










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.104 no.3-4 Pretoria mar./abr. 2008
RESEARCH LETTERS
A partial skull of Paranthropus robustus from Cooper's Cave, South Africa
Christine Steininger; Lee R. Berger; Brian F. Kuhn
Institute for Human Evolution and the Bernard Price Institute for Palaeontological Research, School of Geosciences, University of the Witwatersrand, Private Bag 3, WITS 2050, South Africa
ABSTRACT
A partial hominin skull (COB 101) was identified in the fossil collections of the Transvaal Museum, Pretoria, attributed to the Cooper's Cave site in South Africa. The find represents the most complete hominin specimen recovered from localities at this site to date. COB 101 comprises the supraorbital, zygomatic, infraorbital and nasoalveolar regions of the right side, and the right upper third premolar. The specimen has undergone post-depositional distortion that resulted in the flattening of the facial structures. Here we describe and compare COB 101 with other hominin material from Africa and find that this specimen shares numerous diagnostic features with Paranthropus robustus. The discovery of COB 101 augments the number of specimens attributed to this species from other South African sites and other Cooper's Cave localities.
Introduction
A partial skull, COB 101, was identified in the collections of the Transvaal Museum, Pretoria.1 The fossil was found in the collections that were attributed by museum records to Cooper's Cave. Two other ex situ hominin specimens have previously been described from early collecting activities—an upper third molar attributed to Australopithecus africanus2,3 and an upper central incisor attributed to Homo sp. indet.4 In situ hominin specimens have been recovered from the Cooper's D locality by more recent excavation activities and these are currently being studied and assessed.5
The Cooper's Cave site is hosted in dolomites of the Monte Cristo Formation (Malmani Subgroup, Transvaal Supergroup). The site is situated approximately 45 km northwest of Johannesburg, half-way between Sterkfontein to the west and Kromdraai to the east. Given our understanding based on recent excavations, Cooper's Cave comprises three fossil-rich areas, provisionally recognized as Cooper's A, B, and D localities, with most of the fauna having been recovered from Cooper's D. Based on biostratigraphy, the age of the excavated deposits of Cooper's D is estimated at between 1.6 and 1.9 million years ago (Mya).5,6 This is penecontemporaneous with Sterkfontein Members 4 and 5,7–10 Swartkrans Members 1, 2 and 3,11–13 Kromdraai A and B,12,14 and Drimolen.15,16 Radiometric dating methods are currently being applied to the various Cooper's Cave sites to try to obtain more precise dates. Initial radiometric results are, however, comparable to the biostratigraphic age-estimations (R. Pickering, pers. comm.)
The purpose of this paper is to provide a description and analysis of the hominin specimen, COB 101, which evinces morphology not previously known for the Cooper's Cave hominin assemblage.
Materials and methods
COB 101 was compared with original hominin specimens of A. africanus (n = 17), Paranthropus robustus (n = 15), Homo habilis (n = 7) and H. ergaster (n = 1), and high-resolution casts of A. afarensis (n = 3), P. aethiopicus (n = 1) and P. boisei (n = 6) (Table 1). Observations were aided by stereomicroscopy. All dimensions were chord distances taken with digital callipers to the nearest 0.1 mm. Because of the nature of the preservation of COB 101, measurements of breadth on all taxa were taken from the midsagittal plane. Non-metric facial parameters follow Lockwood17 and Rak.18 Facial measurements follow Martin and Knuβmann19 and Lockwood.17 Dental diameters were measured following Wood.20

Description
Only the right portion of the upper face, including an upper right third premolar, is preserved in COB 101 (Fig. 1). While the specimen is distorted and has post-depositional damage. The supraorbital area, the right side of the glabella and torus are preserved. The supraorbital and zygomatic regions are slightly separated but held together by adhering breccia. The zygomatic has been displaced from its anatomical position and is compressed against the maxilla. The inferior part of the infraorbital and the nasoalveolar regions are the best-preserved areas of the specimen, with the only damage being slight weathering. Age-related features are not well preserved, but based on moderate wear of the permanent premolar, the individual is almost certainly an adult, comparable in dental wear stage to P. robustus specimens SK 49 and SK 57 from Swartkrans.

The temporal line of COB 101 is placed high and is well marked on the frontal squama, indicating the probable presence of a sagittal crest. Recent studies on sexual dimorphism on P. robustus suggest that a prominent sagittal crest reaching onto the frontal bone is consistent with larger males of this species.21 Accordingly, even though this specimen is damaged it is possible to state that it is probably a male. The supraorbital torus is continuous, similar in morphology to Clarke's 'supraorbital rib' attributed to paranthropines22 On the torus, just lateral to the glabella, a small crescent-shaped supraorbital notch changes the relief of the torus slightly. Overall, the torus is thin, 6 mm at its thickest vertical height and vermiculated throughout. The superciliary eminence is absent and this trait is consistent with other specimens attributed to P. robustus. Anteriorly, the preserved portion of the glabella is flat and there seems to be variation of the glabella prominence among most species included in this study, except for P. aethiopicus and P. boisei, where the glabella is consistently marked (Table 2).
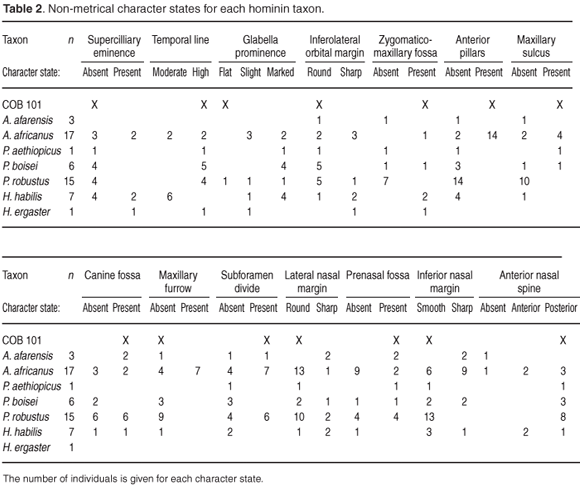
The zygomaticomaxillary fossa of COB 101 is deep. This feature has been noted as dominant in paranthropines and A. afarensis.18 The inferolateral orbital margin is rounded, and is similar in appearance to that of A. africanus and P. robustus. The zygomatic prominence is slight, located inferiorly on the specimen, close to the masseteric origin (Table 2).
On the maxilla, the marginally distorted anterior pillar slightly overhangs the nasal aperture. The widest portion of the preserved anterior pillar is 10.6 mm. The anterior pillar is found in most A. africanus specimens and all P. robustus specimens. A marked canine–premolar eminence forms the inferior part of the anterior pillar. The maxillary sulcus is preserved immediately below the infraorbital foramen. A large canine fossa, at the same level as the inferior nasal margin, faces laterally. A maxillary furrow, which is a variable though prominent feature of most A. africanus specimens, is absent in COB 101. The subforamen divide is present in COB 101 as a thin marked ridge, starting just above the canine alveolar margin that diverts laterally slightly below the infraorbital foramen,thereby framing the maxillary sulcus. The combination of the anterior pillars and the subforamen divide, and the lack of a maxillary furrow, is consistent with most P. robustus specimens (Table 2).
In the nasoalveolar region, the lateral nasal margin is rounded with a deep prenasal fossa24 behind it. The depth of the fossa may be exaggerated by the distortion of the anterior pillars. The nasal spine is posterior from the inferior nasal margin. The nasoalveolar gutter is present, sloping smoothly up into the nasal aperture with no distinct nasal sill present. The slight alveolar prognathism observed is exaggerated by the incisor eminences.
Though we were able to obtain some measurements in the nasoalveolar region of the face, these measurements provide only estimates due to the crushed and distorted nature of COB 101. The measurements are as follows: alveolar height is c. 23.2 mm, nasal breadth is c. 13.1 mm, and snout breadth is c. 22.7 mm.
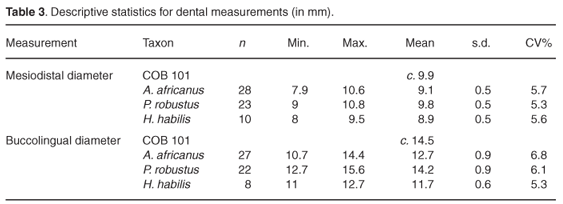
The third premolar crown is almost complete, but is crushed mesiodistally and buccally. The only aspect of the tooth that is not damaged is the lingual face and a small part of the mesiolingual face. Part of the occlusal surface of the protocone is missing and the occlusal surface of the paracone is partially crushed. The occlusal surface and the mesial face are stained by manganese. The tooth is moderately worn, with small patches of dentine visible in the area of the protocone. This surface is worn flat and the occlusal edges of the tooth are rounded. Although the mesial face of the tooth is distorted and slightly crushed, interstitial wear from the canine is still evident and the crown has retained its oval shape. The buccal face is moderately convex occlusocervically, and more so mesiodistally, giving the tooth a bulbous appearance. Buccal grooves are not present. Perikymata are visible on the mesiobuccal aspect of the tooth. The lingual face is similar to the buccal face in its convexity, but is much wider along the cervical margin. Because of damage, the mesiodistal diameter was measured across the least crushed part of the tooth, across the protocone just below the central fissure. The estimated crown diameters are: mesiodistal c. 9.9 mm, buccolingual c.14.5 mm, and preserved crown height c. 6.7 mm (Table 3).
Discussion
The combination of features in the supraorbital and the nasoalveolar region in COB 101 are similar to these morphological areas in P. robustus (see Table 2). These features tend to be related to robusticity and degree of development, and it is important to note that the morphology of the nasoalveolar region is often used to separate paranthropines from australopithecines.19,24 The presence of an anterior pillar and the lack of a maxillary furrow are considered diagnostic features found in most P. robustus specimens.18 In this regard COB 101 most closely resembles two P. robustus specimens from Swartkrans (SK 48 and SK 79).
Morphologically, the molarized upper third premolar of COB 101 is similar to specimens attributed to P. robustus and P. boisei. Notable features are the oval crown, large bulbous appearance of the tooth, and the flat wear on the occlusal surface. The mesiodistal diameter of COB 101 is well within the specimens attributed to P. robustus (9.8 ± 0.5 mm, n = 23) and A. africanus (9.1 ± 0.5 mm, n = 28) where the buccolingual diameter falls within only the P. robustus range (14.2 ± 0.9 mm). The size of the upper premolar of COB 101 indicates affinities with P. robustus (Fig. 2). We hesitate, however, to assume that these dimensions provide evidence for placement within P. robustus. There are two reasons for caution: (1) the damaged state of the upper third premolar means that our measurements are estimates, and (2) mesiodistal and buccolingual diameters of upper third premolars are recognized as being highly variable among hominin species, with a particularly high variation in A. africanus and P. robustus24–27 (Fig. 2).

In summary, most of the morphological evidence in the supraorbital and the nasoalveolar regions of COB 101 is consistent with other specimens attributed to P. robustus, particularly SK 48 and SK 79. In addition, the morphology and wear of the upper third premolar provides further evidence for associating the specimen with the genus Paranthropus. The evidence reviewed above suggests that COB 101 is best attributed to P. robustus.
As COB 101 is of unknown provenance, its precise association with other Paranthropus material recovered from Cooper's D, or the Cooper's specimens attributed to A. africanus and Homo, remains unclear. Owing to the possibility of a temporal overlap between latest A. africanus (c. 1.5–2.1 Mya),7–9 and earliest P. robustus (c. 1.9 Mya),11,13 and considering the early dates proposed for some of the deposits at Cooper's,5 the presence of all three species at this site is plausible.
We thank two reviewers for their helpful comments. Thanks also go to Francis Thackeray, Stephany Potze and Heidi Fourie of the Transvaal Museum, Pretoria, for their assistance with the hominin material in their care. We wish also to thank Andre Keyser and Colin Menter for allowing us brief access to a Drimolen specimen, DNH 7. This research has been funded by the Palaeontological Scientific Trust (PAST) and the University of the Witwatersrand.
1. Steininger C.M. (2003). Taxonomic affinity of a new specimen from Coopers Cave, South Africa. M.Sc. thesis, University of the Witwatersrand, Johannesburg. [ Links ]
2. Broom R. and Schepers G.W. (1946). The South African fossil ape-man: the Australopithecinae. Transvaal Museum Memoir 2, Pretoria [ Links ]
3. Robinson J.T. (1956). The dentition of the Australopithecinae. Transvaal Museum Memoir 9, Pretoria. [ Links ]
4. Berger L.R., Pickford M. and Thackeray F. (1995). A Plio-Pleistocene hominid upper central incisor from the Cooper's site, South Africa. S. Afr. J. Sci. 91, 541–542. [ Links ]
5. Berger L.R., de Ruiter D.J., Steininger C.M. and Hancox P.J. (2003). Preliminary results of excavations at the newly discovered Coopers D deposit, Gauteng, South Africa. S. Afr. J. Sci. 99, 276–278. [ Links ]
6. Delson E. (1988). Chronology of South African australopith site units. In Evolutionary History of the 'Robust' Australopithecines, ed. F. Grine, pp. 317–324. Aldine de Gruyter, New York. [ Links ]
7. Berger L.R., Lacruz R.S. and de Ruiter D.J. (2002). Revised age estimates of Australopithecus-bearing deposits at Sterkfontein, South Africa. Am. J. Phys. Anthrop. 119, 192–197. [ Links ]
8. Thackeray J.F. (2006). Reconceptualisation of African hominins dated between 1.5 and 2.5 mya. Ann. Transv. Mus. 43, 116–117. [ Links ]
9. Thackeray J.F. and Dupont L.M. (2006). Last recorded occurrence of the Australopithecus africanus and climatic change in southern Africa. S. Afr. J. Sci. 102, 50. [ Links ]
10. Partridge T.C. (2006). Dating of Member 4 deposits at Sterkfontein. Trans. R. Soc. S. Afr. 60(2), 107–110. [ Links ]
11. Brain C.K. (1993). Structure and stratigraphy of the Swartkrans Cave in light of the new excavations. In Swartkrans: A Cave's Chronicle of Early Man, ed. C.K. Brain, pp. 23–33. Transvaal Museum Monograph 8, Pretoria. [ Links ]
12. Vrba E.S. (1982). Biostratigraphy and chronology, based particularly on Bovidae, of southern hominid-associated assemblages: Makapansgat, Sterkfontein, Taung, Kromdraai, Swartkrans; also Elandsfontein (Saldanha), Broken Hill (now Kabwe) and Cave of Hearths. In Home erectus et la place de l'homme de tautavel parmi les hominidés fossiles, ed. M.A. de Lumley, Première Congrès International de Paléontologie Humaine, UNESCO Colloque International du CNRS, prétirages, pp. 707–752. [ Links ]
13. Albarede F., Balter V., Brage J., Blichert-Toft J., Telouk P. and Thackeray F. (2006). U-Pb dating of enamel from the Swartkrans Cave hominid site (South Africa) by MCICP-MS. Geochim. Cosmochim. Acta 70, A7. [ Links ]
14. Thackeray J.F., Kirschvink J.L. and Raub T.D. (2002). Palaeomagnetic analyses of calcified deposits from the Plio-Pleistocene hominid site of Kromdraai, South Africa. S. Afr. J. Sci. 98, 537–540. [ Links ]
15. Keyser A.W. (2000). The Drimolen skull: the most complete australopithecine cranium and mandible to date. S. Afr. J. Sci. 96, 189–193. [ Links ]
16. Keyser A.W., Menter C.G., Moggi-Cecchi J., Pickering T.R. and Berger L.R. (2000). Drimolen: a new hominid-bearing site in Gauteng, South Africa. S. Afr. J. Sci. 96, 193–197. [ Links ]
17. Lockwood C.A. (1997). Variation in the face of Australopithecus africanus and other African hominoids. Ph.D. dissertation, University of the Witwatersrand, Johannesburg. [ Links ]
18. Rak Y. (1983). The Australopithecine Face. Academic Press, New York. [ Links ]
19. Martin R. and Knuβmann R. (1988). Anthropologie: Handbuch der Vergleichenden Biologie des Menschen. Gustav Fisher-Verlag, Stuttgart. [ Links ]
20. Wood B.A. 1991. Koobi Fora Research Project IV: Hominid Cranial Remains From Koobi Fora. Clarendon Press, Oxford. [ Links ]
21. Lockwood C.A., Menter C.G., Moggi-Cecchi J. and Keyser A.W. (2007). Extended male growth in fossil hominin species. Science 319, 1443–1446. [ Links ]
22. Clarke R.J. (1977). The cranium of the Swartkrans hominid, SK 847 and its relevance to human origins. Ph.D. dissertation, University of the Witwatersrand, Johannesburg. [ Links ]
23. McCollum M.A., Grine F.E., Ward S.C. and Kimbel W.H. (1993). Subnasal morphological variation in extant hominoids and fossil hominids. J. Hum. Evol. 24, 87–111. [ Links ]
24. Grine F.E. and Strait D.S. (1994). New hominid fossils from Member 1 'Hanging Remnant', Swartkrans Formation, South Africa. J. Hum. Evol. 26, 57–75. [ Links ]
25. Moggi-Cecchi J., Tobias P.V. and Beynon A.D. (1998). The mixed dentition and associated skull fragments of a juvenile fossil hominin from Sterkfontein, South Africa. Am. J. Phys. Anthrop. 106, 425–465. [ Links ]
26. Braga J. and Thackeray J.F. (2003). Early Homo at Kromdraai B: probabilistic and morphological analysis of the lower dentition. C. R. Palevol. 2, 269–279. [ Links ]
27. Moggi-Cecchi J., Grine F.E. and Phillip T.V. (2006). Early hominid dental remains from Members 4 and 5 of the Sterkfontein Formation (1966–1996 excavations): Catalogue, individual associations, morphological descriptions and initial metrical analysis. J. Hum. Evol. 50, 239–328. [ Links ]
Received 12 October 2007. Accepted 2 March 2008.
Author for correspondence. E-mail: christine.steininger@student.wits.ac.za

