










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Science
versão On-line ISSN 1996-7489
versão impressa ISSN 0038-2353
S. Afr. j. sci. vol.104 no.11-12 Pretoria Nov./Dez. 2008
RESEARCH ARTICLES
Description of the postcranial anatomy of Aulacephalodon baini and its possible relationship with 'Aulacephalodon peavoti'
Romala Govender
Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS 2050, South Africa. E-mail: romala@discoverymail.co.za
ABSTRACT
A recent analysis of the postcranial anatomy of Triassic dicynodonts renewed interest in the anatomy of Aulacephalodon. This study focused initially on Aulacephalodon baini as it represents medium to large Permian dicynodonts but also took into account 'Aulacephalodon peavoti'. The postcranial skeletons of A. baini and 'A. peavoti' show several significant differences. The scapula blade of A. baini is more curved than and not as upright as in 'A. peavoti'. A. baini has a rectangular delto-pectoral crest and has a better developed olecranon. In the pelvic girdle the anterior process is higher than the posterior process of the ilium in A. baini and the acetabulum is directed more ventrally. The femoral head is separated from the greater trochanter in A. baini. A comparison also shows that both A. baini and 'A. peavoti' are different from Rhachiocephalus in that the latter has a widely expanded scapula blade and the proximal end of the humerus is wider than the distal. The glenoid is elliptical with a convex surface. Although it could not be conclusively decided what the taxonomic position of 'A. peavoti' is, this study provides information about the postcranial skeleton which could aid its detailed analysis. This study will also assist in clarifying the relationships as well as providing insight into the palaeobiology of these extinct tetrapods.
Introduction
Dicynodonts are non-mammalian synapsids (therapsids) that rose to become the dominant terrestrial vertebrate herbivores during the Late Permian.1 They maintained their dominance until the Triassic, when they eventually became extinct.2 Permian dicynodonts have been the focus of numerous studies which have included an analysis of the postcranial anatomy.3–5 Some of these studies have also included a functional analysis of the postcranial skeleton.3–5
Aulacephalodon was originally described by Owen in 1844 as Dicynodon baini and was later referred to Aulacephalodon baini. Subsequently, seventeen species were assigned to the genus Aulacephalodon, mostly based on fragmentary material.6 Bainia peavoti was described by Broom in 1921 based on a well-preserved skull.7 In 1932, Broom recognized the validity of the subgenus Aulacephalodon (Seeley 1876) and referred, amongst others, Dicynodon baini and Baini peavoti to 'Aulacocephalodon.'6 Dicynodon (Aulacocephalodon) tigriceps was described by Olson in 1935; upon further inspection, however, it was concluded that the specimen should be referred to Aulacocephalodon peavoti Broom.8 Tollman and Grine undertook an analysis of the cranial morphology of the species of Aulacephalodon and concluded that the series of skulls were a 'morphometrically homogenous, species specific, ontogenetic growth series'.6 The postcranial anatomy of 'A. peavoti' was described in detail in 1938. In 1986, Defauw referred this particular specimen to the genus Rhachiocephalus.
It would seem that the identification of the Field Museum specimen has been called into question at least twice in its history.4,8 For there to be a definitive answer as to the exact identification of FMNH 1532 will require a comparison of skulls, which will prove a problem, as the holotype of 'A. peavoti' is missing (K.D. Angielcyzk, pers. comm.). The postcranial skeleton of FMNH 1532 at least provides information about the species and gives at least some opportunity to determine if it is a species of Aulacephalodon. The study reported here was undertaken in order to describe the postcranial anatomy of Aulacephalodon baini and to determine if it was possible to establish a relationship between A. baini and 'A. peavoti'.
Material
AM5731 represents a juvenile Aulacephalodon. This specimen consists of skull and some postcranial remains which include scapula and humerus.
NMQR 1478 represents an adult Aulacephalodon. This is a fairly large individual and consists of a skull as well as postcranial elements. There is a well-preserved sternum, scapulo-coracoid, ulna, ilium, femur, tibia and fibula. The humerus present in this specimen is a cast. It is also noted that the surface of the bones has suffered cracking and in some instances it is difficult to distinguish between striations and the cracking.
The comparison with 'Aulacephalodon peavoti' is based on photographs provided by K.D. Angielczyk of the specimen (FMNH 1532) at the Field Museum, Chicago, as well as the published description of the specimen from the Field Museum by Olson and Byrne in 1938.
Postcranial anatomy
At this time the taxonomic position of 'Aulacephalodon peavoti' remains poorly understood. The two specimens that are currently assigned to this species were described by Broom (1921) and, later, the specimen described by Olson (1935), which have features that are shared with Aulacephalodon baini (for example, the presence of nasal bosses, wide intertemporal, pineal boss). Furthermore, 'A. peavoti' does not have tusks, which can be considered as a non-Aulacephalodon feature. 'A. peavoti' is included here to determine whether or not it shows any similarity to the postcranial anatomy of A. baini. In order to understand the relationship between A. baini and 'A. peavoti' better, an attempt was made earlier this year to review the holotype—however, it has been lost, which contributes to the taxonomic uncertainty of 'A. peavoti' (K.D. Angielczyk, pers. comm. 2008). The following description is a comparative analysis of the postcranial anatomy of A. baini. The specimen previously described by Olson and Byrne (1938) as 'Aulacephalodon peavoti', who had based their identification on Broom's description, which is considered inadequate (K.D. Angielcyzk, pers. comm.), was referred to Rhachiocephalus by Defauw (1986). There also is a possibility that the specimen might represent Pelanomodon or Odontocyclops, but its identity must remain uncertain until it is re-examined in detail (Angielczyk, pers comm. 2007).
Pectoral girdle and forelimb
Scapula
The scapula of Aulacephalodon baini is a short, fairly robust bone. Dorsally, the border is moderately expanded antero-posteriorly with concave anterior and posterior margins as compared with 'Aulacephalodon peavoti', which has a slightly wider dorsal border and less concave, almost straight anterior and posterior margins. This results in the blade not being angled posteriorly as seen in A. baini (Fig. 1A, B). When compared with other Permian dicynodonts, the scapula of A. baini is proportionally much wider than that of Eodicynodon (pers. obs.),9 Robertia,10 Cistecephalus11 and Diictodon.5 Aulacephalodon baini shows a similar width to that of Kingoria but it is significantly different from that of Kawingasaurus,12 which has a blade-shaped scapula. 'Dicynodon' trigonocephalus has a rectangular blade that is slightly expanded proximally3 but it is not as spatulate as that of Odontocyclops; however, it does have similar curvature along the posterior border.13 A comparison with the similar sized Rhachiocephalus magnus shows that its scapula has a wider flat blade with similarly concave anterior and posterior margins (SAM-PK-K6446).17
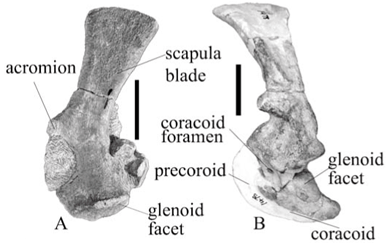
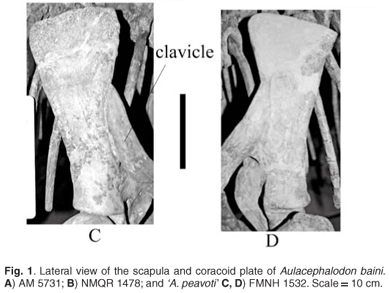
Proximally on the lateral surface of the scapula blade of A. baini there is a shallow fossa that extends to the level of the acromion, which has a smooth surface and narrows at the distal end, which is slightly deeper than a fossa in the same position in 'A. peavoti' (Fig. 1C, D). This fossa is bordered anteriorly and posteriorly by low broad ridges; however, the anterior one is more prominent than the posterior ridge, and has a more rugose surface, which again is more prominent than the borders seen on the scapula of 'A. peavoti'(FMNH 1532).8 Maisch17 described the dorsal third of the scapula of Rhachiocephalus as being almost flat, which suggests that a fossa may well be present but is more shallow than that of A. baini.
There is no evidence of the presence of a scapula spine in A. baini but the surface of the anterior border is antero-posteriorly flattened as seen in other dicynodonts (e.g. Kannemeyeria). From the photographs provided as well as the description of the skeleton by Olson and Byrne,8 there is no scapula spine present on the scapula of 'A. peavoti'; furthermore no scapula spine has been described for Rhachiocephalus.17 A prominent anterior ridge starts about a third of the way below the dorsal border of the scapula in A. baini and has a rugose surface. This is different from other Permian dicynodonts like Kingoria,15 Kawingasaurus12 and Odontocyclops13 as well as some of the Triassic dicynodonts (e.g. Kannemeyeria, Ischigualastia), which all have well-developed scapula spines.
A small, narrow acromion process of A. baini (Fig. 1B) is directed anteriorly with a truncated anterior end, which is similar to that of Robertia,10 as well as that of Rhachiocephalus (PK-K6446).17 It is not well developed as in other dicynodonts but projects slightly beyond the distal end of the scapula. The acromion forms a narrow triangle in Diictodon,4,5 is considered to be continuous with the anterior edge of the scapula blade in Cistecephalus11 and is therefore not considered to be a distinct process.14 In Eodicynodon (pers. obs.)9 and 'D.' trigonocephalus the acromion is rectangular.3 The rugose surface of the ridge along the anterior border of the scapula of A. baini extends diagonally across the lateral surface of the bone passing over the acromion to end below the fossa. Below the acromion the distal end scapula of A. baini is narrower than the proximal end and the lateral surface is concave above the coracoid articulation. Laterally, the surface of the coracoid articulation is slightly concave. The position of the articulating surfaces of the glenoid and the coracoid articulation give the appearance that they are positioned at a small angle to each other. The glenoid facet is round and concavo-convex in the adult A. baini and almost flat in the juvenile, whereas in 'A. peavoti' it has a concave, oval facet and in Rhachiocephalus the facet is elliptical and is antero-medially expanded with a convex surface.17 The glenoid facet of Robertia is almost circular and faces ventrally, posteriorly and slightly laterally.10 In Kawingasaurus the distal end of the scapula and coracoid plate are twisted so that the glenoid faces laterally.12 In Diictodon the glenoid is flat,4 faces caudo-laterally and is sharply defined,5 which is similar to that of A. baini.
The proximal third of the scapula is covered in plaster, but from below it is clearly evident that the medial surface is slightly concave. From about halfway down the medial surface of the blade, a low, broad ridge runs down the middle of the bone to end below the acromion process. The medial surface of the acromion is longitudinally concave and narrow. Posterior to the acromion's medial surface a narrow, shallow groove passes down the distal surface to the distal end of the bone, where it continues onto the proximal surface of the precoracoid. Along the anterior border above the acromion there is a low, oblong tubercle. The surface of the tubercle is very rugose.
Coracoid plate
The coracoid plate is fairly long in A. baini, which is similar to that of 'A. peavoti'. It consists of the precoracoid and coracoid, which are mostly complete in the larger individual (NMQR 1478), although the distal margins of the bones are reconstructed in plaster.
Precoracoid
The precoracoid of A. baini (Fig. 1B) has a wedge-shaped appearance with a fairly long posterior articulation for the coracoid, whereas that of 'A. peavoti' is incompletely preserved and is triangular in Rhachiocephalus.16 Below the dorsal border, almost in the middle of the bone is the oval coracoid foramen in A. baini similar to that seen in Eodicynodon (pers. obs.),8 Diictodon,4,5 Kawingasaurus12 and Cistecephalus,11 whereas in 'A. peavoti' and Rhachiocephalus,13 like Kingoria,15,16 the coracoid foramen is located between the scapula and the precoracoid, which both form the dorsal and ventral parts of the foramen.17 The foramen is surrounded by a relatively narrow, deep groove that continues from the scapula above in A. baini. The distal end of the groove is marked by the distal end of the coracoid foramen. It has a smooth, shiny surface that slopes towards the coracoid foramen.
Coracoid
The coracoid of A. baini (Fig. 1B) forms a right-angled triangle with a rounded distal end and has a straight border where it articulated with the precoracoid. This differs from 'A. peavoti', which has an elongated, almost rectangular coracoid (based on photographs) like that of Robertia,10 Kingoria15,16 and that of Rhachiocephalus, which is shaped like an axe-blade.17 The coracoid of Eodicynodon9 is triangular and square in 'D.' trigonocephalus.3 Along the dorsal border is the facet for the glenoid. The glenoid facet is robust and is separated from the rest of the bone by a broad, distinct groove. Posteriorly, the edge of the glenoid facet is thin and projects slightly over the remainder of the bone. Below this the lateral surface of the coracoid is concave. Posteriorly the narrow distal border projects beyond the glenoid, forming a triangle.
Glenoid
The scapula facet is round with a concavo-convex surface that has a rugose appearance and is convex towards its distal end, where it articulates with the coracoid facet. The coracoid facet is elongated giving it a rectangular appearance with a concavo-convex surface that is convex towards the anterior end of the glenoid. Ridges demarcate the glenoid facet of the coracoid. The glenoid of A. baini is directed caudo-laterally like that seen in Rhachiocephalus16 but the angle between the scapula facet and the coracoid facet of the glenoid of A. baini is slightly greater than 90°, giving the glenoid an open appearance whereas that of Rhachiocephalus has an angle of about 80° between the scapula and coracoid, the glenoid of 'A. peavoti' is directed more laterally but is also more open. The glenoid of Robertia is almost circular and faces ventrally, posteriorly and slightly laterally;10 in Kawingasaurus the distal end of the scapula and coracoid plate are twisted so that the glenoid faces laterally.12 In Diictodon the glenoid is flat,4 faces caudo-laterally and is sharply defined.5
Forelimb
Humerus
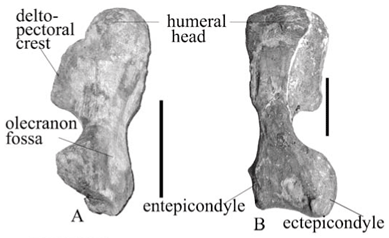
The proximal expansion is almost rectangular in both the larger individual and the smaller one of A. baini (Fig. 2A, B) as well as in 'A. peavoti' (Fig. 2C, D). The proximal and distal ends of the humerus are expanded equally in A. baini and Robertia,10 whereas in Rhachiocephalus, Diictodon and Cistecephalus the proximal expansion is wider than the distal bone.4,11,17 In 'D.' trigonocephalus and Odontocyclops the distal expansion is wider than the proximal one.3,13 In dorsal view the large, almost square head (NMQR 1478) is positioned more anteriorly along the dorsal border of the proximal expansion (Fig. 2A, B) but is not very prominent in the smaller individual (AM 5731), where it appears almost round (Fig. 2A). In 'A. peavoti' the humeral head is round (FMNH 1532).8 The head of the humerus is not prominent in the smaller specimen of A. baini (AM 3731), similar to the situation in Robertia,10 but it is almost square in the larger individual (NMQR 1478). The variation in the shape of the humeral head between the small and large individuals could be explained by ontogeny. In comparison, it is hemispherical in Diictodon,4 Rhachiocephalus17 and sub-hemispherical in Odontocyclops.13 In Cistecephalus the head is clearly demarcated, forming a distinct condyle that is directed dorsally,11 and in Kawingasaurus the humeral head is directed proximo-dorsally,12 forming an elongated condyle. Posterior to the head there is a shallow concave fossa which is also present in 'A. peavoti' but is much shallower (from photographs of FMNH 1532). Its posterio-dorsal border is marked by an irregular, low tubercle.
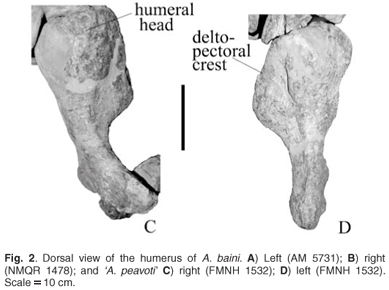
Along the anterior edge of the proximal expansion is a plate-like delto-pectoral crest in A. baini that thins particularly towards the anterior edge, which gives it a rectangular appearance similar to that seen in Rhachiocephalus17 compared with 'A. peavoti', which has an almost semi-circular delto-pectoral crest. There is much variation in the shape of the delto-pectoral crest among the Permian dicynodonts. Robertia has a short, broad and rectangular delto-pectoral crest that is separated from the shaft by a ridge,10 whereas in Diictodon the crest has a straight margin4 and is orientated cranio-ventrally at an angle of 130° to the shaft with a thick cranial edge.5 In Cistecephalus the delto-pectoral crest is thin but broad with a thick leading edge,11 giving it a triangular appearance, which is similar to the morphology of Eodicynodon (pers. obs.). Although the crest in Kawingasaurus is large, it does not extend far distally.12 'D.' trigonocephalus has a robust, massive delto-pectoral crest that is directed anteriorly.3
The delto-pectoral crest is narrow proximally and becomes thick at its ventral end in A. baini (Fig. 2B). Dorsally, the surface of the delto-pectoral crest is flat to slightly concave in A. baini, whereas in 'A. peavoti' (Fig. 2D) the delto-pectoral crest curves ventrally (Fig. 2A). Its posterior extent is marked by a raised oval rugose area and anteriorly the entepicondylar ridge begins below the delto-pectoral crest in A. baini. The shaft of the humerus is broad, short and twisted so that the ectepicondyle is directed dorsally and the surface is concave towards the ectepicondyle in A. baini. Anteriorly on the humerus of A. baini there is a thin entepicondylar ridge that passes over the shaft to the entepicondyle, while the remaining surface is concave and ends on the triangular distal end.
Distally the dorsally directed ectepicondyle is round and the convex articulating surface is directed ventrally and the entepicondyle faces posteriorly on the humerus of A. baini. Below the posterior border of the biccipital fossa is the opening of the oval entepicondylar foramen, with the anterior and posterior ends demarcated by ridges in A. baini.
In ventral view the narrow biccipital fossa has a concave surface in A. baini as compared with that of 'A. peavoti', which has a wider, shallower concave surface. Below the posterior border of the biccipital fossa in A. baini is the narrow, oval opening of the entepicondylar foramen (Fig. 2B). Its borders are marked by narrow ridges anteriorly and posteriorly. Although the humerus of the larger specimen (NMQR 1478) is made up completely of plaster, it does, however, bear a striking resemblance to the figure of the ventral view of Rhachiocephalus(ref. 17, Fig. 3b, p. 166).
The posterior surface of the humerus of A. baini is concave proximally along the biccipital fossa. At the distal end of the biccipital fossa there is a low, irregular tubercle that extends onto the dorsal surface. In posterior view the triangular distal end is made up mostly of the olecranon fossa. It has an almost circular appearance and is only slightly concave with a sharp ridge demarcating the distal border of the fossa.
Ulna
The left ulna is medio-laterally flattened and is a fairly gracile bone in A. baini (Fig. 3A, B); it is more robust than that of 'A. peavoti' (FMNH 1532)8 (Fig. 3C, D). It is wide proximally and narrows to form the shaft and distal end. Proximally the olecranon has a wide, low appearance and there is no evidence of a suture contact between the olecranon and the rest of the bone as compared with that of 'A. peavoti' for which the proximal end is much smaller and the olecranon is smaller and more round (FMNH 1532).8 The surface of the olecranon is rugose. The olecranon of Rhachiocephalus (GPIT K30) is considered also to be poorly developed17 as seen in Robertia, Diictodon and D. trigonocephalus.3,5,10 Cistecephalus has a wide and powerful olecranon,11 and Kawingasaurus has a large olecranon with no evidence of suture contact between the olecranon and the rest of the ulna.12
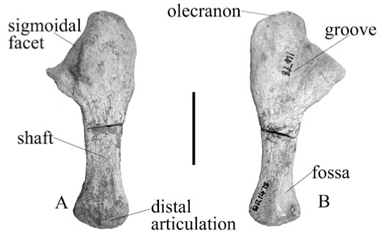
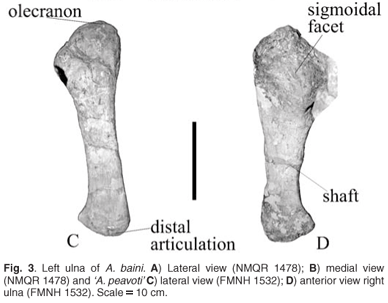
Anterior to the olecranon is a broad, anteriorly directed sigmoidal face in A. baini. It is open and slopes ventrally. Below the sigmoidal face is the shallow triangular radial facet, which is located along the anterior border just above the shaft.
The lateral border of the sigmoidal face projects above the lateral surface of the ulna, forming a broad, flat ridge in A. baini (Fig. 2C). The ridge is short and ends at the proximal end of the shaft. Posterior to this ridge is a narrow, shallow groove that does not extend far down the shaft. The shaft is medio-laterally flat and is antero-posteriorly narrow.
Distally, the ulna of A. baini (Fig. 3A) is antero-posteriorly elongated with a slightly convex articulating surface, which is more convex in the middle and slightly flat on either side. On the lateral surface there is a convex, slightly raised area forming a small, round tubercle.
On the medial surface of the proximal end of the ulna of A. baini there is a narrow, fairly deep groove that is more concave in the middle (Fig. 3B). In front of the groove the surface becomes raised and has a flat surface. Along the middle of the shaft is a shallow groove that extends to the distal end. On the anterior border of the shaft there is an elongated, almost oval depression on the bone that becomes deeper distally. The distal end of the depression is demarcated by a thin ridge.
Pelvic girdle and hindlimb
Ilium
The only element of the pelvic girdle of A. baini present is the ilium; the anterior process is completed in plaster. The blade is antero-posteriorly short with a wide anterior process and a narrow posterior process similar to that of 'A. peavoti' (FMNH 1532). In Rhachiocephalus (SAM-PK-K6446) the ilium blade is antero-posteriorly longer than that of A. baini and the anterior process is narrower and longer than that of A. baini (NMQR 1478) and 'A. peavoti' (FMNH 1532). The anterior process projects further anteriorly than that of A. baini and is located well in front of the acetabulum. The anterior process is located higher than the posterior process and is laterally everted in A. baini (Fig. 4A), whereas in 'A. peavoti' (Fig. 4B) the anterior and posterior processes are almost at the same level (FMNH 1532); the anterior process is only slightly higher in Rhachiocephalus. The dorsal border of the ilium blade is slightly rugose, and just below the middle of the dorsal border the blade has a concave surface. The ilium blade is expanded and fan-shaped in most dicynodonts, with the possible exception of Eodicynodon, which has a rectangular blade (NMQR 3155). The ilium is widely expanded in Robertia and Diictodon,4,5,10 whereas that of Kingoria is triangular15,16 and positioned well in front of the pubo-ischiadic plate,15 which is also the situation in Cistecephalus.11 The ilium of 'D.' trigonocephalus is a compact, flat, dorso-medially orientated plate.16 Medially, the ilium is convex and the surface is marked by the attachment sites for at least four sacral ribs.
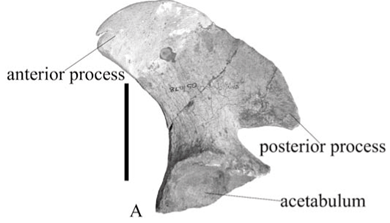
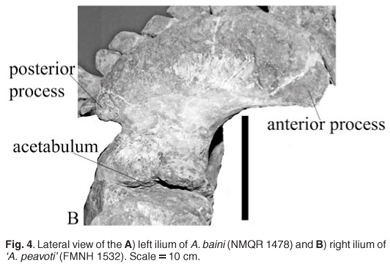
The ilium blade narrows towards the acetabulum to form a neck. It is short, broad and fairly robust. The neck of the ilium is also slightly longer and narrower in A. baini (NMQR 1478) than in 'A. peavoti' (FMNH 1532). Below the neck the acetabulum is restricted to the anterior two-thirds of the distal end of the ilium. It is almost circular with a rugose, concave surface that is directed postero-laterally. The supra-acetabular ridge is narrow and has a notch about two-thirds of the way posteriorly. The ischial articulation is larger than that for the pubis.
In 'A. peavoti' the pubis and ischium have also been preserved (FMNH 1532). The ischium has a triangular appearance, possibly contributing only a small portion of the acetabulum and the obturator foramen. In 'A. peavoti' the ischial contribution to the acetabulum faces slightly antero-laterally. Unfortunately, the pubis is obscured by the femur on the mounted specimen. The acetabulum in 'A. peavoti' faces ventrally as well as antero-laterally (FMNH 1532).
Hindlimb
Femur
The left femur of A. baini is a well-preserved, robust bone. It has a wide, long proximal end with a short wide shaft attaching to a narrow distal end (Fig. 5A, B). Similarly, the proximal end of the femur of 'A. peavoti' (Fig. 5C, D) forms a rectangle with a short wide shaft which grades into a wide distal end (FMNH 1532); however, the femur of Rhachiocephalus (SAM-PK-K6446) is more curved than that of A. baini. The head of the bone is oval, medially inflected but is not continuous with the greater trochanter in A. baini (NMQR 1478). It is raised slightly above the surface of the bone and is separated from the greater trochanter by a fossa in front of the head (NMQR 1478). The shape of the bone is similar to that of A. baini, which leads one to suggest that shape of the head most likely is oval. The head of the femur of 'A. peavoti' (FMNH 1532) and of Rhachiocephalus (SAM-PK-K6446), however, is continuous with the greater trochanter. The femoral head of A. baini also is more distinctly developed than that of Rhachiocephalus (SAM-PK-K6446), and its articulating surface is directed antero-dorsally in A. baini, whereas it is directed more anteriorly in Rhachiocephalus (SAM-PK-K6446).
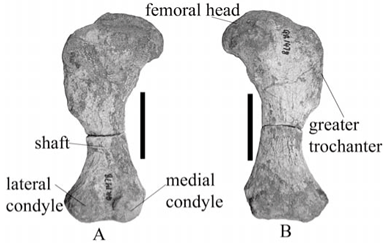
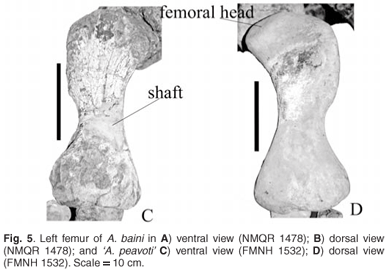
The greater trochanter of A. baini is wide proximally and narrows in a ventral direction. It has a rugose surface, and is parallel to the long axis of the bone, which is similar to that of 'A. peavoti' (photographs of FMNH 1532). In Rhachiocephalus the greater trochanter is wider and more rugose than that of A. baini (NMQR 1478). Below the greater trochanter the bone narrows to form the shaft, which is narrower in A. baini than 'A. peavoti'. It has a concave lateral border and a straighter medial border in both A. baini (NMQR 1478) and 'A. peavoti' (FMNH 1532). Ventrally, the minor trochanter of A. baini forms a small, elongated oval tubercle. The intertrochanteric fossa is small and has an irregular shape.
Distally, the articulating condyles are small but are still raised above the surface of the bone in A. baini (Fig. 5A, B). The ventral articulating surfaces of the condyles are at the same level. Ventrally, the surfaces of the articulating condyles show some damage due to weathering. The intercondylar fossa is slightly wide and deep. Above the condyles is the popliteal fossa, which is almost circular and fairly deep.
Among the Permian dicynodonts there is much variation in the shape of the femur. Eodicynodon has a gracile femur with a triangular head that is continuous with the greater trochanter. In Robertia it is sigmoidal with a distinct shaft, and the head of the femur is located along the anterior margin and extends dorsally.10 Diictodon has a moderately convex head that is angled dorsally and medially, and it merges with the greater trochanter.4 The head of the femur extends dorso-ventrally and is offset slightly from the rest of the bone in Diictodon.5 A different morphology is seen in Kingoria, where the proximal half of the femur forms a flat plate and the proximal extremity has a curved appearance.16 In Kingoria the femoral head extends dorsally and ventrally, and is offset from the shaft.15,16 The head is continuous with the greater trochanter and there is no evidence of an internal or fourth trochanter.3 In Cistecephalus the head of the femur is well-defined and carried medially.11 The greater trochanter is located on the lateral 'corner', does not extend far down the shaft, and is separated from the head by a groove.11 The femur of D. trigonocephalus is dorso-ventrally flattened with the head located along the anterior margin. It encroaches on the dorsal surface and the greater trochanter forms a plate.3
Tibia
Both the left and right tibiae of A. baini (NMQR 1478) are preserved and are complete. The lateral border is convex whereas the medial is concave (Fig. 6A, B, D, E). In proximal view the articulating surface has a triangular appearance. Proximally, the tibia of 'A. peavoti' (Fig. 6C, F) is more widely expanded than that of A. baini. Posterior to the tibial process are two concave articulating facets with the medial one larger and slightly more concave. The tibial process is not very prominent but is an elongated oval with a rugose convex surface (Fig. 6A, B).

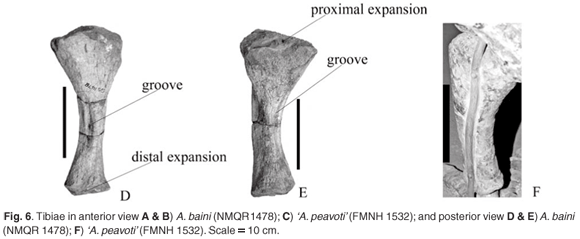
Proximally on the lateral border of the tibia of A. baini is an oval groove that is wide proximally and narrows distally. Along the medial border it becomes flat to slightly concave. The distal end is slightly concavo-convex.
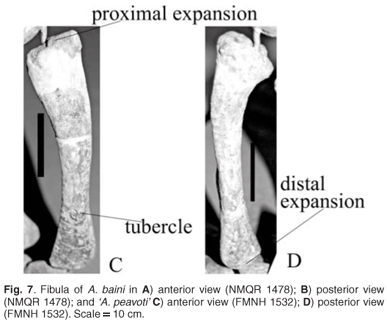
Fibula
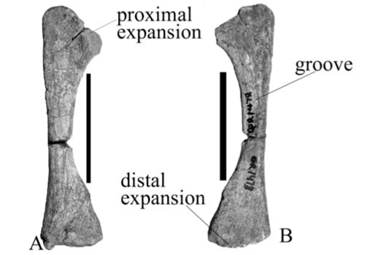
The fibula of A. baini is a gracile bone with slightly expanded ends and a narrow shaft (Fig. 7A, B). Along the anterior surface of the shaft is a low broad ridge that extends from the proximal end of the shaft to its distal end closer to the lateral border in A. baini than in 'A. peavoti' (Fig. 7C, D). On the anterior surface of the distal end of the fibula of 'A. peavoti' a prominent, oval tubercle is present. The medial border is deeply concave and the lateral border is straight in both A. baini and 'A. peavoti'.
Discussion
An analysis of the postcranial skeleton of Aulacephalodon baini and a comparison with 'Aulacephalodon peavoti' has shown that these two taxa have similarities but also that there are some significant differences. Recent analyses of the skeleton of 'A. peavoti' has raised questions as to its identification; moreover, the holotype material has been misplaced (Defauw;4 Angielcyzk, pers. comm. 2007; 2008). The scapula blade of A. baini is more curved than that of 'A. peavoti', whose blade is more upright. The delto-pectoral crest forms a rectangle in A. baini and is semi-circular in 'A. peavoti'. In A. baini the articulating surface of the glenoid is flat to concavo-convex, while that of 'A. peavoti' is concave. Another striking difference is that the coracoid foramen is positioned on the precoracoid in A. baini whereas it is located between the coracoid and scapula in 'A. peavoti' as seen in some Triassic dicynodonts (e.g. Ischigualastia). The ulna of A. baini is more robust than that of 'A. peavoti' and the proximal end of the ulna is longer in A. baini, which results in the shaft being shorter. In A. baini the olecranon is better developed and more prominent than in 'A. peavoti'. In A. baini the anterior process of the ilium is positioned higher than the posterior process as compared with 'A. peavoti', where the anterior and posterior processes are almost at the same level. The head of the femur is separated from the greater trochanter in A. baini and is continuous with the greater trochanter in 'A. peavoti'. The femoral shaft and distal end are narrower in A. baini than 'A. peavoti'.
There are also similarities between the skeletal elements. Neither taxon has a scapula spine present. The olecranon fossa in A. baini and 'A. peavoti' forms a very shallow concave surface. There are similarities in the femora of these taxa. The proximal expansion is rectangular in both and the greater trochanter is parallel to the long axis of the bone. In A. baini and 'A. peavoti' (based on the right femur) the femoral head is separated from the greater trochanter and raised above the surface of the bone; the articulating surface is directed antero-dorsally.
Another point of interest is a comparison with Rhachiocephalus magnus, which shows that there are significant differences between it and A. baini and 'A. peavoti'. Rhachiocephalus has a wider scapula blade than either of the other two. The glenoid facet is elliptical and has a convex articulating surface compared with the concavo-convex articulating surface in A. baini and a concave surface in 'A. peavoti'. The humeral head also shows variation in shape, from round in the small individual of A. baini and 'A. peavoti' to square shaped in the large specimen of A. baini to hemispherical in Rhachiocephalus. The olecranon is better developed in A. baini than in 'A. peavoti' and R. magnus. In the ilium the anterior process is distinctly higher than the posterior process in A. baini, whereas it is lower in R. magnus and is more or less at the same level in 'A. peavoti'. The acetabulum is directed laterally in A. baini and 'A. peavoti' while in R. magnus it is directed more ventrally and the lateral wall of the acetabulum projects ventrally. The femoral head appears not to project above the surface of the bone as it does in A. baini and 'A. peavoti' and, as in the latter, it is continuous with the greater trochanter in R. magnus.
Systematic palaeontology
Synapsida Osborn, 1903
Therapsida Broom, 1905
Anomodontia Owen, 1859
Dicynodontia Owen, 1859
Aulacephalodon baini(Owen 1844) Seeley 1898
Diagnosis: cranial diagnosis as in King.1 Medium-sized, tusked dicynodont with wide intertemporal region. Parietals exposed on skull roof. Pineal foramen surrounded by a boss. Postorbitals in the intertemporal region steep-sided and drawn up into a ridge dorsally. Postorbitals overhang side wall of braincase, forming recess beneath skull roof on each side. Boss on jugal of zygoma. Jugal has tall dorsal process extending far up postorbital bar. Prefrontal and nasal bosses. No postfrontal bone. Nostril deeply recessed with definite posterior margin. Ridge on maxilla posterior to nostril. Interpterygoid vacuity moderately long. Ventral edge of vomerine plate without trough. Palatines bulbous. No postcaniniform crest. Massive dentary symphysis with dentary tables. Lateral dentary shelf occludes mandibular fenestra. Dentary sulcus present.
A. baini differs from 'A. peavoti' in that it has a narrower, more curved scapula blade. Laterally, on the proximal third of the scapula blade is a shallower fossa. Small, narrow acromion process. Glenoid facet is round and concavo-convex in the adult A. baini and almost flat in the juvenile. Glenoid is directed postero-laterally. Precoracoid is wedge shaped. Triangular coracoid. Glenoid is directed caudo-laterally and more open. Square humeral head. Delto-pectoral crest is rectangular and is directed anteriorly. Longer proximal end with a more prominent olecranon. The anterior process is positioned higher than the posterior process. Femoral head is separated from the greater trochanter. The shaft and distal end are narrower. Tibia is not as widely expanded proximally. Fibula does not have tubercle distally on the shaft.
Rhachiocephalus differs from A. baini in that it has a wider scapula blade. Dorsal third of the scapula blade is almost flat. Glenoid facet of the scapula is elliptical and has a convex surface. Coracoid foramen is located between the scapula and precoracoid. Coracoid is 'axe-shaped'. Proximal expansion of the humerus is wider than the distal one. Humeral head is hemispherical. Poorly developed olecranon. Femoral head continuous with greater trochanter.
Odontocyclops differs from A. baini in that the scapula blade is spatulate. A well-developed scapula spine is present. Distal expansion is wider than proximal in the humerus. Humeral head is subhemispherical.
A. baini is similar to 'A. peavoti' in that there is no scapula spine present. Proximal expansion of humerus is rectangular. Ilium acetabular facet is directed ventrally. Head of the femur is similar in that it is oval and directed dorsally. Greater trochanter is parallel to the long axis of the bone.
Aulacephalodon peavoti Broom 1921
Diagnosis ('A. peavoti'): Cranial diagnosis as in Broom.7 Snout and the whole front half of skull flatter than other anomodonts. Premaxilla small and unusually flat. Nasals are short but very broad and each has a well-developed boss which overhangs nostrils. Small septomaxilla. Maxilla is shallow. No tusks. Prefrontal small triangular bone that has a small prominent boss above orbit. Small lacrimal. Large jugal. Large wide frontal bone articulates with postorbital but is separated from frontal by postfrontal. Postfrontal scarcely visible on surface. Postorbital unusually large.
'A. peavoti' differs from A. baini: it has a wider, more upright scapula blade. On the proximal third of the scapula's lateral surface is a deeper fossa. Scapula glenoid facet is oval and concave. Coracoid foramen is located between the scapula and the precoracoid. Delto-pectoral crest is semi-circular. Ulna has a poorly developed olecranon. Longer shaft. Oval femoral head is continuous with the greater trochanter. Proximal end of the tibia is more widely expanded. Tubercle present on the distal end of fibula shaft above distal expansion.
Rhachiocephalus differs from 'A. peavoti'. It has a wider scapula blade. The proximal third of the scapula blade is almost flat. Glenoid facet is elliptical with a convex articulating surface. Humeral head is hemispherical. Anterior process of the ilium is higher than the posterior process. Acetabulum is directed more ventrally.
Remarks. Recent studies of Aulacephalodon resulted in the number of species being reduced from seventeen to one including 'A. peavoti', which had been synonymised with A. baini and referred to the genus Rhachiocephalus.4 Examination of the material has led to renewed questions about 'A. peavoti's taxonomic position as it possibly resembles Pelanomodon or Odontocyclops, which remains uncertain until it is re-examined (K.D. Angielczyk, pers comm. 2007). Confusion as to where 'A. peavoti' belongs taxonomically is compounded by the missing holotype. There is currently a fairly complete specimen of Odontocyclops housed at the South African Museum in Cape Town, which to date remains undescribed. It does, however, provide the opportunity for more detailed comparison of Odontocyclops with other taxa.
Conclusion
A. baini and 'A. peavoti' have a number of similarities in their postcranial anatomy and are differentiated by several significant differences. Whether or not A. baini is the only species is dependent on the findings of the re-evaluation of the skull of 'A. peavoti' (FMNH 1532). A comparison with Rhachiocephalus shows a number of differences with both A. baini and 'A. peavoti'. The results of the analysis of the postcranial skeleton will, one hopes, provide some useful information should there be detailed investigation of the identification and taxonomic position of 'A. peavoti'.
I would like to thank the National Research Foundation for funding the postdoctoral fellowship that resulted in this work. I am grateful to Billy de Klerk and Elize Butler for all their help as well as for allowing me access to the Aulacephalodon material in their respective collections. I am extremely grateful to Ken Angielczyk for all his help with the writing of this paper as well for taking the time to review material housed at the Field Museum in Chicago. I would also like to express my gratitude to the two reviewers of the manuscript for their helpful comments.
1. King G.M. (1988). Anomodontia. Encyclopedia of Paleoherpetology 17C, 64–171. [ Links ]
2. King G.M. (1990). Life and death in the Permo-Triassic: the fortunes of the dicynodont mammal-like reptiles. Sidney Haughton Memorial Lecture 3, 1–17. Royal Society of South Africa, Cape Town. [ Links ]
3. King G.M. (1981). The functional anatomy of a Permian dicynodont. Phil. Trans R. Soc. B 291, 243–322. [ Links ]
4. Defauw S.L. (1986). The appendicular skeleton of African dicynodonts. Ph.D. thesis, Wayne State University, Detroit, Michigan. [ Links ]
5. Ray S. and Chisamy A. (2003). Functional aspects of the postcranial anatomy of the Permian dicynodont Diictodon and their ecological implications. Palaeontology 46(1), 151–183. [ Links ]
6. Tollman S.M., Grine F.E. and Hahn B.D. (1980). Ontogeny and sexual dimorphism in Aulacephalodon (Reptilia; Anomodontia). Ann S. Afr. Mus. 81(4), 159–186. [ Links ]
7. Broom R. (1921). On some new genera and species of anomodont reptiles from the Karoo beds of South Africa. Proc. Zool. Soc. Lond. XLIV, 44–674. [ Links ]
8. Olsen E.C. and Byrne F. (1938). The osteology of Aulacephalodon peavoti Broom. J. Geol. LXVI, 177–109 [ Links ]
9. Rubidge B.S., King G.M. and Hancox P.J. (1994). The postcranial skeleton of the earliest dicynodont synapsid Eodicynodon from the Upper Permian of South Africa. Palaeontology 37(2), 397–408. [ Links ]
10. King G.M. (1981). The postcranial skeleton of Robertia broomiana, an early dicynodont (Reptilia, Therapsida) from the South African Karoo. Ann. S. Afr. Mus. 84(5), 203–231. [ Links ]
11. Cluver M.A. (1978). The skeleton of the mammal-like reptile Cistecephalus. Ann. S. Afr. Mus. 76(5), 213–245. [ Links ]
12. Cox C.B. (1972). A new digging dicynodont from the Upper Permian of Tanzania. In Studies in Vertebrate Evolution, eds K.A. Joysey and T.S. Kemp, pp. 173–189. Oliver & Boyd, Edinburgh. [ Links ]
13. Angielczyk K.D. (2002). Redescription, phylogenetic position, and stratigraphic significance of the dicynodont genus Odontocyclops (Synapsida: Anomodontia). J. Paleontol. 76(6), 1047–1059. [ Links ]
14. Angielczyk K.D. (2007). New specimens of the Tanzanian dicynodont 'Cryptocynodon' parringtoni Von Huene, 1942 (Therapsida, Anomodontia), with an expanded analysis of Permian dicynodont phylogeny. J. Vertebr. Paleontol. 27(1), 116–131. [ Links ]
15. Cox C.B. (1959). On the anatomy of a new dicynodont genus with evidence of the position of the tympanum. Proc. Zool. Soc. Lond. 132, 321–367. [ Links ]
16. King G M. (1985). The postcranial skeleton of Kingoria nowacki (von Huene) (Therapsida: Dicynodontia). Zool. J. Linn. Soc. 84, 263–289. [ Links ]
17. Maisch M. (2004). Postcranial morphology of Rhachiocephalus Seeley, 1898 (Therapsida: Dicynodontia) from the Upper Permian of Tanzania and the status of Platypodosaurus robustus Owen, 1880. Geol. et Palaeont. 38, 161–175. [ Links ]
Received 29 February. Accepted 14 November 2008.
Institutional abbreviations
AM, Albany Museum, Grahamstown, South Africa.
NMQR, National Museum, Bloemfontein, South Africa.
FMNH, Field Museum of Natural History, Chicago, U.S.A.
SAM-PK, Iziko South African Museum, Cape Town, South Africa.

