










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.106 no.1-2 Pretoria ene./feb. 2010
REVIEW ARTICLE
A review of phytoplankton dynamics in tropical African lakes
Mzime R. Ndebele-MurisaI,III; Charles F. MusilII; Lincoln RaittIII
IDepartment of Biological Sciences, University of Zimbabwe, Harare, Zimbabwe
IIClimate Change and Bio-Adaptation Division, South African National Biodiversity Institute, South Africa
IIIDepartment of Biodiversity and Conservation Biology, University of the Western Cape, Cape Town, South Africa
ABSTRACT
This paper provides a synthesis of current knowledge on phytoplankton production, seasonality, and stratification in tropical African lakes and considers the effects of nutrient enrichment and the potential impacts of climate warming on phytoplankton production and composition. Tropical African lakes are especially sensitive to climate warming as they experience wide fluctuations in the thermocline over a narrow range of high water temperatures. Recent climate warming has reduced phytoplankton biomass and production in the lakes. A decline in the production of palatable chlorophytes and an increase in cyanobacteria has led to reduced zooplankton production and a consequent decline in fish stocks, all of which can be associated with the elevated water temperatures. This indicates that even moderate climate warming may destabilise phytoplankton dynamics in tropical African lakes, thereby reducing water quality and food resources for planktivorous fish, with consequent negative impacts on human livelihoods.
Keywords: phytoplankton; African lakes; changing environment; nutrient; habitat
INTRODUCTION
Tropical African lakes provide an important source of dietary protein to rural communities; potable water for humans, livestock and agriculture; a means of transportation of goods and generation of hydro-electric power; and a source of revenue from fishing and eco-tourism. Also, many of these lakes are havens for diverse endemic fish fauna of immense ecological and scientific importance that are dependent on sustained phytoplankton seasonality and stratification. Previous reports on phytoplankton in tropical African lakes have had a predominantly taxonomic focus, with some considering phytoplankton composition and production in specific lakes as a reflection of that lake's overall health. However, a comprehensive review of phytoplankton production, seasonality and stratification in all tropical African lakes, as well as phytoplankton responses to eutrophication and climate warming, and the consequent effect of this on fish stocks and human livelihoods, has not been undertaken. This paper seeks to address these issues, considering that such information is especially pertinent in the wake of the deteriorations in water quality and declines in fish abundance and diversity that have been recently experienced in many tropical African lakes, as a result of climate change and increased anthropogenic pressures.
PRIMARY PRODUCTION
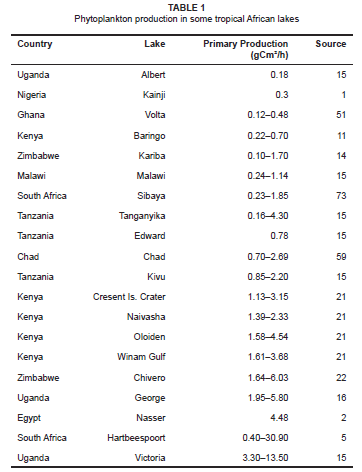
Tropical African lakes display a considerable range in phytoplankton production (see Table 1) which reflects their trophic state. Lake Kainji is a typical example of a nutrient-poor (oligotrophic) African lake with a low phytoplankton production of 0.3 gCm2/h.1 Towards the upper end of the production scale, is the nutrient-rich (eutrophic) Lake Nasser, whose phytoplankton production has been reported to average 4.48 gCm2/h.2 Increased nutrient inflows resulting from expanding urbanisation and agriculture have elevated primary production levels in many African lakes. A good example is Lake Chivero, whose primary production levels have increased by over 10 orders of magnitude from its previously reported range of 1.64 gCm2/h to 6.03 gCm2/h in 1979 to its current range of 18.5 gCm2/h to 140 gCm2/h.3,4 Similarly, in the physionomically similar Hartbeespoort Dam, primary production levels, which normally range between 0.40 gCm2/h and 30.90 gCm2/h, have risen to as high as 185 gCm2/h.5 These elevated primary production levels correspond with algal blooms and are a direct consequence of high nutrient inflows which overburden the lakes' natural purification systems.5,6 In deeper African lakes, nutrient inflows also exert prominent effects on primary production. Examples include Lake Malawi, where primary production levels of 0.24 gCm2/h and 1.14 gCm2/h have risen to between 14.04 gCm2/h and 26.20 gCm2/h in recent years7 and Lake Victoria where primary production of 3.30 gCm2/h to 13.50 gCm2/h has risen to as high as 234 gCm2/h.8 Primary production in tropical African lakes is often low at or near the water surface due to photo-inhibition. The optimal habitat for phytoplankton is in the first 3 m - 5 m of the water column. Production tends to decrease with depth, until light becomes the limiting factor after the euphotic depth. Light is the primary limiting factor to the growth of phytoplankton in Lake Chivero,9 whereas conductivity and trophic status (measured as chlorophyll a) are the most important environmental variables influencing the distribution of ciliate species in this and other African lakes.10 These variables are compliant with a classification of 17 east African lakes on the basis of water conductivity and associated phytoplankton species.11
STRATIFICATION
Environmental factors such as water pH, conductivity, dissolved oxygen and nutrient concentration and light intensity influence primary production and these in turn are affected by thermal stratification, which is a common feature of tropical African lakes. Stratification is a result of thermal differences in the upper warm layer of water, (the epilimnion), the lower cold layer (the hypolimnion), and a zone of steep temperature gradient, (the thermocline), which separates the two and is usually created during periods of high summer temperatures. Stratification is characterised by vertical profiles of water temperature, conductivity, dissolved oxygen concentration and pH. In Lake Tanganyika, a fairly stable stratification is established during the wet season, which is generated by increased water temperatures and diminished wind intensities. However, currents and internal waves, as well as coastal jets and return flows, do cause localised up-welling, which may partly disrupt this stratification.12 Stratification is also prevalent in many deep tropical African lakes, such as Albert, Cabora Bassa, Chad, Kariba, Malawi, Niger and Turkana. This often influences the seasonality and abundance of phytoplankton.
The general annual cycle of thermal stratification in these lakes includes a short phase of partial vertical mixing which is often accompanied, or immediately followed, by a peak of algal abundance to which diatoms make a major contribution.12 A second peak, dominated by blue-green algae, often develops after re-stratification.12 In Lake Ogelube in Nigeria, phytoplankton biomass during the rainy season (April-October) is much higher than that during the dry season (November-March). Chlorophyceae (mostly desmids) are most abundant during the rainy season, followed (in decreasing order of abundance) by Cyanophyceae, Bacillariophyceae, Euglenophyceae, Dinophyceae, Cryptophyceae, Chrysophyceae and Xanthophyceae.13 This order changes slightly in the dry season, when the relative abundance of Bacillariophyceae is greater than that of Cyanophyceae, and Dinophyceae is greater than that of Euglenophyceae. A similar seasonal pattern is found in other African lakes, such as Kariba, Malawi, Tanganyika and Victoria, where blue-green algae dominate during summer stratification and diatoms tend to dominate in winter at turn-over, when stratification breaks down.14,15
Density stratification is also a natural feature of many shallow, naturally saline African lakes such as George16,17, Naivasha18and Nakuru19. However, this stratification can be disrupted by strong winds which influence water circulation, thereby preventing the development of a consistent thermocline.20 This feature has been observed in the four Kenyan lakes: Naivasha, Crescent Island Crater, Oloiden Lake and Winam Gulf, which displayed complete vertical mixing and well-oxygenated water throughout the water profile during periods of strong winds.21Stratification in some shallow African lakes also traps nutrients from riverine inflows contaminated by effluent discharges. This is apparent in Lake Chivero during hot summer months,22 with stratification breakdown during winter months resulting in the temporary upwelling of nutrients and localised algal blooms.6
Stratification generally has a negative impact on overall primary production as it confines nutrients to deeper waters of greater density beyond the euphotic zone,23 with phytoplankton species diversity and total biomass elevated only in nutrient rich waters at river inflows.24 As a result, the highest primary production normally occurs in the first 3 m of the water column, and is often inversely correlated with water transparency (measured using a Secchi disc). This is usually depressed at the water surface, particularly where temperatures exceed 30 ºC.3 In most African lakes, strong thermal gradients often develop from daytime warming in the 0 m - 5 m layer. These gradients frequently trap photosynthetic oxygen to form well-defined maxima which demarcate deeper, oxygen poor layers.25 In some lakes, the de-oxygenation goes so far that oxygen disappears from the hypolimnion and hydrogen sulphide is produced. The epilimnion maintains a high oxygen concentration throughout the year, except during partial stratification, which may develop during the day and cause a gradient of decreasing oxygen concentration with depth. Some features of stratification in artificial African lakes are also influenced by the positioning of turbines. For instance, Lake Kainji's rapid deepening of the epilimnion during stratification has been attributed to discharges from the lake as the levels of the spillways and turbine intakes are situated below the thermocline.26
SEASONALITY
Most African lakes have a well-established seasonality in phytoplankton abundance, which is governed mainly by climate. The community of larger phytoplankton in Lake Tanganyika is mainly composed of chlorophytes and diatoms (Nitzschia spp.), with large blooms of diastrophic filamentous cyanobacteria also periodically observed during September to November at the onset of the rainy season.27,28 However, more recently, these cyanobacteria have not been observed as a major constituent of the phytoplankton biomass.29,30 Rather the biomass exhibits contrasting picocyanobacteria and diatom concentrations due to changes in water column stability also observed at the southern and northern ends of the lake.31,32 A seasonal succession of major algal groups similar to that in Lake Tanganyika is also found in the natural oligotrophic Lakes Edward, Kivu and Malawi, which have a low phytoplankton biomass and species diversity,15 as well as in the artificial, oligotrophic Lake Kariba, where cyanobacteria (Cylindrospermopsis, Anabaena, Pseudoanabaena) and blue-green algae dominate when the phytoplankton biomass reaches its peak (up to 1.52 mg per litre of chlorophyll a) during the rainy season, with diatoms dominating at turnover during the dry season.33 Similarly, diatoms are the dominant group in Lake Kivu during episodes of deep mixing in the dry season, with filamentous, diastrophic cyanobacteria and picocyanobacteria forming a substantial fraction of the autotrophic biomass under conditions of reduced nutrient availability in the stratified water column during the rainy season.34 Lake Kivu's phytoplankton species assemblage is somewhat intermediate in composition, between the oligotrophic Lakes Malawi and Tanganyika and the more eutrophic Lake Victoria, though the dominant diatoms of Lake Kivu (Urosolenia sp., Nitzschia bacata, Fragilaria danica) are normally associated with oligotrophic, phosphorous-deficient African lakes.34
The concept of stable phytoplankton communities has been questioned with regards to some African lakes. Observations of primary production and phytoplankton densities at regular (weekly and fortnightly) intervals in Lake Kinneret, Israel, over a 35-year period (1969-2004), has led to a repudiation of the concept of a stable phytoplankton seasonality.35 These observations demonstrated remarkable stability in the composition and abundance of algal species in summer and spring algal blooms in the lake during the first 24 years of monitoring. After 1994, however, deviations from the previously predictable annual pattern of algal blooms were apparent. They were characterised by the absence of prevailing spring Peridium gatunense blooms, intensification of winter Melosira granulata (=Aulacoseira) blooms and increased variability in the magnitude of blooms in other species. Also, nanoplanktonic palatable forms in the high summer phytoplankton biomass were replaced by less palatable forms. There was a new appearance and increase in absolute biomass of toxin-producing, nitrogen-fixing cyanobacteria as well as an increased incidence of fungi attacking Peridium gatunense.35 In Lake Tanganyika, shifts in phytoplankton seasonality have also been detected through the application of Empirical Orthogonal Function Analysis to a seven-year satellite observation record.36 Similarly, the phytoplankton composition in Lake Kariba between 1959 and 1964 has been compared with that between 1986 and 1990 and showed a change from a riverine-dominated species component rich in desmid flora and large algal species, to that dominated by small-celled chlorococcal green algae. This change was mainly a result of the apparent stabilisation of the lake since its creation.14
THE INFLUENCE OF NUTRIENTS
In the tropics and at mid-latitudes, phytoplankton are typically nutrient-limited; and reduced biological productivity, phytoplankton biomass and growth have been linked to increased water temperatures and reduced nutrient supply.36Nitrogen and phosphorous have been identified as the major nutrients governing primary production and phytoplankton biomass in tropical African lakes.11,37 Indeed, in many African lakes nitrogen-fixing taxa such as Anabaena and Cylindrospermum proliferate due to the limiting concentrations of this nutrient for other taxa in their waters.11,12,18,30,38 The low phytoplankton biomass variability of Lake George in Uganda has been attributed to both nitrogen and phosphorous limitations, and the extreme predominance of algal production over that of other biota.38 Similarly, in Lake Kariba it has been demonstrated that the waters are principally phosphorous limited for most of the year with a possibility of nitrogen co-limiting at other times.39 In this case, the ability to fix nitrogen becomes an added advantage and may explain the dominance of nitrogen-fixing taxa. In Lake Naivasha, nitrogen is more limiting than phosphorous, with an algal preference for ammonium over nitrate.18 In Lake Tanganyika, the addition of nitrogen, phosphorous and iron stimulates total phytoplankton production. This finding was based on both high-performance liquid chromatography (HPLC) pigment analysis and cellular counts, using epifluorescence microscopy.30 Iron enhances production of mainly prokaryotic pico-phytoplankton, nitrogen and phosphorous increase green algae and, in some cases, diatom production and iron in combination with nitrogen and phosphorous improves picocyanobacteria production.30 Silica is known to increase diatom production because of their siliceous shells and this has been demonstrated in mesocosm experiments.30
The composition and abundance of phytoplankton often reflect the nutrient status of African lakes with, for example, the low Euglenophyceae biomass in Lake Ogelube proposed to be indicative of low organic pollution and the predominance of desmids indicative of oligotrophic conditions.13 Low nutrient contents have also been observed in the normally oligotrophic Lakes Kariba, Kivu and Malawi. Despite this, many African lakes are now prone to nutrient enrichment from anthropogenic activities and this has led to eutrophication and associated problems. Classical examples of eutrophic African water bodies are Lakes Chivero and Victoria and the Hartbeespoort Dam, with recorded nitrogen concentrations of over 50 mg/L. Elevated nutrient levels in Lake Chivero, caused by discharge of partially treated sewage and industrial effluent into the lake, has resulted in an increase in phytoplankton biomass by a factor of more than ten and corresponding reductions in water transparency.6 Associated with these changes is a progressive decrease in phytoplankton species diversity, with the cyanobacterial species Microcystis aeruginosa dominating and Melosira spp. co-dominating the phytoplankton community.4,6 Cyanobacterial blooms have also been reported in nutrient-enriched South African impoundments, namely the Hartbeespoort Dam40 and the Erfenis and Allemanskraal Dams.41 Similarly, nutrient-rich inflows into a shallow tropical lake in the Cameroon (Yaoundé Municipal Lake) have resulted in a massively increased phytoplankton biomass of 225 µg/mL and chlorophyll a concentration of 566 mg/m3, dominated by Euglenophyta and Chlorophyta.42 Eutrophication tends to affect shallow lake systems more than deeper tropical lakes, such as Lakes Malawi and Victoria, with incidences of localised pollution only occasionally reported in the deeper lakes such as Kariba.43 In fact, in Lake Kariba the water has changed from an initial nutrient-rich (eutrophic) state to that of its current nutrient-poor (oligotrophic) state, with phosphorous limiting primary production.39 However, the high concentrations of faecal coliform bacteria recorded along the northern shoreline of Lake Kariba44 indicated that the lake waters are verging towards mesotrophy. The initial eutrophic state in the lake has led, in the past, to the proliferation of the noxious aquatic weed Salvinia molesta (Kariba weed) and, more recently, to the propagation of another aquatic weed, Eichhornia crassipes (water hyacinth), which is currently prevalent in Lakes Chivero and Victoria, as well as in Lakes Baringo, Chad, Malawi, Naivasha, Nakuru, Tanganyika and Victoria, where it is adversely affecting fishing industries.45
CLIMATE WARMING
The impact of global warming projected by climate change models46,47,48 on the function of African aquatic ecosystems is uncertain, although some likely effects have been reported49. Inter-annual variations in phytoplankton composition and biomass in temperate regions are known to reflect changes in climate.50 Reduced depths of light transparent epilimnions, due to upward shifts in thermoclines in response to elevated temperatures, could potentially adversely affect primary production in deep African lakes such as Kariba, Malawi, Tanganyika and Victoria. However, relatively limited data exist on the influence of climate variability on phytoplankton biomass and composition in tropical African lakes. Only a few phytoplankton studies, which do not specifically address climate-driven changes, have been undertaken in Africa, such as in Lake Volta51 and in Lakes Malawi, Tanganyika and Victoria52. However, some tendencies have been noted.49,53 East African lakes, for example, are known to be potentially highly sensitive to climate change54 because small variations in climate cause wide fluctuations in the thermocline over a narrow range of high water temperature.55 Indeed, thermal stratification, which isolates nutrients from the euphotic zone is strongly linked to hydrodynamic and climatic conditions in Lake Tanganyika.56 Also, an analysis of climatic data for the middle Zambezi valley shows that warming around Lake Kariba is proceeding at a faster rate than regional models have predicted, which points to future shifts in phytoplankton species richness and production.57 In Lake Tanganyika, it was reported that regional climate change over the past 80 years has reduced productivity and, more specifically, primary production.53 However, other climatic factors such as the South-East trade winds also impact the primary production in this lake by increasing wave action and upwelling, which brings nutrients from deeper regions of the lake to the surface. Conversely, during less windy El Niño years the lake water column is more stable, resulting in enhanced nutrient depletion in surface waters and diminished phytoplankton production.53,58 These findings were corroborated by another study that demonstrated a close correspondence between phytoplankton biomass and climate in Lake Tanganyika.36 This seven-year study revealed that shifts in phytoplankton dominance were due to the lake's high sensitivity to climate change, as variations in wind and air temperatures favoured the release or entrapment of nutrients present in deeper waters.
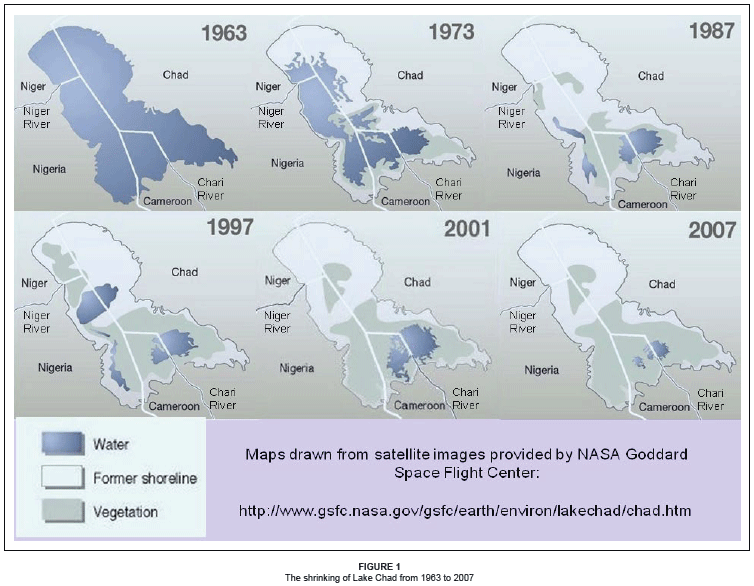
Much of what is happening to lakes across Africa is attributable to years of drought that have reduced river inflows and rising temperatures and caused increased evaporative water loss.48 An extreme example of this is Lake Chad. Once the world's sixth-largest lake, it has since decreased to 5% of its original size of approximately 25 000 km2 in 1963, to 1 350 km2 in 2001 (Figure 1). This decrease can also be attributed to large and unsustainable irrigation projects built by Niger, Nigeria, Cameroon and Chad, which have diverted water from the lake as well as the Chari, Logone and Niger Rivers, and major overgrazing in the region resulting in a loss of vegetation and serious deforestation contributing to a drier climate.59,60 Another example of an African lake that may be responding to increasing temperatures driven by climate change is Lake Turkana, the largest, most northerly and most saline of Africa's Rift Valley lakes, whose level has dropped 10 m between 1975 and 1992 due to reduced inflow.61 Similarly, Lake Victoria's level has dropped over 1.5 m in five years.62,63 For the Zambezi basin, simulated runoff under climate change is projected to decrease by about 40% or more, which would greatly reduce the size and depth of Lakes Cabora Bassa, Kafue and Kariba.64 In the deep, meromictic Lake Tanganyika, warming has increased water column stability and decreased vertical mixing, thereby reducing nutrient loading to the upper mixed layer.53 Since 1974, observations in this lake have documented decreases in phytoplankton abundance and increases in dissolved silica, both indicative of declining primary production, with negative implications for the pelagic fishery which accounts for more than 90% of all landings.53,58 However, fishing effort continues to rise and technologies have improved to such an extent that landings have increased during the warming period, making it difficult to decipher any effect on fish productivity in response to warming. In Lake Victoria, climate warming has been implicated in the eutrophication of the world's largest lake fishery.65 However, increased phosphorus loading is more likely the direct cause, with the climate probably playing a smaller role by increasing stability of seasonal stratification and accelerating the onset of hypolimnetic deoxygenation.65 Consequences of lake warming for the hundreds of endemic cichlid fishes, especially those occurring in littoral areas of the lake, are speculative.63
Increased water temperatures associated with climate warming are likely to cause a shift in phytoplankton species composition from Chlorophyceae to Cyanophyceae, which are competitively superior at higher temperatures. Microcosm studies have demonstrated that elevated temperatures suppress total zooplankton biomass by altering phytoplankton community composition towards high temperature tolerant species, though total phytoplankton biomass is not usually altered.66 For the phytoplankton community of Lake Chivero, it was found that the water temperature at which the blue-green algae (Cyanophyceae) dominated cultures was between 25 ºC and 26 ºC, whereas for green algae (Chlorophyceae) it was between 21 ºC and 23 ºC.67 These findings may explain the low biomass of the previously dominant Chlorophyceae in Lake Kariba where 73% of the temperatures recorded at a depth of 5 m exceeded 24 ºC.57 Since its creation, Lake Kariba has exhibited distinct changes in water chemistry and thermal properties, the latter seemingly due to global warming, as the rate of warming in the Zambezi valley has nearly doubled the IPCC global average of 0.2 ºC per decade.57 This change in the lake's thermal properties is reflected in an upward migration of the thermocline, apparent from the smaller metalimnion temperature gradient and an upward movement of the low (> 2 mg/L DO) oxygen layer. These changes in thermal properties ostensibly correspond with declines in Limnothrissa fish populations.49
As water temperatures increase, algal succession follows a progression - from diatoms to chlorophytes to cyanobacteria.33 In Lake Victoria, the phytoplankton blooms observed during February and August comprised over 90% Microcystis at concentrations of 34 000 colonies per millilitre (77.6 mg/L), the blooms corresponding with high temperatures, discharge of nutrients from river inflows, nutrient upwelling and nutrient release from sediments.68 The preponderance of cyanobacteria at higher water temperatures (> 25 ºC) has led to concerns that increased water temperatures due to global warming could result in a decline in the production of palatable chlorophytes, leading to decreased zooplankton production and a consequent decline in fish stocks. Indeed, fisheries data from Lake Tanganyika show significant correlations with climatic (ENSO) data over the last 40 years.58 This suggests that moderate warming could destabilise plankton dynamics, thereby potentially reducing water quality and food resources for higher trophic levels such as planktivorous fish as seen in shallow cold-water ecosystems.69 Also, oxygen depletion and release of toxins caused by algal bloom die-off may lead to massive fish mortalities, as reported in Lake Chivero.6,70 In this lake, the dissolved oxygen concentrations following an algal bloom die-off, ranged from 2 mg/L at a depth of 5 m to 3.9 mg/L at the lake surface. These oxygen concentrations were even lower than those (3.2 mg/L to 4.8 mg/L) measured in the Nyanza Gulf of Lake Victoria, following the collapse of a phytoplankton bloom and associated with high fish mortalities of Oreochromis niloticus.68 Algal toxins caused by algal bloom die-off have also been previously detected in Lake Chivero and in tap water emanating from the lake,6,22 but it has not been established whether these are toxic to humans and other fauna.70 However, toxic strains of Microcystis have been identified in other African fresh water bodies, as well as in fresh water systems elsewhere in the world.71
HUMAN LIVELIHOODS
The future of tropical African lakes in a changing climate seems bleak and this poses several challenges, the most important of which is an understanding of how changes in climate will affect aquatic ecosystems and human livelihoods, need to be overcome.47,49 The consequences of climate warming on lake hydrology (through destabilisation of stratification) and on lake temperature, could affect phytoplankton production by reducing both abundance and composition in many tropical African lakes. The consequent cascade through a longer trophic chain due to the dominance of smaller phytoplankton, could potentially affect zooplankton and fish production and ultimately threaten the viable fishery industries that sustain the livelihoods of riparian communities. Unfortunately, the poor are most at risk because they depend on the natural resources provided by many African lakes, yet policies around these resources do not benefit them much. For instance, upon Lake Kariba's creation, 57 000 people were resettled to make way for the lake, but suitable land was not sufficiently available in the direct vicinity.72 These people should presently be reaping the benefits of the artificial lake, but there is a disparity in the allocation of resources and policies that govern the use of the lake. The value of African lakes is without question, for they contribute significantly to poverty reduction and food security. They are a source of cheap dietary proteins, with lakes such as Chilwa, Kariba, Malawi, Tanganyika and Victoria contributing more than 60% of the dietary protein to bordering rural communities. The lakes also provide potable water for human and livestock use, irrigation water for agriculture, are a means of transportation of goods and generate hydro-electric power and are a source of revenue from fishing and eco-tourism. Lakes Malawi, Tanganyika and Victoria, particularly, are a haven of diverse endemic fish fauna of immense ecological and scientific importance. Generating deep concern are the declines in fish abundance and diversity and the deterioration in water quality experienced by many tropical African lakes. Although policies have been implemented to mitigate these threats, there remains an urgent need to consistently monitor the health of tropical African lakes to circumvent adverse effects caused by anthropogenic pressures and a changing climate.45
REFERENCES
1. Karlman SG. The annual flood regime as a regulatory mechanism for phytoplankton production in Kainji Lake, Nigeria. Arch. Hydr. 1982;86:93-97. [ Links ]
2. Samaan AA. Report on the trip of Lake Nasser to investigate its primary productivity during March 1971. Report to Lake Nasser Development Centre Project, Aswan (mimeo). [ Links ]
3. Robarts RD. Underwater light penetration, chlorophyll a and primary production in a tropical African lake (Lake Mcllwaine, Rhodesia). Arch Hydrobiol. 1979;86:423-444. [ Links ]
4. Ndebele MR. Some aspects of the phytoplankton community of Lake Chivero, Harare, Zimbabwe. MSc. thesis, University of Zimbabwe, Harare; 2003. [ Links ]
5. Robarts RD. Factors controlling primary productivity in a hypereutrophic lake (Hartbeespoort Dam, South Africa). J Plankton Res. 1984;6:91-105. [ Links ]
6. Mhlanga L, Day J, Cronberg C, Chimbari M, Siziba N, Annadotter H. Observations on limnological conditions associated with a fish kill of Oreochromis niloticus in Lake Chivero following collapse of an algal bloom. Afr J Ecol. 2006;44:199-208. [ Links ]
7. Guildford SJ, Bootsman HA, Taylor WD, Hecky RE. High variability of phytoplankton photosynthesis in response to environmental forcing in an oligotrophic Lake Malawi/Nyasa. J Great Lakes Res. 2007;33:170-185. [ Links ]
8. Mugidde R. The increase in phytoplankton activity in Lake Victoria (Uganda). Verh. Int. Ver. Limnol. 1993; 25:846-849. [ Links ]
9. Robarts RD, Southall GC. Nutrient limitation of phytoplankton growth in seven tropical man-made lakes, with special reference to Lake Mcllwaine, Rhodesia. Arch Hydrobiol. 1977;79:1-35. [ Links ]
10. Yasindi AW, Taylor WD. The abundance, biomass and composition of pelagic ciliates in East African lakes of different salinity and trophy. Aquat Ecosyst Health Manage. 2003;6:289-297. [ Links ]
11. Talling JF, Talling IB. The chemical composition of African lake waters. Int Rev Gesamten Hydrobiol. 1965;50:421-463. [ Links ]
12. Chale FMM. Inorganic nutrient concentrations and chlorophyll in the euphotic zone of Lake Tanganyika. Hydrobiologia 2004;00:1-9. [ Links ]
13. Nweze NO. Seasonal variations in phytoplankton populations in Ogelube Lake, a small natural West African Lake. Lakes Reserv Res Manage. 2006;11:63-72. [ Links ]
14. Cronberg G. Phytoplankton in Lake Kariba. In: Moreau J, editor. Advances in the ecology of lake Kariba. Harare: University of Zimbabwe Publications, 1997; p.66-72 [ Links ]
15. Hecky RE, Kling HJ. Phytoplankton ecology of the great lakes in the rift valleys of Central Africa. Arch Hydrobiol. Beiheft Ergebnisse Limnol. 1987;25:197-228. [ Links ]
16. Ganf GG. Photosynthetic production and irradiance-photosynthesis relationship of the phytoplankton from a shallow equatorial lake (Lake George, Uganda). Oceologia (Berlin) 1975;18:165-183. [ Links ]
17. Wood RB, Prosser MV, Baxter RM. The seasonal pattern of thermal characteristics of four of the Bishoftu crater lakes, Ethiopia. Freshwat Biol. 1976;6:519-530. [ Links ]
18. Hubble DS, Harper DM. Nutrient control of phytoplankton production in Lake Naivasha, Kenya. Hydrobiologia. 2002;488: 99-105. [ Links ]
19. Vareschi E. The ecology of Lake Nakuru (Kenya). III. Abiotic factors and primary production. Oceologia. 1982;55:81-101. [ Links ]
20. Langenberg VT, Mwape LM, Tshibangu K, et al. Comparison of thermal stratification, light attenuation and chlorophyll-a dynamics between the ends of Lake Tanganyika. Aquat Ecosyst Health Manage. 2002;5:255-265. [ Links ]
21. Melack JM. Photosynthetic rates in four tropical African fresh waters. Freshw Biol. 1979;9:555-571. [ Links ]
22. Ndebele MR, Magadza CHD. The occurrence of Microcystin-LR in Lake Chivero, Zimbabwe. Lakes Reserv.: Res Manage. 2006;11:57-62. [ Links ]
23. Spigel RH, Coulter GW. Comparison of hydrology and physical limnology of the East African Great Lakes: Tanganyika, Malawi, Victoria, Kivu and Turkana (with reference to some North American Great Lakes). In: Johnson TC, Odada E. The limnology, climatology and paleoclimatology of the east African lakes. Toronto: Gordon and Breach; 1996. Toronto: Gordon and Breach; 1996. [ Links ]
24. Vuorio K, Nuottajarvi M, Kalevi S, Sarvala J. Spatial distribution of phytoplankton and picocyanobacteria in Lake Tanganyika in March and April 1998. Aquat Ecosyst Health Manage. 2003;6:263-278. [ Links ]
25. Talling JF, Wood RB, Prosser MV, Baxter RM. The upper limit of photosynthetic productivity by phytoplankton: evidence from Ethiopian Soda lakes. Freshwat Biol. 1973;3:5376. [ Links ]
26. Henderson F. A limnological description of Kainji Lake 1969-1971. FAO repository report prepared for the Kainji Lake Project FI: DP/NIR 66/524/10, Rome; 1973. [ Links ]
27. Salonen K, Sarvala J, Jarvinen M, et al. Phytoplankton in LakeTanganyika- vertical and horizontal distribution of in vivo fluorescence. Hydrobiologia. 1999;407:89-103. [ Links ]
28. Cocquyt C, Vyverman W. Phytoplankton in Lake Tanganyika: a comparison of community composition and biomass off Kigoma with previous studies 27 years ago. J Great Lakes Res. 2005;31:535-546. [ Links ]
29. Descy JP, Hardy MA, Stenuite S, et al. Phytoplankton pigments and community composition in Lake Tanganyika. Freshw Biol. 2005;50:668-684. [ Links ]
30. De Wever A, Muylaert K, Langlet D, et al. Differential response of phytoplankton to additions of nitrogen, phosphorus and iron in Lake Tanganyika. Freshw Biol. 2008;53:264-277. [ Links ]
31. Hecky RE., Kling HJ. The phytoplankton and proto-zooplankton of the euphotic zone of Lake Tanganyika- species composition, biomass, chlorophyll content, and spatio-temporal distribution. Limnol Oceanogr. 1981;26:548-564. [ Links ]
32. Vuorio K, Nuottajarvi M, Kalevi S, Sarvala J. Spatial distribution of phytoplankton and picocyanobacteria in Lake Tanganyika in March and April 1998. Aquat Ecosyst Health Manage. 2003;6:263-278. [ Links ]
33. Ramberg L. Phytoplankton succession in the Sanyati basin, Lake Kariba. Hydrobiologia 1987;153:193-202. [ Links ]
34. Sarmento H, Isumbisho M, Descy JP. Phytoplankton ecology of Lake Kivu (eastern Africa). J Plankton Res. 2006;28:815-829. [ Links ]
35. Zohary T. Changes to the phytoplankton assemblage of Lake Kinneret after decades of a predictable, repetitive pattern. Freshw Biol. 2004;49:1355-1371. [ Links ]
36. Bergamino N, Loisellea SA, Cozar A, Dattiloa AM, Bracchini L, Rossi C. Examining the dynamics of phytoplankton biomass in Lake Tanganyika using empirical orthogonal functions. Ecolog. 2007 Model. 204,156-162. [ Links ]
37. Viner AB. Relationships of nitrogen and phosphorus to a tropical phytoplankton population. Hydrobiologia. 1977;52:185-196. [ Links ]
38. Ganf GG. Photosynthetic production and irradiance-photosynthesis relationship of the phytoplankton from a shallow equatorial lake (Lake George, Uganda). Oceologia. 1975;18:165-183. [ Links ]
39. Moyo S.M. Cyanobacterial nitrogen fixation in Lake Kariba, Zimbabwe. Verh Int Ver Limnol. 1991;24:1123-1127. [ Links ]
40. Robarts RD, Zohary T. The influence of a cyanobacterial hyperscum on the heterotrophic activity of planktonic bacteria in a hypereutrophic lake. Appl Environ Microbiol. 1986;51:609-613. [ Links ]
41. Van Ginkel CE, Hohls BC. Toxic algae in Erfenis and Allemanskraal dams. Occasional report by the Institute for Quality Studies, Department of Water Affairs and Forestry. N/C400/03/DEO/1999. [ Links ]
42. Kemka N, Njine T, Togouet SHZ, et al. Eutrophication of lakes in urbanized areas: The case of Yaounde Municipal Lake in Cameroon, Central Africa. Lakes Reserv Res Manage. 2006;11:47-55. [ Links ]
43. Feresu SB, Van Sickle J. Coliforms as a measure of sewage contamination of the River Zambezi. J Appl Bacteriol. 1990;68:397-403. [ Links ]
44. Magadza CHD, Dhlomo EJ. Wet seasons coincidence of coliform bacteria in inshore waters of Kariba town area. Lakes Reserv Res Manage. 1996;2:89-96. [ Links ]
45. Ogutu-Ohwayo R, Balirwa JS. Management challenges of freshwater fisheries in Africa. Lakes Reserv. Res Manage. 2006;11:215-226. [ Links ]
46. Hulme M. Climate change and southern Africa: an exploration of some potential impacts and implications in the SADC region. WWF International, Avenue du Mont Blanc, Gland, Switzerland; 1996. [ Links ]
47. Hulme M, Doherty R, Ngara T, New M, Lister D. African climate change: 1900-2100. Climate Res. 2001;17:145-168. [ Links ]
48. Intergovernmental Panel on Climate Change (IPCC). Freshwater Resources and Their Management. A Special Report of Working Group II. Fourth Assessment Report on Climate Change. Cambridge University Press, Cambridge, United Kingdom; 2007. [ Links ]
49. Magadza CHD. Climate change: some multiple impacts in Southern Africa. In: Downing T-E, editor. Climate change and food security. NATO ASI Series. Series 1: Global Environmental Change. 1994;37:662. [ Links ]
50. Lehman JT, Litt AH, Mugidde R, Lehman DA. In: Lehman JT, editor. Environmental change and response in east African lakes. Amsterdam: Kluwer. [ Links ]
51. John DM. The inland waters of tropical West Africa. An introduction and botanical review. Arch Hydrobiol Ergebneisse Limnol. 1986;23:1-244. [ Links ]
52. Cocquyt C, Vyverman W. Composition and diversity of the algal flora in the East African great lakes: a comparative survey of lakes Tanganyika, Malawi (Nyasa), and Victoria. Arch. Hydrobiol. Beiheft Ergebnisse Limnol. 1994;44:161-172. [ Links ]
53. O'Reilly CM, Alin SR, Plisnier PD, Cohen AS, MaKee BA. Climate change decreases aquatic ecosystem productivity of Lake Tanganyika, Africa. Nature. 2003;424:766-768. [ Links ]
54. Johnson TC, Odada EO. The limnology, climatology and paleoclimatology of the East African Lakes. Amsterdam: Gordon and Breach; 1996. [ Links ]
55. Lewis MW. Primary production in the plankton community of a tropical lake. Ecolog Monogr. 1974;44:377-409. [ Links ]
56. Plisnier PD, Chitamwebwa D, Mwape L, Tshibangu K, Langenberg V, Coenen E. Limnological annual cycle inferred from physical-chemical fluctuations at three stations of Lake Tanganyika. Hydrobiologia. 1999;407:45-58. [ Links ]
57. Magadza CHD. State of Environment on Lake Kariba and the Zambezi valley. Lakes Reserv. Res. Manage. In press; 2009. [ Links ]
58. Stenuite S, Pirlot S, Hardy MA, Sarmento H, Tarbe A, LB, Leporcq B, Descy JP. Phytoplankton production and growth rate in Lake Tanganyika: evidence of a decline in primary productivity in recent decades. Freshw Biol. 2007;52:2226-2239. [ Links ]
59. Carmouze JP, Durand JR, Lévêque C. Lake Chad. Ecology and productivity of a shallow tropical ecosystem. Monogr Biol. 1983;53. [ Links ]
60. Coe MT, Foley JA. Human and natural impacts on the water resources of the Lake Chad basin. J. Geophysical Res. (Atmospheres) 2001;106:3349-3356. [ Links ]
61. McGinley M. Lake Turkana National Parks, Kenya. In Encyclopaedia of earth.Washington DC: Environmental Information Coalition, National Council for Science and the Environment; 2008. [ Links ]
62. Awange JL, Sharifi MA, Ogonda G, Wickert J, Grafarend EW, Omulo MA. The falling Lake Victoria water level: GRACE, TRIMM and CHAMP satellite analysis of the Lake Basin. Water Resour Manage. 2008;22:775-796. [ Links ]
63. Phoon SY, Shamseldin AY, Vairavamoorthym K. Assessing impacts of climate change on Lake Victoria Basin, Africa. In proc. Of the 30th WEDC International Conference: People-centred approaches to water and environmental sanitation. Vientiane, Lao PDR. 2004. [ Links ]
64. Desanker P, Magadza CHD. In: McCarthy JJ, Canziani O-F, Leary N, Docken DJK, White S, editors. Climate change 2001: Impacts and adaptations. CUP; 2001. [ Links ]
65. Fryer G. Biological implications of a suggested late Pleistocene desiccation of Lake Victoria. Hydrobiol. 1997;354:177-182. [ Links ]
66. Doney SC. Plankton in a warmer world. Nature. 2006;444:695-696. [ Links ]
67. Sibanda P. The possible effects of global warming on the growth of algae. B.Sc. thesis, University of Zimbabwe, Harare, Zimbabwe; 2003. [ Links ]
68. Ochumba OP, Kibaara DI. Observations on blue-green algal blooms in the open waters of Lake Victoria, Kenya Afri J Ecol. 1989;27:23-34. [ Links ]
69. Strecker AL, Cobb TP, Vinebrooke RD. Effects of experimental greenhouse warming on phytoplankton and zooplankton communities in fishless alpine ponds. Limnol and Oceanogr. 2004;49:1182-1190. [ Links ]
70. Moyo NAG. The causes of massive fish deaths in Lake Chivero. In: Moyo NAG, editor. Lake Chivero a polluted lake. Harare: University of Harare. [ Links ]
71. Chen J, Xie P, Xu J. First identification of the hepatotoxic microcystin in the serum of a chronically exposed human population together with indication of hepatocellular damage. Toxicol Science 2009;108:81-89. [ Links ]
72. Scudder T. Man-made lakes and population resettlement in Africa. In: Lowe-McConnell RH, editor. Man-made lakes. New York: Academic Press; 1966. [ Links ]
 Correspondence to:
Correspondence to:
Mzime Ndebele-Murisa
Department of Biodiversity and Conservation Biology
University of the Western Cape, Box X17
Bellville 7535, Cape Town, South Africa
email: murisa.mzime@gmail.com
Received: 07 Oct. 2009
Accepted: 11 Dec. 2009
Published: 11 Mar. 2010
This article is available at: http://www.sajs.co.za


{kind=link}