the proportion of Type 2 records for 'noticeable' species (around 35% of the records, from 1750 to 1924) is stable over time (χ2= 3.97; p = 0.68; df = 6).
On average, Type 2 records formed only 18% of the records of the selected 'non-noticeable' species in the early time periods (1775-1899) (Figure 3d); this proportion increased to 70% in the 1900-1924 period. There is a temporal decrease in the proportion of Type 4 records as the most recent period is approached; except for the proportion for the 1775-1799 period, which is anomalously low (Figure 3e).
Exterminations and near-exterminations
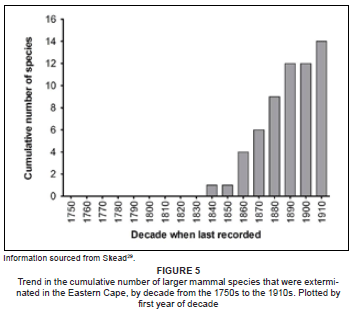
A total of 14 species (brown and spotted hyaenas, cheetah, lion, African wild dog, black rhinoceros, quagga, Cape warthog, hippopotamus, eland, black wildebeest, red hartebeest, blesbok, gemsbok) are known to have been exterminated in the Eastern Cape (as defined in Skead29, Figure 5) during the historical period under review (Table 1). These exterminations started in the 1840s and ended in the 1910s (Figure 5). In addition to these exterminations, a further seven species (leopard, African buffalo, southern reedbuck, springbok, African elephant, serval, Cape mountain zebra) underwent significant decreases in numbers and ranges in this region, leading in some cases to near-extermination. In the case of the latter group of species, their decline also took place mainly during the latter half of the 19th century and the early part of the 20th century.29
DISCUSSION
Some species had physical, behavioural or habitat characteristics that aided the determination of their identity. For example, reference to 'an erectly held tail whilst running' identified the Cape warthog beyond doubt, and prevented confusion with records of the bushpig. Furthermore, there is often confusion in the written accounts about the identities of the similar looking grey rhebok and the two reedbucks (mountain and southern); here, reference to the habitat as 'marshy' confirmed the identity of the southern reedbuck. Similarly, locality and described habitat could be used, in certain cases, to help distinguish between records of the true quagga (flat, or relatively flat, areas) and the Cape mountain zebra (mountainous areas).
The high variation in number of records per species (Table 1) is considered to reflect a variety of factors, including: distinctiveness (i.e. identification potential), behaviour (diurnal or nocturnal, group size [solitary, family groups, herds], shy/unobtrusive), habitat (open or closed vegetation, mountains or valleys and plains), rarity (naturally scarce species, e.g. black rhino and cheetah), global range (marginal in the Eastern Cape, e.g. gemsbok) or economic value (source of tusks, hides and meat).
For example, elephants were extensively hunted for ivory38 and they are large, social and are typically found in lowland habitats close to water; consequently, this species is well represented in the written historical records. In contrast, the klipspringer is small, specific to rugged habitats and of very limited economic value; it is very poorly represented in the historical records. This clearly indicates that there are species-specific biases in the historical reporting frequency and this bias should be considered when evaluating the utility of records for each species.
Trends over time: 'all' species
It is postulated that the low number of records from the 1750-1774 period is a consequence of the relatively few literate observers in the Eastern Cape at that time; settlement by European colonists in this region only began to increase towards the end of the 18th century, with large numbers arriving during the 19th century.39,40
The temporal decrease in the number of Type 2 records (Figure 2a) is counterintuitive, as it would be expected that reporting would improve in more recent periods. This would presumably have been expected to reflect both the increasing number of observers, as well as the improved information describing each species (hence more accurate identifications - see also below regarding improved geographic knowledge). This trend may, however, be explained as a reflection of the progressive decrease in the occurrence and abundance of certain species, especially the large and charismatic ones.29 This hypothesis is supported by the temporal increase in the number of exterminations in the region in question (Figure 5) and also the fact that five of the seven species that underwent major declines, or were almost exterminated, are large and/or charismatic species. If valid, the above hypothesis strengthens the use of certain historical records to reflect not only the distribution of some species, but also their relative abundance over time.
The temporal decrease in the number of Type 3 records (Figure 2b) may additionally reflect the fact that, over time, geographic locations (e.g. towns, roads, railway sidings) were established and landscape features (e.g. mountains, rivers) were named, in the language of the recorders. This would have facilitated the location of sightings on the landscape by early recorders. Therefore, we can predict that those observers who were able to identify the species (one of the requirements for a Type 2 record) were more recently also able to provide an accurate location, thus a greater number of the correctly identified species would also have been precisely located. This effect is not clearly demonstrated by a complimentary increase in the number or proportion of Type 2 records (discussed above), reflecting an overall decline in the total number of Type 2 and 3 records.
The Type 2 data are robust over time (Figure 3a) and consistently represent about 33% of the records. This does not support Bernard and Parker's30 contention that older records are less reliable. In contrast to the observed decline in the number of Type 3 records (Figure 2b), the proportion of this group of records also remains consistent over time (Figure 3b). The absence of a decrease here may be due to the relatively low numbers (a decrease) of Type 4 and 5 records in the periods after 1849 (Figure 2). The latter pattern is also likely to be influenced by the decline or extermination of several prominent species in the region; exterminations started in the Eastern Cape in the 1840s (Figure 5).
Only 17.6% of the records are Type 4 or Type 5 (Table 2), and therefore of little use for distribution mapping purposes. On the other hand, 33% of the records are Type 2 (Table 2) and therefore of good quality, which enables them to be used on a 'stand-alone' basis for distribution mapping purposes. A further 49.1% of the records are Type 3 (Table 2) and, because of their lack of precise locality data, cannot be used on a 'stand-alone' basis; they can, however, be used in support of the Type 2 records (see below). Therefore, overall, some 82% of the historical distribution records can be used for mapping purposes.
The use of Type 2 records for historical distribution maps is appropriate as the quality of these records has remained consistent over time, that is, back to 1750 (Figure 3a). The same holds for Type 3 records (Figure 3b). These data indicate that it is important to evaluate the quality of each historical record critically, both in terms of the correctness of identification and the level of precision of the location. This process will allow doubtful records to be treated as such. It is thus apparent that historical records cannot be summarily discarded on the basis of age alone, as proposed by Bernard and Parker30. This is supported by the view of Morrison9, who states that older data, per se, are not of lower quality, but are rather a product of earlier and different data gathering techniques. Rackham10 argues for the critical interpretation of occurrence records in Great Britain dating back to the Middle Ages (1 000 years).
'Noticeable' versus 'non-noticeable' species
The temporal decline in the number of Type 2 records for 'noticeable' species is ascribed to the progressive disappearance from the landscape, or significant decline in numbers, of the species selected for this analysis (Table 1; Figure 5). This is supported by the fact that five of the seven species that underwent major declines or were almost exterminated are large and/or charismatic species and hence were included in this category of 'noticeable' species. As with the 'all species' data, the use of historical Type 2 data for estimating historical distribution patterns of 'noticeable' species is appropriate, as the quality of these records has remained robust over time, that is, back to 1750. Similarly, as for the 'all species' findings, the proportion of Type 3 records is high (55.1%; Table 2) and when these records are supported by Type 2 records, almost 90% of all records (Table 2) can be used to map distributions of 'noticeable' species. This supports Cramer and Mazel's27 contention that for large charismatic species, such as the giraffe Giraffa camelopardalis, it is unlikely that early recorders would fail to mention them. This again emphasises the point that the utility of historical records will vary between species, but there is no evidence that this declines with the age of the record.
The increased proportion of Type 2 records of 'non-noticeable' species after 1900 is considered to be a reflection of the improved levels, over time, of identification of the species selected for this analysis. This effect is therefore mirrored in the temporal decline in the proportion of Type 4 records of 'non-noticeable' species. Despite the tendency of observers to confuse the identity of these species (a key feature of Type 2 and 3 records),29 it must be recognised that over time 49.1% of the records comprise Type 2 and Type 3 records (Table 2). These data indicate that historical records can still be very useful in indicating the distributions of even 'non-noticeable' species. Again, the discrepancy between the proportions of Type 2 and 3 records for 'noticeable' species versus those of 'non-noticeable' species (89.8% vs 49%) clearly emphasises the point that historical records vary in quality between species.
In contrast to the trends for the 'all species' and 'noticeable species' data, the trends of improved precision in historical records for the 'non-noticeable' species over time does support Bernard and Parker's30 statement that historical records decline with age (i.e. increase in Type 4 records with age). However, the data also clearly show that there is still a significant proportion of usable records (49%) as one goes back in time. This again emphasises the need to examine the data critically, rather than discard all records on the basis of some arbitrary age cut-off, as advocated by Bernard and Parker30.
Time scales and changes in communities
The study area experienced considerable fluctuations in climate during the Late Quaternary, with the Last Glacial Maximum supporting palaeocommunities typical of colder and drier conditions.40 Modern communities, including the large mammal faunal assemblages, emerged in the last 5 000 years under what are considered to be typically modern climatic conditions.41 The moderately stable climatic conditions that have prevailed since then, indicate that anthropogenic effects have been the main drivers of change in the distribution and abundance of the larger mammals, mainly within the past 2 000 years, but especially within the past 400 years.41,42,43,44 In support of this, it is noteworthy that the extermination of 14 larger mammal species in the Eastern Cape occurred over a relatively short period of 70 years (1840s-1910s, Figure 5), when the region was starting to experience the combined impacts of excessive hunting45, habitat transformation and degradation,39 and predator control46,47. The availability and analysis of the historical records assessed here are therefore crucial in our understanding of the changes experienced by these communities.
The issue of 'false positives'
Estimates of historical species distributions may be prone to error because a species may be considered to have been present in an area, whereas it was not. This occurs because the historical distribution map reflects the '"extent of occurrence" and not necessarily the "area of occupancy"', thereby introducing the issue of 'false positives'.13,14 Therefore, owing to the non-systematic nature of historical sight records, it cannot be assumed that a species historically occurred everywhere within a convex polygon created by joining the outermost distribution records, to create a so-called 'extent of occurrence'.
The effects of 'false positives' on the estimation of a species' distribution can be ameliorated by combining historical distribution records with species' ecological habitat requirements, including the use of an expert knowledge approach5,7,21 or by modelling48. For example, where written records may indicate the historical incidence of a species in a general area, it is possible re-introduction locally (e.g. into a national park) must depend on the existence of suitable habitat.
Imprecise location sight records
Even though Type 3 records do not have precise locality attributes, they can, when plotted in conjunction with Type 2 records, which do have these attributes, be supported by the latter (Figure 1). Hence, Type 3 records should not be discarded from distribution maps, unless they are clear outliers.
General statements made by early observers about some of the larger mammals that they saw, in relation to prominent landscape features (e.g. mountain ranges, rivers), can also be useful in establishing the regional distribution of certain species, notwithstanding the fact that, in such cases, the locality information is of a highly imprecise nature. For example, based on early observer accounts, the Orange (Gariep) River featured prominently as the southernmost limit in the distribution of the blue wildebeest Connochaetes taurinus and Burchells' zebra Equus burchelli in South Africa.49 While such data may be frustrating to ecologists wanting to estimate distributions of species at a fine scale, they are nevertheless of significant value in determining policy. In this case, for example, it may be concluded that blue wildebeest and Burchell's zebra should be considered extralimital if introduced south of the Orange River.
CONCLUSION
The analysis and interpretation of written historical record data from Skead29, as described in this paper, are considered to be influenced by the interplay between the following five variables:
• A temporal and spatial increase in the number of literate observers (i.e. as the region became explored, hunted, settled and farmed, by European colonists from the west to the east).
• The progressive impacts on the region's indigenous large mammal fauna from excessive hunting, habitat transformation and degradation, and predator control, leading, for example, to the decline of certain species to near-extermination and to the extermination of others.
• A temporal increase in geographic knowledge (e.g. naming of towns, rivers, mountain ranges), resulting in more precise record location data.
• A temporal increase in the ability of observers to identify the various larger mammal species.
• The variation in the number and quality of records between species.
Notwithstanding these constraints, discernible temporal patterns and trends in the reliability of the data were evident.
Records that comprise acceptable identification and precise locality information (33% of all records in this study of 27 species) are particularly useful for estimating historical distributions. Additionally, records that have acceptable identification but imprecise locality information (49% of all records in this study) can fulfil a useful function when they are supported by records where both parameters are of good quality. In both cases, the quality of the records is consistent back to 1750. Therefore, the majority (82%) of written historical records from the Eastern Cape29 are useful for compiling historical distribution maps, despite their age. Written records with questionable identification but good location attributes and those with questionable identification and imprecise location data, are of little or no use for this purpose.
The usefulness of written historical records can be enhanced if they are combined with other information, particularly the known ecological requirements of the species concerned (as done by Boshoff and Kerley5), but also the location of the records in relation to major physical features on the landscape.
Thus, written historical distribution records can serve a useful function, but they should be evaluated for reliability and degree of usefulness, rather than simply discarded a priori.
ACKNOWLEDGEMENTS
We thank Derek du Preez for creating Figure 1 and Marietjie Landman for assistance with data analysis. We are particularly grateful to the late C.J. (Jack) Skead for stimulating our interest in the value and study of historical mammalian distribution records.
REFERENCES
1. Fjeldsa J, Burgess ND, Blyth S, De Klerk HM. Where are the major gaps in the reserve network for Africa's mammals? Oryx. 2004;38:17-25. [ Links ]
2. Lomolino MV, Channell R. Splendid isolation: patterns of geographic range collapse in endangered mammals. J Mammal. 1995;76:335-347. [ Links ]
3. Sanderson EW, Redford KH, Chetkiewicz CB, et al. Planning to save a species: the jaguar as a model. Conserv Biol. 2002;16:58-72. [ Links ]
4. Sanderson EW, Redford KH, Weber B, et al. The ecological future of the North American bison: conceiving long-term, large-scale conservation of wildlife. Conserv Biol. 2008;22:252-266. [ Links ]
5. Boshoff AF, Kerley GIH. Potential distributions of the medium- to large-sized mammals in the Cape Floristic Region, based on historical accounts and habitat requirements. Afr Zool 2001;36:245-273. [ Links ]
6. Egan D, Howell EA, editors. The historical ecology handbook: a restorationist's guide to reference ecosystems. Washington DC: Island Press; 2005. [ Links ]
7. Kerley GIH, Pressey RL, Cowling RM, Boshoff AF, Sims-Castley R. Options for the conservation of large and medium-sized mammals in the Cape Floristic Region hotspot, South Africa. Biol Conserv. 2003;112:169-190. [ Links ]
8. Meine C. Its about time: conservation biology and history. Conserv Biol1999;13:1-3. [ Links ]
9. Morrison ML. Techniques for discovering historic animal assemblages. In: Egan D, Howell EA, editors. The historical ecology handbook: a restorationist's guide to reference ecosystems. Washington DC: Island Press, 2005; p. 295-316. [ Links ]
10. Rackham O. The history of the countryside: the classic history of Britain's landscape, fauna and flora. London: Phoenix Press; 2000. [ Links ]
11. Skead CJ. The use of early historical references in the study of bird and mammal distribution. Ann Cape Prov Mu. 1962;2:138-142. [ Links ]
12. Courtois R, Ouellet JP, Gingras A, Dussault C, Breton L, Maltais J. Historical changes and current distribution of caribou, Rangifer tarandus, in Quebec Can Field-nat. 2003;117:399-414. [ Links ]
13. Laliberte AS, Ripple WJ. Range contractions of North American carnivores and ungulates. BioScience. 2004;54:128-138. [ Links ]
14. Habib LD, Wiersma YF, Nudds TD. Effects of errors in range maps on estimates of historical species richness of mammals in Canadian national parks. J Biogeogr 2006;30:375-380. [ Links ]
15. Laliberte AS, Ripple WJ. Wildlife encounters by Lewis and Clark: spatial analysis of interactions between native Americans and wildlife. BioScience. 2003;53:94-103. [ Links ]
16. Channell R, Lomolino MV. Trajectories to extinction: spatial dynamics of the contraction of geographical ranges. J Biogeogr. 2000;27:169-179. [ Links ]
17. Feldhamer GA, Thompson BC, Chapman JA. Wild mammals of North America: biology, management and conservation Baltimore: John Hopkins University Press; 2004. [ Links ]
18. Hall ER, Kelson KR. The mammals of North America. New York: Ronald Press; 1959. [ Links ]
19. Emslie RH. Conservation plan for the black rhino (Diceros bicornis) in South Africa 2003-2012. Revised Edition. SADC Rhino Management Group; 2007. [ Links ]
20. SANParks. South African National Parks coordinated policy framework governing park management plans. Pretoria: South African National Parks; 2006. [ Links ]
21. Boshoff AF, Kerley GIH, Cowling RMC. A pragmatic approach to estimating the distributions and spatial requirements of the medium- to large-sized mammals in the Cape Floristic Region. Divers Distrib. 2001;7:29-43. [ Links ]
22. Wood WR. Ethnohistory and historical method. Archaeol Meth Theory. 1990;2:81-110. [ Links ]
23. Forman RT, Russell EW. Evaluation of historical data. Ecol Soc Bull. 1983;64;5-7. [ Links ]
24. Rookmaaker LC. The zoological exploration of southern Africa 1650-1790. Rotterdam: AA Balkema; 1989. [ Links ]
25. Shelton SL, Weckerly FW. Inconsistencies in historical geographic range maps: the gray wolf as an example. Calif Fish Game. 2007;93:224-227. [ Links ]
26. Edmonds M. The pleasures and pitfalls of written records. In: Egan D, Howell EA, editors. The historical ecology handbook: a restorationist's guide to reference ecosystems. Washington DC: Island Press, 2005; p. 73-100. [ Links ]
27. Cramer MD, Mazel AD. The past distribution of giraffe in KwaZulu-Natal. S Afr J Wildl Res 2007;37:197-201. [ Links ]
28. Goodman PS, Tomkinson AJ. The past distribution of giraffe in Zululand and its implications for reserve management. S Afr J Wildl Res. 1987;17:28-32. [ Links ]
29. Skead CJ. Historical incidence of the larger mammals in the broader Eastern Cape, 2nd ed. Centre for African Conservation Ecology, Nelson Mandela Metropolitan University, Port Elizabeth; 2007. [ Links ]
30. Bernard RTF, Parker DM. The use of archaeological and ethnographical information to supplement the historical record of the distribution of mammalian herbivores in South Africa. S Afr J Sci 2006;102:117-119. [ Links ]
31. Du Plessis SF. The past and present geographical distribution of the Perissodactyla and Artiodactyla in southern Africa. MSc thesis, University of Pretoria, 1969. [ Links ]
32. Roberts A. The mammals of South Africa. Cape Town: Central News Agency; 1951. [ Links ]
33. Skead CJ. Historical mammal incidence in the Cape Province: Vol.1 - the Western and Northern Cape Cape Town: Provincial Administration of the Cape of Good Hope; 1980. [ Links ]
34. Smithers RHN. The mammals of the southern African subregion. University of Pretoria; 1983. [ Links ]
35. Brain CK, Roberts A. A lifelong devotion to South Africa's birds and beasts. Cape Town: John Voelcker Bird Book Fund; 1998. [ Links ]
36. Hewitt J. A guide to the vertebrate fauna of the Eastern Cape Province. Part 1 - mammals and birds. Grahamstown: Albany Museum; 1931. [ Links ]
37. Shortridge GC. The mammals of South West Africa. Vol I & II. London: Heinemann; 1934. [ Links ]
38. Carruthers J, Boshoff A, Slotow R, Biggs H, Avery G, Matthews W. The elephant in South Africa: history and distribution. In: Scholes RJ, Mennell KG, editors. Assessment of South African elephant management. Johannesburg: Witwatersrand University Press, 2008; p. 23-83. [ Links ]
39. Beinart W. The rise of conservation in South Africa: settlers, livestock and the environment 1770-1950. London: Oxford University Press; 2003. [ Links ]
40. Mostert N. Frontiers. London: Jonathan Cape; 1992. [ Links ]
41. Deacon J, Lancaster N. Late quaternary paleo-environments of southern Africa. London: Clarendon Press; 1988. [ Links ]
42. Klein RG. The taxonomic status, distribution and ecology of the blue antelope, Hippotragus leucophaeus (Pallas, 1766). Ann S Afr Mus. 1974;65:99-143. [ Links ]
43. Klein RG. Paleo-environmental implications of quaternary large mammals in the Fynbos region. In: Decaon HJ, Hendey QB, Lambrechts JJN, editors. Fynbos paleoecology: a preliminary synthesis. South African National Scientific Programmes Rep. 1983;75:116-138. [ Links ]
44. Radloff FGT. The ecology of large herbivores native to the coastal lowlands of the Fynbos Biome in the Western Cape, South Africa. Ph.D thesis, University of Stellenbosch, 2008. [ Links ]
45. Roche C. Ornaments of the Desert - springbok treks in the Cape Colony, 1774-1908. M.Sc. thesis, University of Cape Town, 2004. [ Links ]
46. Beinart W. The night of the jackal: sheep, pastures and predators in the Cape. Past and Present. 1998:158;172-206. [ Links ]
47. Van Sittert L. Keeping the enemy at bay: the extermination of wild carnivores in the Cape Colony, 1889-1910. Environ Hist 1998;3:311-332. [ Links ]
48. Illoldi-Rangel P, Sánchez-Cordero V, Peterson AT. Predicting distributions of Mexican mammals using ecological niche modelling. J Mammal 2004;85:658-662. [ Links ]
49. Harris WC. Portraits of the game and wild animals of Southern Africa delineated from life in their native haunts, during a hunting expedition from the Cape Colony as far as the Tropic of Capricorn, in 1836 and 1837, with sketches of the field sports. W Pickering: London; 1840. [ Links ]
 Correspondence to:
Correspondence to:
Andre Boshoff
Centre for African Conservation Ecology
Nelson Mandela Metropolitan University
Port Elizabeth 6031, South Africa
email: andre.boshoff@nmmu.ac.za
Received: 03 June 2009
Accepted: 11 Dec. 2009
Published: 11 Mar. 2010
This article is available at: http://www.sajs.co.za


{kind=link}
{kind=link}