










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.105 no.3-4 Pretoria mar./abr. 2009
RESEARCH LETTERS
A lovebird (Psittaciformes: Agapornis) from the Plio-Pleistocene Kromdraai B locality, South Africa
T.A. Stidham
Department of Biology, Texas A&M University, 3258 TAMU, College Station, Texas 77843-3258, U.S.A. E-mail: furcula@mail.bio.tamu.edu
ABSTRACT
The previous report of the presence of lovebirds (Agapornis) at the Plio-Pleistocene Kromdraai B fossil locality is confirmed by the identification of a humerus from Member 3. That fossil specimen differs from extant lovebird species in morphology, and its slightly smaller size indicates that it likely represents an extinct taxon. This lovebird specimen is likely from the early Pleistocene (younger than 1.95 Myr) and appears to have been deposited around the same time as the fossil hominid specimens that have been uncovered at the site. The modern preference of lovebirds for a wide variety of wooded and forested habitats adds support to the mammalian-based idea that the palaeohabitat around Kromdraai B where Australopithecus robustus lived in the early Pleistocene was a mosaic that included wooded or forested habitats.
Key words: parrot, lovebird, Kromdraai B, hominid
Introduction
The fossil record of parrots in Africa is poorly known and is mostly represented by undescribed material from the Pliocene Olduvai Bed I, Tanzania,1 Miocene Varswater Formation, South Africa,2 and Plio-Pleistocene Kromdraai B, South Africa.3 The sole illustrated and described specimen of a fossil parrot in Africa is a lovebird (Agapornis) quadrate from the external Pleistocene deposits (~1 Myr) at Plovers Lake, South Africa.4 Lovebirds, genus Agapornis, comprise nine extant species of small-bodied, colourful parrots that occur only in Africa. The only other discussion of the fossil record of parrots from hominid-bearing cave deposits in Gauteng Province, South Africa, is the reported presence of lovebirds at Kromdraai B.3 In that publication, the parrot specimens were not described or illustrated, and the method of their identification was not mentioned. Pocock3 allocated the Kromdraai B material to two different lovebird species (one larger and one smaller). The specimens that were the focus of that study have since been lost.4 Although a diminutive lovebird specimen has been described from Plovers Lake,4 the evidence for the presence of Agapornis at Kromdraai B is still tentative. Fortunately, a recently identified humerus from the Transvaal Museum Kromdraai B collections appears to represent a small species of lovebird and confirms the presence of that taxon in the early Pleistocene of South Africa.
The site
Kromdraai B is a set of cave breccia deposits in the Bloubank River Valley approximately 2 km from Sterkfontein in Gauteng Province, South Africa. Those sediments have produced a diverse vertebrate fauna, including multiple specimens of the hominid Australopithecus (Paranthropus) robustus.5 Many workers have described the details of the geology and stratigraphy of the calcified and decalcified cave breccias at Kromdraai B and divided the deposits into various members.6–11 Analysis of the mammalian fauna from those deposits has suggested that the vertebrate fossils (including the birds and hominids) were deposited between 1.9 Myr and 2.0 Myr.12 Palaeomagnetic data from a flowstone bounding the lower poorly fossiliferous grey breccia and the higher fossiliferous red breccia (Members 1–5) have been used to suggest the flowstone's deposition during the Matuyama palaeomagnetic chron, and thus the grey breccia is older than 1.95 Myr and the fossiliferous red breccia is likely very early Pleistocene.8,13
The Kromdraai B lovebird material reported by Pocock3 was collected by C.K. Brain from the decalcified deposits exposed on the north face of the cave deposit.14 Pocock3 did not report from which stratigraphic level those specimens were derived, and therefore the parrot specimens could have come from almost anywhere in the deposit's stratigraphy.10 The specimen described below was uncovered during Vrba's excavations in 1977 to 1980, and it is from her Member 3 where most fossils, and all hominid fossils, have been collected.5,10 Therefore, this lovebird would have lived penecontemporaneously with Australopithecus robustus.
Institutional abbreviations: KB, Kromdraai B collection housed at the Transvaal Museum, Pretoria, South Africa; MVZ, Museum of Vertebrate Zoology, University of California, Berkeley, U.S.A.; NMB, National Museum, Bloemfontein, South Africa; TM, Ornithology Collection, Transvaal Museum; ZO, Iziko South African Museum, Cape Town.
Order Psittaciformes Wagler 1830
Family Psittacidae Illiger 1811
Genus Agapornis Selby 1836
Description
The specimen is KB 5525, a complete right humerus (Fig. 1). The bone is black in colour, covered in part with rust-coloured sediment, and has slight damage to the distal dorsal corner of the bone.
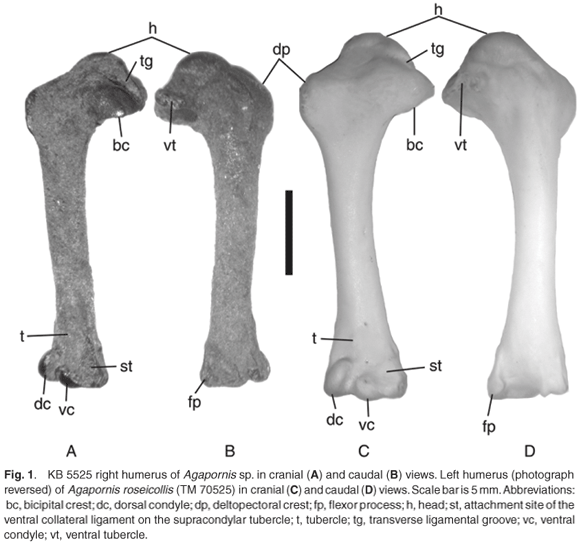
Caudal view: The head is rounded and projected proximally. Extending dorsally from the head dorsodistally is an elongate ridge representing an elongated dorsal tubercle. However, there is no distinct dorsal tubercle on that ridge. The area distal to the head and ridge is concave. The ventral tubercle is a slightly expanded spot on a dorso-ventrally oriented ridge. The bicipital crest extends distal to the ventral tubercle, but not as far distal as the distal end of the deltopectoral crest. The pneumotricipital fossa is relatively shallow and reaches proximally to the level of the distal edge of the head. The capital incisure (between the head and the ventral tubercle) is an elongate furrow. The dorsoventral midpoint of the head lies ventral to the humeral shaft. The shaft is straight and nearly uniform in diameter. The distal end widens slightly. The flexor process extends distally past the distal condyles. The flexor process has a concave dorsal face and a distinct pit in it for the origin of the flexor carpi ulnaris muscle. The olecranon fossa is relatively wide, and it has a groove on its dorsal side. The ventral condyle extends distal to the level of the dorsal condyle.
Cranial view: The transverse ligamental groove extends from the ventral edge (at the bicipital crest) dorsally to the midpoint of the head, and it is narrower at its ventral end. The area between the head, deltopectoral crest, and transverse ligamental groove is slightly concave. The straight deltopectoral crest is distal to the level of the transverse ligamental groove. The distal end of the humerus has a relatively straight margin. A small tubercle is present proximal to the midpoint between the distal condyles. The brachial fossa is very indistinct overall, deepens ventrally, and is deepest ventral to the level of the ventral condyle. The dorsal condyle extends proximal to the level of the midpoint between the ventral condyle and tubercle. The attachment site of the ventral collateral ligament (located on the ventral supracondylar tubercle) is subcircular in outline and does not extend as far proximally as the dorsal condyle does. Proximal to the ventral supracondylar tubercle is the ridge that bounds the ventral margin of the brachial fossa.
Ventral view: There are two pits on the distal end for muscle origination, and they are subequal in size. The area proximal to the pits is relatively flat, and there is a pit at the proximal end (for the origin of one of the pronator muscles) that is adjacent to the proximal end of the ridge.
Dorsal view: The area containing the distal muscle origination pits is indistinct. There is a slightly raised ridge of the dorsal surface that extends proximal to the level of the dorsal tubercle (on the anterior surface) and ventral ridge.
Comparisons
The fossil humerus is smaller overall than the same element in the extant Agapornis specimens examined (Table 1), but it is close in size to Agapornis lilianae. I was not able to examine one of the extant species, A. swinderianus from Central Africa. It is likely that the humerus of A. swinderianus would not be significantly smaller than any of the other taxa examined because its published body size measurements15 are within the range of the remaining eight species of lovebirds. Overall, the fossil and extant specimens have a very similar morphology. All specimens (fossil and recent) have a very small tubercle on the distal end of the humerus proximal to the midpoint between the distal condyles. The attachment site of the ventral collateral ligament is more triangular in outline in all extant specimens examined than the more rounded shape present in the fossil. The ridge proximal to the ventral supracondylar tubercle that bounds the brachial fossa is much less distinct in A. roseicollis than in the fossil.

The presence of a very small tubercle on the anterior face of the distal end, proximal to the midpoint between the distal condyles, occurs only in Agapornis, among the genera of parrots that occur in Africa (Psittacus, Psittacula and Poicephalus). However, this morphological detail also occurs in some Asian/ Australasian parrot genera (Micropsitta meeki proxima MVZ 90139 and Loriculus galgulus MVZ 169143) that have been hypothesised to be close relatives of Agapornis.16 The position of that tubercle is more dorsally located (proximal to the dorsal condyle) in the other African parrot genera Poicephalus, Psittacus and Psittacula (as well as in many other genera examined from Australasia and South America). That dorsal position is likely the plesiomorphic condition among parrots. The apparently derived, more ventrally located tubercle position, along with the morphology of the hypotarsus,16 appears to reinforce the idea that Agapornis, Loriculus, and Micropsitta are close relatives.
Additional details of the fossil's humeral morphology differ from those of extant Agapornis species examined (Table 1) mainly in the shape of the attachment site of the ventral collateral ligament, and the shape of the brachial fossa. In addition, the fossil specimen's smaller size compared with the extant taxa measured (that include the smallest extant species and the species that are geographically nearest to Kromdraai B today), suggests that the Kromdraai B lovebird is an extinct taxon distinct from extant members of Agapornis. The subcircular shape of the attachment site of the ventral collateral ligament may be an autapomorphy of that extinct species, but additional fossil material is required to determine if that character is variable (or not) among other individuals. The Kromdraai B lovebird's smaller size is consistent with the presence of a small-sized lovebird at Plovers Lake,4 but the Kromdraai B specimen does not appear to be as small as the Plovers Lake taxon. The Kromdraai B specimen is ~91% the size (in humeral length) of A. roseicollis, and the Plovers Lake specimen is ~70% of the size (in quadrate height) of A. roseicollis. Given the discrepancy in size between the fossil specimens and one extant species, there may have been two taxa of lovebirds (that differed in size) in the Pleistocene of the Bloubank River Valley, as originally suggested by Pocock.3
Palaeoenvironment
A variety of palaeoenvironmental interpretations of the area around Kromdraai B have been proposed over the years, but overall it has been suggested to differ from the typical grassland savannah palaeohabitat hypothesised at other sites in the Bloubank River Valley. Brain6 suggested a wetter environment (with over 1 m of rain per year) than at present, based on his interpretation of the Kromdraai B sediments. Partridge17 also suggested a more humid habitat than today. Based on the mammalian fauna, the palaeoenvironment was interpreted as having very little bush cover;18 taxonomic revision changed this interpretation to a habitat with greater abundance of water and forest than at present.10 Scott and Bonnefille19 presented pollen samples from Kromdraai B that are dominated by grass and aloes, possibly suggesting the presence of a protea savanna.17 The most recent analysis of the Kromdraai B mammalian fauna suggests an even greater past presence of closed and forested habitats, since the fauna is dominated by open and closed woodland and forest taxa.20 The bird fauna from the site previously has not been used to posit a reconstruction of the palaeoenvironment, but instead the avian fossils were interpreted only as components of an assumed grassveld environment.3 Lovebirds today are typically residents of a wide variety of forested and wooded environments and are not migratory.15 Despite that overall habitat preference, the diet of some lovebird species includes grass seeds, and most species can occur in mixed grassy and wooded environments.15 Therefore, the presence of resident lovebirds at this site is consistent with and helps to reinforce the suggestion that the dominant habitat type around Kromdraai B at the time when Australopithecus robustus occupied the valley was wooded or forested (though with some grass component). In addition, the presence of lovebirds in the Bloubank Valley (indicated by specimens at Plovers Lake and Kromdraai B) points to the potential occupation of the valley by lovebirds for approximately one million years (~1 Myr to 1.9 Myr). The absence of lovebirds in the region today, may be related to the significant climatic and environmental changes (driven by the glacial and interglacial cycles of the Pleistocene) that also presumably affected the hominids and coeval mammalian taxa of the valley.21
I wish to thank Stephany Potze at the Transvaal Museum for her invaluable assistance. Graham Avery, Dawie de Swardt, Tamara Cassidy and Carla Cicero provided access and aid with modern comparative skeletal material. I thank Darryl de Ruiter, C.K. Brain, and Francis Thackeray for discussions about Kromdraai B and the Bloubank Valley deposits. Graham Avery and an anonymous reviewer provided valuable comments on an earlier draft of the manuscript.
1. Brodkorb P. (1985). Preliminary report on Pliocene/Pleistocene birds of East Africa. In Acta of the 18th International Ornithological Congress, eds V.D. Ilyichev and V.M. Gavrilov, pp. 174–177. Nauka, Moscow. [ Links ]
2. Stidham T.A. (2006). Parrots (Aves: Psittaciformes) from the Miocene Varswater Formation, Langebaanweg, South Africa. Afr. Nat. Hist. 2, 198–199. [ Links ]
3. Pocock T.N. (1971). Pleistocene bird fossils from Kromdraai and Sterkfontein. The Ostrich suppl. 8, 1–6. [ Links ]
4. Stidham T.A. (in press). A small Pleistocene lovebird (Psittacidae: Agapornis) from Plovers Lake, South Africa. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. [ Links ]
5. Thackeray J.F., deRuiter D.J., Berger L.R. and van der Merwe N.J. (2001). Hominid fossils from Kromdraai: a revised list of specimens discovered since 1938. Ann. Transvaal Mus. 38, 43–91. [ Links ]
6. Brain C.K. (1958). The Transvaal ape-man-bearing cave deposits. Transvaal Mus. Mem. no. 11. [ Links ]
7. Partridge T.C. (1982). Some preliminary observations on the stratigraphy and sedimentology of the Kromdraai B hominid site. Palaeoecology of Africa and the Surrounding Islands 15, 3–12. [ Links ]
8. Thackeray J.F., Kirschvink J.L. and Raub T.D. (2002). Palaeomagnetic analyses of calcified deposits from the Plio-Pleistocene hominid site of Kromdraai, South Africa. S. Afr. J. Sci. 98, 537–540. [ Links ]
9. Thackeray J.F., Sénégas F. and Wallace I. (2005). The distribution of cave breccias at Kromdraai A and B in relation to dolomite. Ann. Transvaal Mus. 42, 89–91. [ Links ]
10. Vrba E.S. (1981). The Kromdraai australopithecine site revisited in 1980; recent investigations and results. Ann.Transvaal Mus. 33, 17–60. [ Links ]
11. Vrba E.S. and Panagos D.C. (1982). New perspectives on taphonomy, palaeoecology, and chronology of the Kromdraai apeman. Palaeoecology of Africa and the Surrounding Islands 15, 13–26. [ Links ]
12. McKee J.K. (1995). Further chronological seriations of Southern African Pliocene and Pleistocene mammalian faunal assemblages. Palaeontologia Africana 32, 11–16. [ Links ]
13. Gradstein F., Ogg J. and Smith A. (2004). In A Geologic Time Scale 2004. Cambridge University Press, Cambridge. [ Links ]
14. Brain C.K. (1975). An interpretation of the bone assemblage from the Kromdraai australopithecine site, South Africa. In Paleoanthropology, Morphology and Paleoecology, ed. R.H. Tuttle, pp. 225–243. Mouton Publications, The Hague. [ Links ]
15. Juniper T. and Parr M. (1998). In Parrots: A Guide to Parrots of the World. Yale University Press, New Haven. [ Links ]
16. Mayr G. (2008). The phylogenetic affinities of the parrot taxa Agapornis, Loriculus and Melopsittacus (Aves: Psittaciformes): hypotarsal morphology supports the results of molecular analyses. Emu 108, 23–27. [ Links ]
17. Partridge T.C. (1986). Palaeoecology of the Pliocene and lower Pleistocene hominids of Southern Africa: how good is the chronological and palaeoenvironmental evidence? S. Afr. J. Sci. 82, 80–83. [ Links ]
18. Vrba E.S. (1975). Some evidence of chronology and palaeoecology of Sterkfontein, Swartkrans, and Kromdraai from the fossil Bovidae. Nature 254, 301–304. [ Links ]
19. Scott L. and Bonnefille R. (1986). Search for pollen from the hominid deposits of Kromdraai, Sterkfontein, and Swartkrans: some problems and preliminary results. S. Afr. J. Sci. 82, 380–382. [ Links ]
20. de Ruiter D.J., Sponheimer M. and Lee-Thorpe J.A. (2008). Indications of habitat association of Australopithecus robustus in the Bloubank Valley, South Africa. J. Human Evol. 55, 1015–1030. [ Links ]
21. deMenocal P.B. (2004). African climate change and faunal evolution during the Pliocene-Pleistocene. Earth Planet. Sci. Lett. 220, 3–24. [ Links ]
Received 19 February. Accepted 4 May 2009.

