










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.105 no.3-4 Pretoria mar./abr. 2009
RESEARCH ARTICLES
Metal–biomass interactions: a comparison of visualisation techniques available in South Africa
B.A. MooreI, V, *; C. MackII, V; J.R. DuncanIII; J.E. BurgessIV, V
IInstitute of Water Research, Rhodes University, P.O. Box 94,Grahamstown 6139, South Africa
IIGolder and Associates, P.O. Box 6001, Halfway House 1685, South Africa
IIIDean of Research, Rhodes University, P.O.Box 94, Grahamstown 6139, South Africa
IVWater Research Commission, Private Bag X03, Gezina 0031, South Africa
VFormerly: Department of Biochemistry, Microbiology and Biotechnology, Rhodes University
ABSTRACT
The interaction of metals and biological materials is of interest for reasons such as metal recovery, toxicity and production of high-value products such as gold and platinum nanoparticles. Understanding the way in which metals interact with the biomass surface and intracellular components provides insights into the biosorption and bioaccumulation processes and increases the potential for process optimisation. Three technologies are available for the qualitative visualisation of metal–biomass interactions in South Africa, namely, micro-PIXE, transmission electron microscopy and scanning electron microscopy coupled with energy-dispersive analysis of X-rays. Each technique provides unique information and has specific shortcomings which should be taken into account when selecting the appropriate technology. This paper focuses on evaluating the various techniques.
Key words: bioaccumulation, biosorption, visualisation, TEM, SEM-EDAX, micro-PIXE
Introduction
Concern regarding the release of metals into the environment is a recurring theme in the fight against pollution of natural habitats.1 Most conventional metal analysis techniques are useful for the determination of bulk metal concentrations. These include atomic absorption spectroscopy (AAS), atomic flame spectroscopy (AFS) and inductively coupled plasma-atomic emission spectrometry (ICP-AES) to name a few.2
However, more specific information with regard to metal localisation and concentrations is required for the analysis of interactions of metals with living cells3 to fully understand and exploit the bioremediation potential of a wide variety of organisms.
Several technologies are available to qualitatively determine the interactions of metals with living and non-living biomass. These include micro-particle-induced X-ray emission (micro-PIXE), scanning electron microscopy with energy-dispersive analysis of X-rays (SEM-EDAX) and transmission electron microscopy (TEM), all of which have established themselves as sensitive, qualitative, visual metal analysis techniques.4–6
This work aims to compare the various techniques available in South Africa, in terms of the qualitative information generated, with regard to the interaction of platinum with fungal and yeast biomass.
Materials and methods
Unless otherwise stated, all growth media and reagents were reagent-grade Biolab and Saarchem brands, respectively, supplied by Merck Chemicals (Pty) Ltd, South Africa and used as received. All glassware was acid-washed in 5% HNO3 for a minimum of five hours, rinsed in deionised water overnight and air-dried before use.
Micro-particle-induced X-ray emission (micro-PIXE) analysis
Malt extract broth (20 g l–1 malt extract, 10 g l–1 glucose and 2 g l–1 yeast extract) was inoculated with the fungus Phoma glomerata and allowed to grow at 28°C with shaking for four days, after which platinum solution (1 000 mg l–1 platinum in 0.5 M hydrochloric acid (HCl), EC Labs SA) was added to generate a final concentration of 100 mg l–1 of each metal ion. After an additional two days, biomass samples were removed for micro-PIXE analysis.
The biomass specimen was prepared by homogenising one mycelial ball in distilled water, mounting on a double 1% formvar (Sigma-Aldrich, South Africa) frame and cryofixing (Leica EM CPC, Austria) in instrument-grade liquid propane (Afrox, South Africa). The specimen was then freeze-dried at –90°C (Leica EM CFD, Austria) for 24 hours.
Microanalysis of the specimen was performed using a nuclear microprobe (Materials Research Group, iThemba Labs, South Africa).7 A proton beam with a current of 300 pA and 3.0 MeV was focused on a 5 × 5 µm area of the specimen and raster scanned. Backscattering and PIXE were used concurrently with an external 155 µm Kapton absorber placed between the PIXE Si(Li) detector and the specimen. Data were collected using a XSYS data acquisition system and further processed with GeoPIXE-II software.8 Quantitative elemental maps were produced using the 'dynamic analysis' method.9 Average elemental concentrations for the specimen were calculated on the basis of the PIXE spectra obtained from the selected area and analysis of the corresponding backscattering spectra using a RUMP simulation package. Non-Rutherford cross-sections for isotopically-natural carbon and oxygen, at a laboratory angle of 170°C, provided the data on matrix composition and areal density.
Transmission electron microscopy (TEM)
Transmission electron microscopy was carried out as described by Cross et al.10 Phoma glomerata samples used for TEM were prepared in platinum solution as described above and fixed in cold buffered fixative (2.5% (v/v) gluteraldehyde in 0.1 M sodium phosphate buffer, pH 7) at 4°C for 12 hours, after which they were exposed to osmium tetroxide for 60 minutes before undergoing two 10-minute wash steps with 2.5% gluteraldehyde in 0.1 M sodium phosphate buffer (pH 7). Samples were then dehydrated with a series of increasing ethanol concentrations (30, 50, 70, 80, 90, 100%) with two changes of 100% ethanol. Once complete, the ethanol was discarded and replaced with propylene oxide (Sigma-Aldrich, South Africa) and left for 15 minutes, after which it was replaced with fresh propylene oxide for a further 15 minutes. The propylene oxide was discarded and the specimens exposed to a series of propylene oxide:resin (v/v) mixtures with increasing concentrations of resin (75:25, 50:50, 25:75) each of which was allowed to infiltrate for 90 minutes. The specimens were each transferred to pure resin and allowed to infiltrate overnight.
Specimens were placed at 60°C for 36 hours to allow the resin to polymerise after which they were sectioned and viewed with a transmission electron microscope at 90 kV (JEOL 1210, Japan).
Scanning electron microscopy with energy-dispersive analysis of X-rays (SEM-EDAX)
Yeast biomass (Saccharomyces cerevisiae) was immobilised using polyethyleneimine (PEI) and gluteraldehyde (GA) as embedding and cross-linking agents, respectively; they were subsequently treated with hot alkali solution11 (3% potassium hydroxide (KOH), Merck Chemicals, South Africa). Forty grams (wet weight) of yeast was mixed with 4 ml distilled water to form a smooth paste. A 25% (v/v) solution of GA (50% aqueous gluteraldehyde, Sigma-Aldrich, South Africa) was added to the yeast paste and blended until smooth. A 33% (v/v) PEI (50% aqueous polyethyleneimine, Sigma-Aldrich, South Africa) solution was added and the mixture was blended until a moist dough was obtained. This dough was crumbled to form granules which were then oven-dried overnight at 80°C.
The immobilised yeast pellets were suspended in 3% (w/v) KOH solution and heated to 70°C in a water bath. The granules were then settled and separated from the alkali solution. The alkali solution was retained and the biomass washed twice with distilled water (the wash water being added to the alkali solution each time). The alkali-soluble components of the biomass were reconstituted by acidification of the alkali solution with 32% HCl. The HCl was added with stirring until a pH of 6.0 was obtained. Subsequent centrifugation (400 g for 10 minutes) pelleted out the reconstituted biomass. The washed and reconstituted biomass was then added to the alkali-insoluble fraction and both were oven-dried overnight at 80°C. Two grams of the PEIGA immobilised S. cerevisiae were incubated with 20 mg l–1 of platinum for three hours, after which the biomass was separated and dried at 50°C overnight and then stored in airtight Eppendorf tubes.
Scanning electron microscope images were taken using a Quanta 600 SEM (FEI, USA) equipped with X-ray microanalysis capabilities (Genesis, EDAX, USA). Samples were carbon-coated to minimise the effect of electron charging of the surface, which would have distorted the images.
Two modes of viewing were used; backscattered electron mode, which highlights areas of non-biological composition (i.e. metals) and simultaneous secondary electron mode which allows for the observation of biological microstructures.
Results and discussion
Micro-PIXE
Micro-PIXE analysis of P. glomerata was undertaken with the Materials Research Group, iThemba Laboratory for Accelerator Based Sciences, at Faure in the Western Cape. After preparation of the specimen, it was discovered that the single hyphae were too small to determine localisation patterns of the platinum ions within the cell so an area with a high hyphal density was selected for evaluation of the metal concentrations.
The areal density of the specimen was determined to be 0.65 mg cm–2 and further determinations of elemental concentrations were calculated based on these values.
In Fig. 1 a quantitative map of the P. glomerata and platinum specimen can be seen. The platinum concentration was determined to be 5 617 mg kg–1.
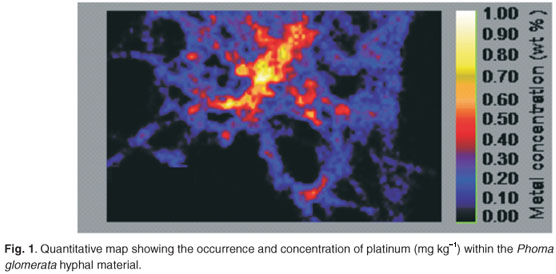
The majority of previous work published on fungal–metal interaction analysis with micro-PIXE has been undertaken with arbuscular mycorrhizal fungi and their respective plant associations.4,12–14 Phoma glomerata is a non-mycorrhizal fungus with smaller hyphae than the mycorrhizal fungi used in previous work. Thus the fundamental difference between the assessment of elemental distribution of metals within mycorrhizal fungi and the results reported here is the respective size of the fungal structures. Weiersbye et al.4 examined a mycorrhizal fungus specimen with a 1 000 P. glomerata× 1 000 µm2 area, while Turnau et al.14 used a mycorrhizal fungus with a scan size of 10 × 10 µm2. The individual P. glomerata hyphae were determined to be approximately 1 µm in diameter, smaller than the individual beam spot of the instrument (3 × 3 µm). While the facility in South Africa may not be sensitive enough for the determination of metals in specimens as small as 1 µm, upgraded instruments (University of Liepzig, Germany) with a beam size of 0.5 µm2 make it possible to achieve a resolution of 260 × 280 nm2 at 10 pA 2.25 MeV.15
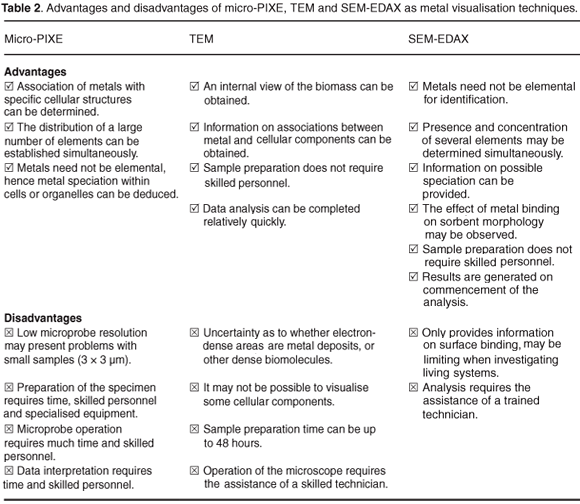
The key advantage of using micro-PIXE to determine metal distribution within organisms or cells is that, if large enough, association of metals with specific cellular structures can be determined. In addition, the distribution of a large number of elements can be established simultaneously and since metals need not be elemental, metal speciation within the cells or organelles can be deduced.
Limitations of this technique for metal–biomass interactions include the following: the specimen needs to be large enough for analysis to be effective; preparation of the specimen may take a number of days and requires skilled personnel and equipment (freeze driers and cryogenic equipment); and operation of the nuclear microprobe and post-analysis data interpretation requires highly-skilled individuals and analysis time is comparatively long (12 hours or more). In addition, while subsidy is provided by the National Research Foundation of South Africa, under specific intellectual property and collaborative conditions, if users decide to retain full intellectual property of their analysis, use of the microprobe may be prohibitively expensive.
Transmission electron microscopy (TEM)
Transmission electron microscopy was carried out on P. glomerata, growing in a platinum solution, to ascertain if the metal was being transported into the cell or binding to the cell surface alone. The presence of metals was detected by TEM as electron-dense areas (dark areas) on the biomass specimens.16 Figure 2 shows P. glomerata biomass with metal deposition on the outside of the cell wall. By contrast, the control had no electron-dense areas around the cell wall (data not shown). In addition to the metal deposits around the outside of the cell, discrete particles were observed within the cytoplasm of the fungal cells. The metal appeared to be evenly distributed throughout the cytoplasm, rather than being associated with a particular cellular structure.
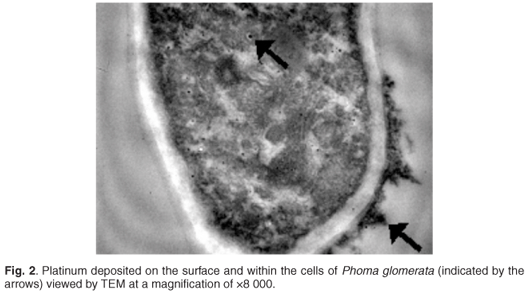
The particles seen within the cytoplasm have been documented by Gericke and Pinches5 as being metal nanoparticles produced by microbes in the presence of metal ions, the size and shape of which are dependent on the organism and culture conditions.
There did not appear to be any specific localisation of the platinum within the cell. However, there may have been binding to cellular structures, but since staining with osmium tetroxide was necessary to visualise the biomass, metal ions associated with specific organelles may have been masked as the tissue became darker.10
As transmission electron microscopy provides an indication of the presence of elemental metal it is possible that some of the metal ions taken up by the P. glomerata biomass could not be visualised as they may not have been in the elemental form, but rather formed complexes with various biomolecules. Nonetheless, metal deposition was clearly visible on the cell surfaces within the cytoplasm. This is supported by Gericke and Pinches5 who screened a range of fungi, yeast and bacteria for their ability to metabolically produce gold nanoparticles. The authors found that the fungus Verticillium lutteoalbum and the yeast Pichia jadinii produced the highest concentration of nanoparticles; however, none of the organisms presented by Gericke and Pinches5 deposited metal ions on the outside of the cell wall. Conversely, Macaskie et al.17 investigated the production of palladium crystals by the bacterium Desulfovibrio desulfuricans and reported that the nanoparticle crystals were only formed on the outside of the cell.
The most useful aspect of this technique for visualisation of metal–biomass interaction is that an internal view of the biomass can be obtained, which is especially valuable in bioaccumulation studies. Furthermore, if staining with osmium tetroxide can be avoided, information on associations between metal and cellular components can be obtained. Sample preparation does not require skilled personnel and analysis can be completed relatively quickly (a few hours). Drawbacks of this technique for metal visualisation include: uncertainty as to whether electron-dense areas are metal deposits, or some other dense biomolecules; if the use of osmium tetroxide is avoided, it may not be possible to visualise some cellular components; preparation time is lengthy (up to 48 hours) and operation of the microscope requires the assistance of a skilled technician.
SEM-EDAX
A number of authors have used the SEM-EDAX technique to demonstrate the sorption of metal ions onto a variety of biomass surfaces.6,18,19 In this study PEIGA-immobilised S. cerevisiae was incubated with platinum ions and viewed by SEM-EDAX. Figure 3A illustrates the biological component of the immobilised yeast cells; the light area in Fig. 3B was attributed to the presence of non-biological compounds on the surface of the yeast cells.
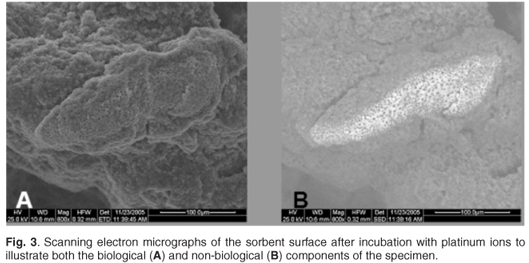
Both the light and dark areas of the specimen in Fig. 3 were scanned using X-ray microanalysis to determine the composition of the non-biological residue. An energy-dispersive spectroscopic representation (Fig. 4) of the surface composition of the dark and light areas revealed that platinum and chloride concentrations in the dark area (Fig. 4A) were lower than in the light area (Fig. 4B). This confirmed the sorption of platinum onto the immobilised yeast surface.
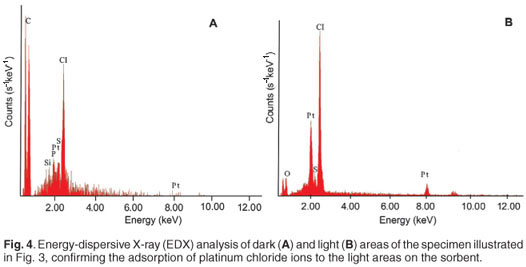
Advantages of using SEM-EDAX are that, as with micro-PIXE, metals need not be in an elemental form for identification and the presence and concentration of a number of elements may be determined simultaneously, providing information on possible speciation. Since the surface of the sorbent is visualised, the effect of metal binding on morphology may be observed using this technique. In addition, sample preparation does not require skilled personnel and results are generated upon commencement of the analysis.
The significant drawback of this technique is that it only provides information on surface binding, which may be limiting when investigating bioaccumulation in living systems. Of less importance is that analysis requires the assistance of a trained technician.
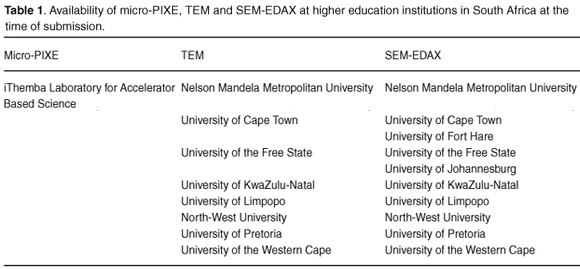
Availability of micro-PIXE, TEM and SEM-EDAX
While analysis of metal–biomass interactions with micro-PIXE may generate the largest amount of data, the restrictions in terms of cost and requirements of additional facilities and trained personnel are compounded by the fact that only one nuclear microprobe is available in South Africa (Table 1). The limitations of the instrument, in terms of minimum specimen size, must also be taken into account when considering the use of micro-PIXE for this type of analysis. A number of institutions in South Africa have confirmed the availability of TEM and SEM-EDAX facilities, outlined in Table 1. The use of TEM or SEM-EDAX will depend on the particular information required. Biosorption studies may be undertaken with both TEM and SEM-EDAX to visualise the biomass surface; however, bioaccumulation of metals can only be established through the use of TEM.
Conclusion
The key advantages and disadvantages of the methods described here are summarised in Table 2. While visualisation of metal–biomass interactions may be achieved through any of the methods discussed, perhaps a combination of analyses will provide a more conclusive representation of the true nature of the interactions. Fourier transform infrared spectroscopy (FTIR) and X-ray diffraction (XRD) analysis may supplement visual data, providing a more holistic overview of the bioaccumulation/biosorption process.
1. Volesky B. (2001). Detoxification of metal-bearing effluents: biosorption for the next century. Hydrometallurgy 59, 203–216. [ Links ]
2. Garman E.F. and Grime G.W. (2005). Elemental analysis of protein by microPIXE. Prog. Biophys. Mol. Biol. 89, 173–205. [ Links ]
3. Przybyłowicz W.J., Mesjasz-Przybyłowicz J., Migula P., Turnau K., Nakonieczny M., Augustyniak M. and Głowacka E. (2004). Elemental microanalysis in ecophysiology using ion microbeam. Nucl. Instrum. Meth. B 219–220, 57–66. [ Links ]
4. Weiersbye I.M., Straker C.J. and Przybyłowicz W.J. (1999). Micro-PIXE mapping of elemental distribution in arbuscular mycorrhizal roots and the grass, Cynodon dactylon, from gold and uranium mine tailings. Nucl. Instrum. Meth. B 158, 335–343. [ Links ]
5. Gericke M. and Pinches A. (2006). Biological synthesis of metal nanoparticles. Hydrometallurgy 83, 123–140. [ Links ]
6. Copello G.J., Varela F., Martínez Vivot R. and Díaz L.E. (2008). Immobilized chitosan as a biosorbent for the removal of Cd(II), Cr(III) and Cr(VI) from aqueous solutions. Bioresour. Technol. 99, 6538 – 6544. [ Links ]
7. Prozesky V.M., Przybyłowicz W.J., van Achterbergh E., Churms C.L., Pineda C.A., Springhorn K.A., Pilcher J.V., Ryan C.G., Kritzinger J., Schmitt H. and Swart T. (1995). The NAC nuclear microprobe facility. Nucl. Instrum. Meth. B 104, 36–42. [ Links ]
8. Ryan C.G., van Achterbergh E., Yeats C.J., Drieberg S.L., Mark G., McInnes B.M., Win T.T., Cripps W. and Suter G.F. (2002). Quantitative, high sensitivity, high resolution, nuclear microprobe imaging of fluids, melts and minerals. Nucl. Instrum. Meth. B 188, 18–27. [ Links ]
9. Ryan C.G., Jamieson D.N., Churms C.L. and Pilcher J.V. (1995). A new method for on-line true-elemental imaging using PIXE and the microprobe. Nucl. Instrum. Meth. B 104, 157–165. [ Links ]
10. Cross R., Botha T. and Pinchuck S. (2001). The preparation of biological material for electron microscopy. Electron Microscopy Unit, Rhodes University, Grahamstown. [ Links ]
11. Stoll A. and Duncan J.R. (1997). Comparison of the heavy metal sorptive properties of three types of immobilized, non-viable Saccharomyces cerevisiae biomass. Process Biochem. 32, 467–472. [ Links ]
12. Jurkiewicz A., Turnau K., Mesjasz-Przybyłowicz J., Przybyłowicz W. and Godzik B. (2001). Heavy metal localisation in mycorrhizas of Epipactis atrorubens (Hoffm.) Besser (Orchidaceae) from zinc mine tailings. Protoplasma 218, 117–124. [ Links ]
13. Scheloske S., Maetz M. and Schüßler A. (2001). Heavy metal uptake of Geosiphon pyriforme. Nucl. Instrum. Meth. B 181, 659–663. [ Links ]
14. Turnau K., Przybyłowicz W.J. and Mesjasz-Przybyłowicz J. (2001). Heavy metal distribution in Suillus luteus mycorrhizas – as revealed by micro-PIXE analysis. Nucl. Instrum. Meth. B 181, 649–658. [ Links ]
15. Reinert T., Spemann D., Morawski M. and Arendt T. (2008). Quantitative trace element analysis with sub-micron lateral resolution. Nucl. Instrum. Meth. B 249, 734–737. [ Links ]
16. Yamato H., Koshiba T., Ohwa M., Wernet W. and Matsumura M. (1997). A new method for dispersing palladium microparticles in conducting polymer films and its application in biosensors. Synth. Met. 87, 231–236. [ Links ]
17. Macaskie L.E., Baxter-Plant V.S., Creamer N.J., Humphries A.C., Mikheenko I.P., Mikheenko P.M., Penfold D.W. and Yong P. (2005). Applications of bacterial hydrogenases in waste decontamination, manufacture of novel bionanocatalysts and in sustainable energy. Biochem. Soc. Trans. 33, 76–79. [ Links ]
18. Tomašević M., Vukmirović Z., Rajšić S., Tasi M. and Stevanović B. (2005). Characterisation of trace metal particles deposited on some deciduous tree leaves in an urban area. Chemosphere 61, 753–760. [ Links ]
19. Tunali S., Çabuk A. and Akar T. (2006). Removal of lead and copper ions from aqueous solutions by bacterial strain isolated from soil. Chem. Eng. J. 115, 203–211. [ Links ]
Received 12 January. Accepted 28 April 2009.
* Author for correspondence E-mail: g02m2567@campus.ru.ac.za

