










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.104 n.11-12 Pretoria Nov./Dec. 2008
RESEARCH LETTERS
The first fossil of the Congo peafowl (Galliformes: Afropavo)
Thomas A. Stidham
Department of Biology, 3258 TAMU, Texas A & M University, College Station, TX 77943-3258, U.S.A. E-mail: furcula@mail.bio.tamu.edu
ABSTRACT
A fossil from the Middle Stone Age internal deposits of Plovers Lake Cave in the Sterkfontein Valley, Gauteng province, South Africa, is the first fossil specimen that can be allocated to the Congo peafowl (Afropavo congensis), a species that is currently endemic to the Democratic Republic of Congo in central Africa. The fossil greatly extends the known geographic range of this rainforest taxon and suggests the presence of forested or even rainforest habitats near Plovers Lake Cave during the Middle Stone Age when the fossil was deposited. The presence of this central African taxon in South Africa implies that either its geographic range expanded during a period of climate change during the late Pleistocene or that its currently restricted distribution is a relict of a more extensive past distribution across a larger part of Africa.
Introduction
Excavations at the hominid-bearing locality of Plovers Lake Cave in the Sterkfontein Valley of Gauteng province, South Africa, have produced thousands of fossil vertebrate specimens, including birds. The site includes internal and external deposits. The external deposits are close to 1 Myr in age, and the internal Middle Stone Age deposits are between 62.9 and 88.7 kyr.1–3 Examination of the avifauna from the internal deposits has uncovered a specimen of the Congo peafowl (Afropavo congensis), the first fossil peacock in South Africa and the first record of that species outside of central Africa. The diverse avifauna from the internal Plovers Lake deposits, including guineafowl and passerines, will be described elsewhere.
Until recently, the fossil record of peafowl (Pavonini) in Africa was unknown. However, fossils of the genus Pavo have been recovered from early Pliocene deposits in Ethiopia and Kenya, demonstrating the presence of the peafowl lineage in Africa in the late Neogene.4,5 At present, there is no known fossil record of peafowl in the African Pleistocene and no record of the extant African species prior to historical records. The extant species, Afropavo congensis, is restricted today to the eastern part of the Congo Basin in the Democratic Republic of Congo, and it occurs in undisturbed primary lowland rainforest.6,7 The specimen described below is the first fossil of this unique species and represents a significant expansion of its known geographic range.
Specimen PV 12185 is a proximal right humerus, which is housed at the University of the Witwatersrand in Johannesburg, South Africa.
Description
The specimen is coated over most of its surface with a crust that obscures some of the morphology. There are two punctures in the specimen that appear to be bite marks (Fig. 1). On the posterior surface, one of these punctures is distal to the dorsoventral midpoint of the head and the other is on the anterior surface on the ventral side distal to the level of the distal end of the deltoid crest.
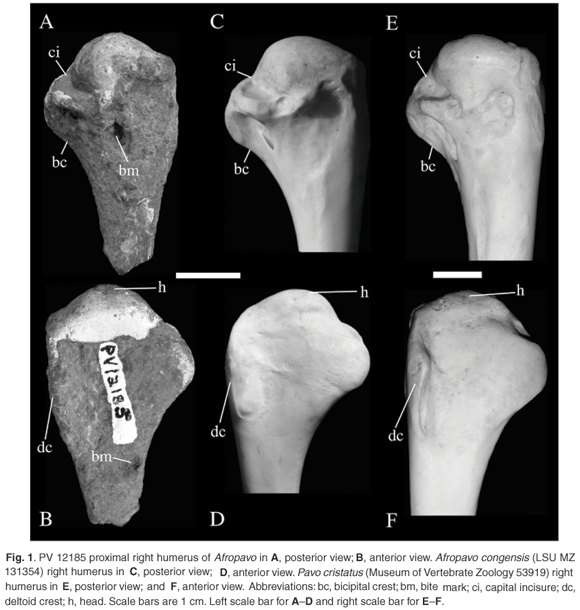
Posterior view. The head is inflated. The dorsal tubercle is an inflated dorsal extension of the head. No ligamental pit is visible on the dorsal tubercle. The dorsal tubercle is proximal to the level of the ventral tubercle. The distal end of the head has a ridge beginning at the dorsoventral midpoint that extends distally and then turns ventrally to the ventral tubercle. The capital incisure is a slit that extends from the proximoventral margin dorsodistally, is deepest anteriorly, and narrows posteriorly. It does not significantly undercut the humeral head. The pneumotricipital fossa is obscured by crust, but appears to be relatively shallow except for the proximal end. The proximal rim of the pneumotricipital fossa is nearly dorsoventrally orientated. The bicipital crest is relatively thick and extends distally about as far as the deltoid crest does.
Anterior view. There is only a slight concavity setting the head off from the rest of the humerus. The deltoid crest is parallel with the long axis of the humerus, and is relatively low. It may be broken, but the area is obscured by the crust. The ligamental groove is very shallow and is ventral to the ventral edge of the head.
Comparisons. The specimen is from a large galliform, placing it in the size range of a guineafowl or a peacock. This fossil specimen has the rounded ventral expansion of the bicipital crest present in Afropavo rather than the more squared-off or blocky outline of the crest in guineafowl. There is also a raised area adjacent to the capital incisure in guineafowl absent in Afropavo and this fossil. Although Pavo and other species of phasianids were present in the late Neogene4,8 of Africa, this fossil has a different morphology from Pavo (see below) and is from a taxon larger than the other phasianids. Other galliforms are present in the fossil deposits at Plovers Lake and are significantly smaller than this fossil taxon. The size of the fossil lies within the range reported for Afropavo and is smaller than that of fossil and recent specimens of Pavo4 (Fig. 1, Table 1).

The fossil's capital incisure is deeper and a more narrow 'V' than in Pavo. The depth and extent of the capital incisure varies among individuals of A. congensis. In Louisiana State University Museum of Zoology (LSUMZ) 131354, the incisure does not undercut the humeral head, as in the fossil specimen (Fig. 1). However, in LSUMZ 172532 the incisure excavates into the ventral side of the head. The fossil specimen's bicipital crest's ventral (distal) margin lacks the more concave shape of Argusianus and Rheinhardia.4 The morphology of the dorsal tubercle is similar to Pavo cristatus, but the fossil most closely resembles that of extant individuals of Afropavo congensis.
Discussion
Afropavo is the sister taxon to Pavo.8 The recent publication of Pliocene specimens of Pavo from Ethiopia and Kenya seems to indicate that the origin of the clade composed of Afropavo and Pavo might have been in Africa.4,5 The split between Afropavo and Pavo has been estimated to be between 9 and 14 Myr,9 older than the oldest fossil specimens of Pavo in Africa.5 The sister relationship between Afropavo and Pavo and the fossil record of Pavo extending into the early Pliocene indicates that Afropavo also should have been present in the early Pliocene. Louchart4 suggested that Afropavo may not have been distributed outside of east-central Africa during its history. The presence of an Afropavo specimen in South Africa younger than the known African specimens of Pavo presents an interesting biogeographical problem. The Plovers Lake specimen indicates that Afropavo was more widely distributed in Africa in the past. The expansion of that taxon outside of central Africa may have been temporary and related to the significant climate change during the late Pleistocene, or its current geographic distribution may in fact be a relict of a larger past distribution that diminished only at the end of the Pleistocene.
Bite marks on the specimen likely indicate that the specimen was brought into the cave by mammalian predators, as many of the coeval mammalian bones appear to have been.2 Thus, the bird probably was not transported very far from where it had lived. The palaeoenvironment of the area around the cave during the Middle Stone Age has been postulated to have been predominantly grassland with some permanent source of water nearby and some small amount of closed/wooded habitat.2,3 However, the presence of the tropical forest taxon Afropavo in the cave deposit is not entirely consistent with that mammal-based palaeoenvironmental hypothesis, and the bird fossil suggests the presence of more closed forest or even rain forest.7 Further analysis of the avian fauna is needed to determine if additional rainforest indicators are present in the cave deposits.
I thank Lee Berger for permission to work on the Plovers Lake avifauna. Darryl de Ruiter and Juliet Brophy provided valuable collections assistance in South Africa. Comments by Marco Pavia and an anonymous reviewer greatly improved the manuscript. I thank the LSU Museum of Natural Science for its generous loan of Congo peacock skeletons. Carl Cicero provided access to skeletons of Pavo species in the Museum of Vertebrate Zoology.
1. Thackeray J.F. and Watson V. (1994). A preliminary account of faunal remains from Plovers Lake. S. Afr. J. Sci. 90, 231–232. [ Links ]
2. Brophy J.K., de Ruiter D.J., Lewis P.J., Churchill S.E. and Burger L.R. (2006). Preliminary investigation of Plover's Lake, South Africa. Curr. Res. Pleist. 23, 41–43. [ Links ]
3. de Ruiter D.J., Brophy J.K., Lewis P.J., Churchill S.E. and Berger L.R. (2008). Faunal assemblage composition and paleoenvironment of Plovers Lake, a Middle Stone Age locality in Gauteng Province, South Africa. J. Hum. Evol. 55, 1102–1117. [ Links ]
4. Louchart A. (2003). A true peafowl in Africa. S. Afr. J. Sci. 99, 368–371. [ Links ]
5. Pickford M., Senut B. and Mourer-Chauviré C. (2004). Early Pliocene Tragulidae and peafowls in the rift valley, Kenya: evidence for rainforest in East Africa. C. R. Palevol 3, 179–189. [ Links ]
6. Dupain J. and Van Krunkelsven E. (1996). Recent observations of the Congo Peacock Afropavo congensis in the Equateur Province, Zaire. Ostrich 67, 94–95. [ Links ]
7. Sinclair I. and Ryan P. (2003). Birds of Africa South of the Sahara. Princeton University Press, Princeton, NJ. [ Links ]
8. Stidham T.A. (2007). Preliminary assessment of the Late Miocene avifauna from Lemudong'o, Kenya. Kirtlandia 56, 173–176. [ Links ]
9. Kimball R.T., Braun E.L. and Ligon J.D. (1997). Resolution of the phylogenetic position of the Congo peafowl, Afropavo congensis: a biogeographic and evolutionary enigma. Proc. R. Soc. Lond. B 264, 1517–1523. [ Links ]
Received 23 January. Accepted 23 November 2008.

