










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.104 n.9-10 Pretoria Sep./Oct. 2008
RESEARCH LETTERS
The earliest primate (Parapapio sp.) from the Cradle of Humankind World Heritage site (Waypoint 160, Bolt's Farm, South Africa)
D. GommeryI, II, *; J.F. ThackerayII; F. SénégasI, II; S. PotzeII; L. KgasiII
IUPR 2147 CNRS, 44 rue de l'Amiral Mouchez, 75014, Paris, France
IIHuman Origins and Past Environments Research Unit, Transvaal Museum, P.O. Box 413, Pretoria 0001, South Africa
ABSTRACT
Waypoint 160 is a breccia occurrence in a dolomite solution cavity on Bolt's Farm on the southern margin of the Cradle of Humankind World Heritage site in South Africa. The calcified breccia has yielded microfauna that has been dated between 4.0 and 4.5 million years. Here we present the first evidence for Parapapio, an extinct monkey, from Waypoint 160. This site is of particular interest as Parapapio is associated with Pliocene or Pleistocene hominids in Africa, for example at Sterkfontein, Taung, Kromdraai, Swartkrans and Makapansgat in South Africa, as well as at Lothagam in East Africa.
Introduction
Several sites on Bolt's Farm have yielded an interesting macrofauna (including primates and non-primates) of Plio-Pleistocene age.1–4 Waypoint 160 is the oldest site on Bolt's Farm. It was discovered by B. Senut, M. Pickford and J. Michaux in 1996.4 The microfaunal assemblage from a dump discarded by lime miners about 60 years ago5,6 is slightly younger (4–4.5 Myr) than the microfauna from Langebaanweg (around 5 Myr), which also yielded remains of Parapapio.7
Macrofaunal fossils are rare at Waypoint 160, which has been a focus of fieldwork and research conducted by French and South African members of the HOPE (Human Origins and Past Environments) programme, based at the Transvaal Museum in Pretoria. Microfaunal taxa indicate local palaeoenvironments,5,6,8 whereas the macrofauna serves to reflect regional palaeoenvironments that prevailed between 4 and 4.5 million years ago. Hominids certainly existed within this period in Kenya, notably at Lothagam9 and in the Baringo Basin.10 The question arises as to whether hominids also existed in southern Africa during the same period. Waypoint 160 offers the potential opportunity to explore this possibility.
Materials
We report on fossils from two blocks of breccia from Waypoint 160. WP02-2 was collected on 16 August 2002 by D. Gommery and J.F. Thackeray, and was prepared in acetic acid by F. Sénégas. It yielded several primate postcranial bones that probably belong to a single individual, and some microfaunal remains that were used to estimate an age between 4 and 4.5 Myr. All the primate specimens from this block of breccia (Figs 1 & 2) have the prefix WP, including a humerus (WP 1), a femoral shaft (WP 2), a patella (WP 3), a thoracic vertebra dorsal arch (WP 4) and a rib (WP 5). The second block, WP 07-10, was collected on 15 May 2007 by D. Gommery, J.F. Thackeray and S. Potze, and includes a molar fragment and the impression of part of a crown of a tooth. The tooth is catalogued as WP 6 (Fig. 3).



These postcranial and cranial elements are all attributed to Parapapio sp., and are compared with extant primate skeletons housed at the Transvaal Museum. The fossils are also compared with two well-preserved fossil monkey bones (SK 591A and C from Swartkrans (Brain 1981) curated at the Palaeontology Department of the Transvaal Museum). SK 591A and C can be attributed to Parapapio. SK 591 A is a left femur with the proximal end and shaft. SK 591C is a complete left humerus. The morphology of SK 591A is comparable to KNM-LT 2974 from Lothagam9 and A.L.363-1d from Hadar,11 which have both been attributed to the genus Parapapio. SK 591C is comparable to KNM-LT 23074, 23077 and 416 from Lothagam,9 and to A.L.363-12 from Hadar,11 also identified as Parapapio.
The following extant specimens were examined for purposes of comparison with the Waypoint 160 primate postcrania (M = male, F = female, R = right and L = left):
• Papio ursinus (baboon): AZ/191 (M), AZ/767 (F), AZ/2921 (?M), AZ/147 R & L (M)
• Mandrillus sphinx: AZ/1427 (F), AZ/1971 (F), AZ/1972 (F)
• Cercopithecus albogularis: AZ/1050 R & L (M)
• Cercopithecus mitis (samango monkey): AZ/328 R & L (M), AZ/779 R & L (M), AZ/327 R & L (F)
• Chlorocebus aethiops pygerythrus (vervet monkey): AZ/587 L, AZ/645 R (M), AZ/576 R & L (F), AZ 575 R & L (M), AZ/990 R & L (F), AZ/759 R & L (M)
• Colobus guereza: AZ/981 R & L (M), AZ/1437 R & L (M)
Results
WP 1 (Figs 1 & 2) is a left humerus with a preserved length of 133.8 mm, similar in size to the humeri of vervet or samango monkeys. The articular ends of WP 1 are missing because they were unfused, indicating that the individual was juvenile at the time of death. Most of the anterior part is broken.
In anterior (Fig. 1) and posterior (Fig. 2) views, the shaft of WP 1 is straight, as in the humeri of SK 591C, Papio, Mandrillus, Cercopithecus albogularis and Colobus. In vervet and samango monkeys, the shaft is curved medially. In medial and lateral view, the humerus is slightly curved posteriorly in its proximal first third as in baboons and SK 591C. It is more curved in the first half in vervet and samango monkeys. The shaft is almost straight in Colobus guereza and Mandrillus sphinx.
In posterior view (Fig. 2) , the shaft is well preserved with a convex appearance on the superior fragment, and flat inferiorly. Superiorly, the pillar under the head is situated medially as in cercopithecids. This pillar has a more central position in Colobus guereza. The robust aspect of the pillar is similar to what is seen in Mandrillus. Distally, the crest of the supinator is straight and robust. This morphology is similar to that of baboons. The proximal part of the olecranon fossa is preserved and the border is inclined as in baboons but less so than in Mandrillus. The medial supracondylar ridge is complete and robust. The distal shaft and the lateral epicondyle are preserved.
In medial view, the lesser and greater tuberosities are not fused. A deep and moderately long fossa (12.4 mm long and 1.5 mm broad) is present below the beginning of the lesser tuberosity and on the Crista tuberculi minoris. This fossa corresponds to the point of insertion of M. teres major. It is situated in a position similar to that which is observed in SK 591C from Swartkrans, very close to the femoral head and close to the proximal position of the maximum development of the Crista tuberculi major. In other primates, where the insertion of M. teres major exists, it looks more like a tubercule.
Posteriorly to the insertion of M. teres major, a small crest exists (9.5 mm long) and probably corresponds to the insertion of the M. corabrachialis caput profondus.
The medial border associated with the Crista tuberculi minus is well defined on the distal part of the shaft, as in SK 591C. This border is less marked in Cercopithecus albogularis and Colobus, and more blunt in Papio and Mandrillus.
In lateral view, just a part of the shaft is preserved. The shaft is thick, as in Papio and Mandrillus, and different from the humerus shaft of Cercocopithecus. The shaft is not flattened as in vervet or in samango monkeys. The crest is straight. The border of the crest is thick and robust, more so than in SK 591C. It is similar to KNM-LT 23074 from Lothagam. In comparison with the maximum length of the humerus, the crest is longer in Colobus and shorter in other primates compared here. The crest of WP 1 is similar to that of Papio, but less developed because the Waypoint 160 primate is smaller.
In conclusion, WP 1 has many similarities with SK 591C, attributed to Parapapio, but is more robust.
WP 2 is a proximal fragment of a left femoral shaft 96.8 mm long (Figs 1 & 2). In anterior view (Fig. 1), close to the lateral border, two depressions exist. A small depression (7.7 mm long) is located proximally and corresponds probably to the end of the gluteal tuberosity. The longer depression is situated on the distal part (37.4 mm long) and is associated with a rounded and robust crest. The fossa and this crest correspond to the linea aspera. The medial ridge of the linea aspera is poorly marked in baboons.11 The fossa is deep as in juvenile or old specimens and, in baboons, is the place of the insertion of M. adductor magnus.11 In posterior view, the shaft surface is very convex medio-laterally, as in SK 591A.
WP 3 is an almost complete right patella (Figs 1 & 2). The lateral border of the distal part is broken. It is 17.9 mm in length, 14.3 mm in breadth and 8.3 mm thick. The characteristic of this bone in Catarrhini is its lengthening in the vertical direction.12 The patella is less elongated than in baboons, but more so than in vervet monkeys.
In anterior view, the medial part is convex and short medio-laterally. The lateral part is less convex and relatively large. In posterior view, the medial facet is high and short. This facet is concave proximo-distally. The lateral facet is low and large with an almost flat aspect in comparison with the medial facet. Distally, the apex is low, large and has a rounded aspect. The two facets have a morphology similar to that found in baboon patellae, and completely different from the situation that occurs in Colobus. The proximal part of the patella is much less developed than the distal part. Proximally, a superficial groove is observed and corresponds to the insertion of the lateral and medial Vasti and the M. rectus femoris. The morphology of the proximal part is closer to that of the vervet monkey. It is flat in Papio and Mandrillus, whereas it is rounded in Colobus.
WP 4 is a fragment of a thoracic vertebra dorsal arch (20.7 mm long, 23.3 mm wide) (Figs 1 & 2). The spinous process is present and well preserved. The left lamina is preserved together with the inferior articular facet seen in anterior view. This facet is elongated medially to laterally (5.1 mm long and 7.5 mm broad). The articular surface is concave from medial to lateral. The robust transverse process is preserved but is eroded at the tip. The beginning of the costal facet is present in superior view. This specimen could be the first thoracic vertebra.13
WP 5 is the proximal part of a rib (28.1 mm long) (Figs 1 & 2). The neck is long and well developed. The head is eroded. The tubercle is broken but a crest starts on the origin and separates the surface in two parts as described by Olivier & Laffont14 for a first rib. The proximal portion of the costal body is 6.2 mm in length, and slightly curved. This fossil is probably a left first rib.
Dentition
WP 6 is a broken primate tooth with the partial mold of the crown (Fig. 3). Some enamel is preserved on the lingual border. The mold of the crown is almost square. The tooth is probably a right upper M1 or a right upper M2.
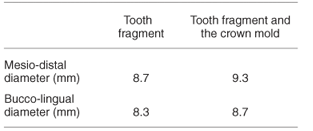
It is difficult to make direct comparisons between the dimensions of WP 6 and other specimens of Parapapio, but the dimensions of the tooth from Waypoint 160 are very similar to the second molar of Parapapio lothagamensis.9
Discussion and conclusions
Postcranial bones of Pliocene cercopithecoid primates are rare in sub-Saharan sites, and difficult to attribute to particular taxa.9,15–19 The difficulty of identifying Parapapio in South Africa is complicated by the fact that isotopic and metric analyses suggest that specimens attributed to Parapapio jonesi might include female specimens of Parapapio broomi.20
The primate postcranial bones from the block of breccia WP 02-2 (humerus, femur and patella) have cercopithecid affinities, especially with papionins. The bones are heavily built, suggesting a terrestrial or a semi-terrestrial adaptation. The associated microfauna at Waypoint 1605,6,8 suggests an open environment that was drier than that of today. The robust postcranial bones of the primate from Waypoint 160 may be related to such environments. El-Zaatari et al.21 concluded that the dietary habits of some species of Parapapio reflect temporal variability, but spatial variability in southern Africa also needs to be considered. The Parapapio from Waypoint 160 is probably different from the Parapapio represented in the upper phase breccia of Makapansgat Limeworks,1,2,22,23 associated with humid and closed palaeoenvironments. Some species of Parapapio may have been frugivorous, while others may have consumed hard food in dry and open habitats, as for example at Waypoint 160.
The morphological patterns of the Waypoint 160 primate postcranial bones, described here, indicate that it is referable to the genus Parapapio. This is the oldest known primate in the Cradle of Humankind World Heritage site. Because Parapapio is known to co-occur with hominids in both South Africa and East Africa, we emphasize that Waypoint 160 deserves attention as a potential hominid site, penecontemporary with Lothagam, Chemeron and Tabarin in East Africa.
This study is part of the Human Origins and Past Environments programme, supported by the PICS CNRS-NRF project entitled 'Plio-Pleistocene hominids and palaeoenvironments of the Cradle of Humankind (South Africa)', the PAI PROTEA French-South African Palaeoanthropology and Palaeoenvironmental Studies in South Africa, financed by the National Research Foundation in South Africa, and Ministries for Research and Foreign Affairs in France, the Transvaal Museum and UPR 2147 of the CNRS. We are grateful to the French Embassy in South Africa for support. We would like to thank J. Klinkert, J. Gaylord and M. Gaylord for permission to access sites on Bolt's Farm, M. Leslie (South African Heritage Resource Agency) for the SAHRA permit, as well as A. Corval (head of the CNRS Office for Sub-Saharan Africa, Johannesburg), S. Elmaleh (Embassy of France in South Africa), T. Perregil, E. Masango and T. Kearney (Transvaal Museum) for their kind assistance.
1. Freedman L. (1957). The fossil Cercopithecoidea of South Africa. Ann. Transv. Mus. 23, 121–262. [ Links ]
2. Freedman L. (1965). Fossil and subfossil primates from the limestone deposits at Taung, Bolt's Farm and Witkrans, South Africa. Palaeont. afr. 9, 19–48. [ Links ]
3. Cooke H.B.S. (1991). Dinofelis barlowi (Mammalia, Carnivora, Felidae) cranial material from Bolt's Farm, collected by the University of California African Expedition. Palaeont. afr. 28, 9–21. [ Links ]
4. Sénégas F., Thackeray J.F., Gommery D. and Braga J. (2002). Palaeontological sites on 'Bolt's Farm', Sterkfontein Valley, South Africa. Ann. Transv. Mus. 39, 65–67. [ Links ]
5. Sénégas F. and Avery M. (1998). New evidence for the murine origins of the Otomyinae (Mammalia, Rodentia) and the age of Bolt's Farm (South Africa). S. Afr. J. Sci. 94, 503–507. [ Links ]
6. Sénégas F. and Michaux J. (2000). Boltimys broomi gen. nov., sp. nov. (Rodentia, Mammalia), nouveau Muridae d'affinité incertaine du Pliocène inférieur d'Afrique du Sud. C. R. Acad. Sci. Paris 330, 521–525. [ Links ]
7. Grine F.E. and Hendey Q.B. (1981). Earliest primate remains from South Africa. S. Afr. J. Sci. 77, 374–376. [ Links ]
8. Sénégas F. (2004). A new species of Petromus (Rodentia, Hystricognatha, Petromuridae) from the early Pliocene of South Africa and its paleoenvironmental implications. J. Vert. Paleont. 24, 757–763. [ Links ]
9. Leakey M., Teaford, M. and Ward C. (2003). Cercopithecidae from Lothagam. In Lothagam: the Dawn of Humanity in Eastern Africa, eds M.G. Leakey and J.M. Harris , pp. 202–248. Columbia University Press, New York. [ Links ]
10. Hill A. (1999). The Baringo Basin, Kenya: from Bishop to BPRP. In Late Cenozoic Environments and Hominid Evolution: a Tribute to Bill Bishop, eds P. Andrews and P. Banham, pp. 85–97. Geological Society, London. [ Links ]
11. Swindler D.R. and Wood C.D. (1973). An Atlas of Primate Gross Anatomy. Baboon, Chimpanzee, and Man. University of Washington Press, Seattle. [ Links ]
12. Olivier G. and Piganiol G. (1957). Le fémur du Semnopithèque. Mammalia 21, 430–451 [ Links ]
13. Olivier G. and Gaillard J. (1964). Les vertèbres du Semnopithèque. Mammalia 28, 476–518 [ Links ]
14. Olivier G. and Laffont J. (1962). Le squelette thoracique du Semnopithèque. Mammalia 26, 431–446. [ Links ]
15. Frost S. (2001). New Early Pliocene Cercopithecidae (Mammalia: Primates) from Aramis, Middle Awash Valley Ethiopia. Am. Mus. Novitates 3350, 1–36. [ Links ]
16. Frost S. and Delson E. (2002). Fossil Cercopithecidae from the Hadar Formation and surrounding areas of the Afar Depression, Ethiopia. J. Hum. Evol. 43, 687–748. [ Links ]
17. Senut B. (1994). Cercopithecoidea néogènes et quaternaries du rift occidental (Ouganda). Geology and Palaeobiology of the Albertine Rift Valley, Uganda-Zaire, Vol. II: Palaeobiology, pp. 195–205. Cifeg Occasional Publications, Orléans. [ Links ]
18. Elton S. (2001). Locomotor and habitat classifications of cercopithecoid postcranial material from Sterkfontein Member 4, Bolt's Farm and Swartkrans Members 1 and 2, South Africa. Palaeont. afr. 37, 115–126. [ Links ]
19. Williams F.L., Ackermann R.R. and Leight S.R. (2007). Inferring Plio-Pleistocene southern African biochronology from the facial affinities in Parapapio and other fossil Papionins. Am. J. Phys. Anthropol. 132, 163–174. [ Links ]
20. Thackeray J.F. and Myer S. (2004). Parapapio broomi and Parapapio jonesi from Sterkfontein: males and females of one species? Ann. Transv. Mus. 41, 79–82. [ Links ]
21. El-Zaatari S., Grine F., Teaford M. and Smith H. (2005). Molar microwear and dietary reconstructions of fossil Cercopithecoidea from the Plio-Pleistocene deposits of South Africa. J. Hum. Evol. 49, 180–205. [ Links ]
22. Freedman L. (1960). Some new cercopithecoid specimens from Makapansgat, South Africa. Palaeont. afr. 7, 7–45. [ Links ]
23. Maier W. (1970). New fossil Cercopithecoidea from the lower Pleistocene cave deposits of Makapansgat Limeworks, South Africa. Palaeont. afr. 13, 69–107. [ Links ]
Received 5 August. Accepted 16 October 2008.
* Author for correspondence. E-mail: dominique.gommery@evolhum.cnrs.fr

