










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.104 n.9-10 Pretoria Sep./Oct. 2008
RESEARCH IN ACTION
First hominoid from the Late Miocene of Niger
Martin PickfordI, II, *; Brigitte SenutII; Jorge MoralesIII; José BragaIV
ICollège de France, 11 place M. Berthelot, 75005, Paris, France
IIUMR 5143 du CNRS, Case postale 38, 8 rue Buffon, 75005, Paris, France
IIIPaleobiologia, Museo Nacional de Ciencias Naturales, CSIC, José Gutierrez Abascal 2, 28006, Madrid, Spain
IVUniversité Paul Sabatier, Toulouse, FRE 2960 CNRS, Anthropobiologie et Imagerie Anatomique, 39 Allées Jules Guesde, 31000, Toulouse, France
ABSTRACT
Since the discovery of Orrorin tugenensis in the Late Miocene Lukeino Formation, Tugen Hills, Kenya, it is generally admitted that the origin of bipedal hominids occurred earlier than 6 Myr ago and that the adaptation to bipedal stance and locomotion initially occurred in a forested or well-wooded setting. In Africa, eight localities aged between 13 and 5.5 Myr have yielded hominoid fossils belonging to nine species. We here report the occurrence of a Late Miocene hominoid in Niger, associated with a restricted fauna which indicates an age of c. 11–8 Myr. The Niger fossil locality is 940 km north of the nearest known extant hominoids, and 1000 km west of the nearest recorded fossil hominoid from Chad. The scientific value of the Niger specimen resides in its discovery locus far from any other known fossil hominoids, its Late Miocene age and the attention that it will focus on the Neogene fossil record of West Africa, currently almost unknown.
Introduction
It is now widely accepted that the family Hominidae originated in Africa during the Late Miocene. However, the African fossil record for this time period is not well endowed with hominoid fossils, making it difficult to trace the transition from pre-hominid to hominid status. Since 1997, nine hominoid taxa have been reported from eight localities in Africa, ranging in age between 13 and 5 Myr: one species in Namibia [Otavipithecus (12–13 Myr)]; five taxa in Kenya14,25,28 [a chimp-like hominoid (12.5 Myr), Nakalipithecus (9.5–10 Myr), Samburupithecus (9.5 Myr), a gorilla-like hominoid (6 Myr), and Orrorin (6 Myr)]; two in Ethiopia29 [Chororapithecus (10–10.5 Myr) and Ardipithecus (5.7 Myr)]; and one in Chad3 [Sahelanthropus (6–10 Myr)23]. Despite the drawbacks of its Late Miocene fossil record, Africa has now yielded a respectable diversity of hominoids comparable with that of Europe and Asia, making it more likely to be the cradle of hominid evolution than Eurasia.1 All these African Late Miocene taxa are poorly known, and a considerable amount of debate has taken place, even concerning some relatively well-preserved specimens.3 Each addition to the African Late Miocene hominoid fossil sample is therefore precious.
In 1964, a small collection of vertebrate fossils from Niger, which includes a fragment of hominoid mandible, was sent to the Muséum National d'Histoire Naturelle in Paris by the Bataafse Internationale Petroleum Maatshappij (now Shell), where it is curated under register No. 1964-27.885. The precise point of discovery is not known, but it is probably close to 5°43'E, 15°32'N, where some Mesozoic Chelonians2 with the register No. 840 were collected by a Mr Nieuwenhuys, who found the mammals.
Palaeontology
The assemblage of fossils from N 885, Niger, comprises well-mineralized skeletal remains of lacustrine and terrestrial animals stained dark brown to black. The fauna is restricted, but contains a Nile perch (Lates niloticus), a crocodile (Crocodylus cf. niloticus), a chimp-sized hominoid, a medium-sized species of anthra-cothere (Libycosaurus sp.), and a bovid.
Pisces: Lates niloticus (Nile perch) is represented at the Niger site by the posterior basal part of the skull.
Crocodylia: Crocodylus cf. niloticus (Nile crocodile) is represented at locality N 885 by a brevirostrine mandible with buccal depressions between the teeth, two dermal scutes, two articulated vertebrae, the occipital part of a skull and diverse fragments of skull.
Hominoidea is represented at site N 885 by a right mandible fragment (Fig. 1) containing the roots of the first molar. The mandible is slender and moderately deep. The jaw beneath m/1 is 13.2 mm thick, and its depth from the alveolar process to the ventral margin is 31.6 mm. The preserved part of the tooth measured at cervix is 9.4 ×9.7 mm, but judging from the position of the alveoli mesial and distal to the preserved roots, the crown length would originally have been c. 11 mm and the breadth about 9.9 mm. The sublingual fossa is shallow and low down.

This mandible fragment is compatible in size and proportions to medium-sized hominoids with slender mandibular rami. It is too gracile to belong to an australopithecine, even a small individual such as AL 288-1 (Lucy)13 from Hadar, Ethiopia. Comparisons with several hominoids, both extinct and extant, reveal overall similarities to both Pan troglodytes (Fig. 1) and Homo rather than to any other genera. The jaw is slightly less deep and more gracile than that of Orrorin28 and the sublingual fossa is not as deeply excavated.
We consider that the best match for the Niger hominoid fossil is with Pan, but because the fossil is fragmentary, we hesitate to attribute it to this or to any other hominoid genus.
Anthracotheriidae: Libycosaurus is represented at the Niger site by a distal metapodial attributed to a medium-sized species, larger than L. petrocchii and smaller than the large species from Sahabi and Chad.16,23 The section of the bone at the break reveals a marked degree of dorso-plantar compression which distinguishes this fossil from hippopotamid metapodials24 with which it might otherwise be confused.
Bovidae: The family Bovidae is represented at point N 885 by a left frontal with horn core, a distal left radius and a damaged left calcaneum. The horn core belongs to a reduncine antelope, morphologically close to but smaller than Redunca darti.8,10
Age of the Niger fossils
Two specimens from the Niger locality are useful for biochronology. The most telling evidence is the presence of an anthracothere, Libycosaurus sp., equivalent to the medium-sized species from Toros-Ménalla, Chad and Sahabi, Libya.16,23 Secondly, the bovid horn core has some resemblances to, but is smaller than, material from Sahabi15 (Libya), Lukeino,30 Mpesida30 and Lothagam11 (Kenya) and Langebaanweg9 (South Africa). An age of between 11 and 8 Myr is possible for the deposits that yielded these fossils.
Palaeoenvironment and palaeoclimatology
The presence of Nile perch, Lates niloticus, in the Niger deposits indicates the former presence of a freshwater lake or large river in the country. The anthracothere Libycosaurus was likely a denizen of shallow swampy parts of a palaeolake, and the reduncine probably lived close to water. This evidence, combined with the presence of a hominoid primate, indicates that the region was appreciably more humid (at least 750 mm mean annual rainfall) during the Late Miocene than it is today (ranging from sahel to total desert over large areas of the country).
Discussion
Apart from humans, hominoids are today confined to forest and woodland, but the common chimpanzee can survive in sub-humid environments as long as there are enough trees to supply adequate food and security.17 This means that the longest dry period should not exceed two months, and that the area should be well endowed with riparian forest so that there is always a supply of fresh vegetation in the form of fruit, leaves, pith and bark, even during the driest months of the year. In general, however, chimpanzees and gorillas flourish best in tropical rainforest of Guineo-Congolian affinities.31 At present, the most arid environment inhabited by chimpanzees is wooded savanna in Liberia and Miombo woodland in Tanzania.17,32 The presence of a chimp-sized hominoid in the Late Miocene of Niger therefore provides evidence that the country was considerably more humid then than it is today, and was at least covered in wooded savanna if not denser vegetation categories.
The discovery of a Late Miocene aquatic and terrestrial fauna in Niger confirms the view that the Sahara is a relatively young desert, post-dating the Late Miocene. Recent work in Egypt has revealed that central Egypt was covered in woodland to forest some 11–10 Myr ago26,27 and Chad is long known to have been more humid in the past than it is today5,6,20 as were Libya7 and Tunisia.21,22
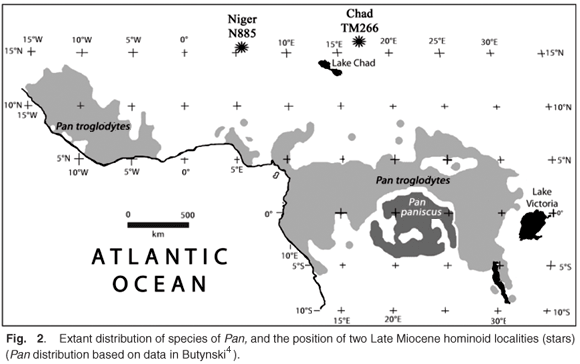
At present, the chimpanzees living closest to the Niger fossil site occur 940 km to the south in Ghana and Nigeria and 1700 km to the west in Mali (Fig. 2).4,32 Niger thus joins an increasing list of African countries that have yielded remains of Late Miocene (c. 11–5.5 Myr ago) hominoids including: Kenya (Lukeino, 6 Myr, two taxa);19,25,28 (Samburu Hills, 9.5 Myr);12 (Nakali, c. 10 Myr);14 Ethiopia (Ch'orora, c. 10.5 Myr, Middle Awash, c. 5.7 Myr)29 and Chad (Toros-Menalla, c. 10–6 Myr)3,23 and it indicates that further research in West Africa will undoubtedly add fuel to the debate about hominid origins. It is clear that during the Late Miocene, hominoids were widespread in Africa and, although the quantity of fossils is limited, the diversity is large, with at least seven lineages already recognized in the time range 11–5.5 Myr ago (in the order in which they were reported: Samburupithecus, Orrorin, Ardipithecus, Sahelanthropus, an unnamed protogorilla-like form, Chororapithecus, and Nakalipithecus) and two between 13 and 11 Myr ago (Otavipithecus and an unnamed chimp-like form). In the circumstances, it seems superfluous to invoke a Late Miocene reintroduction of hominoids into Africa from Eurasia in order to give rise to hominids,1 a view that was attractive only while the Late Miocene hominoid fossil record of Africa was poorly known.
Conclusions
A restricted vertebrate fauna of Late Miocene age from Niger is of interest on account of its age and the presence within it of a hominoid primate and an anthracothere. The discovery helps to fill what was a vast gap in the geographic coverage of fossil hominoids; the nearest known fossil specimen of comparable age being from Chad, over 1000 km to the east and 3500 km from sites in Ethiopia and Kenya that have yielded Late Miocene hominoids. The site is closer to the Spanish Late Miocene hominoid sites (3000 km)18 than it is to the East African ones. The find site, the position of which is not precisely known at present, is about 940 km north of the closest extant hominoids (Pan troglodytes) in Ghana and Nigeria. The fauna as a whole indicates the former presence of a freshwater lake or large river in Niger, and a Late Miocene palaeoclimate considerably more humid than that of today. The scientific value of the Niger hominoid specimen resides in the attention that it will focus on a palaeontologically poorly known part of the continent, which nevertheless probably has a great deal to contribute to our knowledge about the origin of hominids.
We thank Christine Argot and Claire Sagne for their assistance with the collections and registers of fossils at the MNHN. Thanks also to Philippe Richir for making casts and Jacques Treil (MEDES, Toulouse) for CT scans.
1. Begun D. (2001). European hominoids. In The Primate Fossil Record, ed. W.C. Hartwig, pp. 339–368. Cambridge University Press, Cambridge. [ Links ]
2. Bergounioux F. and Crouzel F. (1968). Deux tortues fossiles d'Afrique. Bull. Soc. Hist. Nat. Toulouse 104, 179–186. [ Links ]
3. Brunet M., Guy F. Pilbeam D., Taisso Mackaye H., Likius A., Ahounta D., Beauvilain A., Blondel C., Bocherens H., Boisserie J-R., De Bonis L., Coppens Y., Dejax J., Denys C., Duringer P., Eisenmann V., Fanone G., Fronty P., Geraads D., Lehmann T. Lihoreau F., Louchart A., Mahamat A., Merceron G., Mouchelin G., Otero O., Pelaez Campomanes P., Ponce de Leon M., Rage J-C., Sapanet M., Schuster M., Sudre J., Tassy P., Valentin X., Vignaud P., Viriot L., Zazzo A. and Zollikofer C. (2002). A new hominid from the Upper Miocene of Chad, Central Africa. Nature 418, 145–151. [ Links ]
4. Butynski T.M. (2004). Le chimpanzé commun, Pan troglodytes, taxinomie, distribution, effectif et statut de la conservation. In Chimpanzés d'Afrique de l'Ouest, pp. 15–22. Union Mondial pour la Nature, Gland and Cambridge. [ Links ]
5. Coppens Y. and Koeniguer J-C. (1967). Sur les flores ligneuses disparues plio-quaternaires du Tchad et du Niger. C. R. Acad. Sci. Paris 265, 1282–1285. [ Links ]
6. Coppens Y. and Koeniguer J-C. (1976). Signification climatique des paléoflores ligneuses du Tertiaire et du Quaternaire du Tchad. Bull. Soc. géol. Fr. 18, 1009–1015. [ Links ]
7. Dechamps R. (1987). Xylotomy of fossil wood from the Sahabi Formation. In Neogene Palaeontology and Geology of Sahabi, ed. N. Boaz, pp. 37–41. Alan Liss, New York. [ Links ]
8. Gentry A.W. (1978). The fossil Bovidae of the Baringo Area, Kenya. In Geological Background to Fossil Man, ed. W.W. Bishop, pp. 294–308. Scottish Academic Press, Edinburgh. [ Links ]
9. Gentry A.W. (1980). Fossil Bovidae (Mammalia) from Langebaanweg, South Africa. Ann. S. Afr. Mus. 79, 213–337. [ Links ]
10. Gentry A.W. and Gentry A. (1978). Fossil Bovidae (Mammalia) of Olduvai Gorge, Tanzania. Bull. Br. Mus. (Nat. Hist.) Geol. 29, 289–446 and 30, 1–83. [ Links ]
11. Harris J. (2003) Bovidae from the Lothagam Succession. In Lothagam: The Dawn of Humanity in Eastern Africa, eds M. Leakey and J. Harris, pp. 531-579. Columbia University Press, New York. [ Links ]
12. Ishida H. and Pickford M. (1997). A new late Miocene hominoid from Kenya: Samburupithecus kiptalami gen. et sp. nov. C.R. Acad. Sci. Paris 325, 823–829. [ Links ]
13. Johanson D.C., White T.D. and Coppens Y. (1978). A new species of the genus Australopithecus (Primates, Hominidae) from the Pliocene of East Africa. Kirtlandia 28, 1–14. [ Links ]
14. Kunimatsu Y., Nakatsukasa M., Sawada Y., Sakai T., Hyodo M., Hyodo H., Itaya T., Nakaya H., Saegusa H., Mazurier A., Saneyoshi M., Tsujikawa H., Yamamoto A. and Mbua E. (2007). A new Late Miocene great ape from Kenya and its implications for the origins of African great apes and humans. Proc. Natl Acad. Sci. USA 104, 19220– 19225. [ Links ]
15. Lehmann U. and Thomas H. (1987). Fossil Bovidae (Mammalia) from the Mio-Pliocene of Sahabi, Libya. In Neogene Paleontology and Geology of Sahabi, ed. N. Boaz et al., pp. 323–335. Alan Liss, New York. [ Links ]
16. Lihoreau F. (2003). Systématique et Paléoécologie des Anthracotheriidae (Artiodactyla: Suiformes) du Mio-Pliocène de l'Ancien Monde: Implications paléobiogéographiques. Thèse, University of Poitiers, France. [ Links ]
17. Moore J. (1992). 'Savanna' Chimpanzees. In Topics in Primatology, 1: Human Origins, eds T. Nishida, W. McGrew, P. Marler, M. Pickford and F. de Waal, pp. 99–118. University of Tokyo Press, Tokyo. [ Links ]
18. Moyà-Solà S. and Köhler M. (1996). A Dryopithecus skeleton and the origins of great ape locomotion. Nature 379, 156–159. [ Links ]
19. Nakatsukasa M., Pickford M., Egi N. and Senut B. (2007). Body weight, femoral length and stature of Orrorin tugenensis, a 6 Ma hominid from Kenya. Primates 48, 171–178. [ Links ]
20. Otero O., Pinton A., Vignaud P. and Brunet M. (2006). The fish fauna from TM 266 (Late Miocene from Toros-Menalla, Western Djurab, Chad), environment and biogeography. Annual Symposium of Vertebrate Palaeontology and Comparative Anatomy, Paris, Abstracts, p. 10. [ Links ]
21. Pickford M. (1999). Aubréville's hypothesis of a southwards shift of Africa's vegetation belts since the Miocene. In Wood to Survive, Liber Amicorum Roger Dechamps, eds F. Maes and H. Beeckman. Ann. Sci. Econ. Mus. Roy. Afr. Cent. Tervuren 25, 195–212. [ Links ]
22. Pickford M. (2000). Crocodiles from the Beglia Formation, Middle/Late Miocene Boundary, Tunisia, and their significance for Saharan palaeoclimatology. Ann. Paléont. 86, 59–67. [ Links ]
23. Pickford M. (2008). Libycosaurus petrocchii Bonarelli, 1947, and Libycosaurus anisae (Black, 1972) (Anthracotheriidae, Mammalia): nomenclatural and geochronological implications. Ann. Paléont. 94, 39–55. [ Links ]
24. Pickford M. (2008). The myth of the hippo-like anthracothere: The eternal problem of homology and convergence. Revista Española de Paleontologia 23, 31–90. [ Links ]
25. Pickford M. and Senut B. (2005). Hominoid teeth with chimpanzee- and gorilla-like features from the Miocene of Kenya: implications for the chronology of the ape-human divergence and biogeography of Miocene hominoids. Anthropol. Sci. 113, 95–102. [ Links ]
26. Pickford M., Wanas H., Mein P. and Soliman H. (2006). Indications for a humid climate in the Western Desert of Egypt 11–10 million years ago: evidence from Galagidae (Primates, Mammalia). C. R. Palévol 5, 935–943. [ Links ]
27. Pickford M., Wanas H., Mein P. and Soliman H. (2008). Humid conditions in the Western Desert of Egypt during the Vallesian (Late Miocene). Proc. Third International Conference on the Geology of the Tethys, South Valley University, Aswan, January, 2008, pp. 1–17. [ Links ]
28. Senut B., Pickford M., Gommery D., Mein P., Cheboi K. and Coppens Y. (2001). First hominid from the Miocene (Lukeino Formation, Kenya). C. R. Acad. Sci. Paris 332, 137–144. [ Links ]
29. Suwa G., Kono R.T., Katoh S., Asfaw B. and Beyene Y. (2007). A new species of great ape from the late Miocene epoch in Ethiopia. Nature 448, 921–924. [ Links ]
30. Thomas H. (1980). Les Bovidés du Miocène supérieur des couches de Mpesida et de la formation de Lukeino (District de Baringo, Kenya). Actes 8ème Congr. Panafr. Préhist. Etudes Quat., Nairobi, pp. 82–91. [ Links ]
31. White F. (1986). La végétation de l'Afrique. Mémoir accompagnant la carte de végétation de l'Afrique, Unesco/AETFAT/UNSO. Orstom–Unesco, Paris. [ Links ]
32. Wolfheim J.H. (1983). Primates of the World. Distribution, Abundance and Conservation. New York Zoological Society, University of Washington Press, Seattle and London. [ Links ]
* Author for correspondence. E-mail: pickford@mnhn.fr

