










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Science
versión On-line ISSN 1996-7489
versión impresa ISSN 0038-2353
S. Afr. j. sci. vol.104 no.1-2 Pretoria ene./feb. 2008
RESEARCH LETTERS
Antiplasmodial activities of some abietane diterpenes from the leaves of five Plectranthus species
R.L. van ZylI; F. KhanII; T.J. EdwardsIII; S.E. DrewesIII, *
IPharmacology Division, Department of Pharmacy and Pharmacology, Faculty of Health Sciences, University of the Witwatersrand, 7 York Rd, Parktown 2173, South Africa
IISchool of Chemisty, University of KwaZulu-Natal, Pietermaritzburg 3200, South Africa
IIISchool of Biological and Conservation Sciences, University of KwaZulu-Natal, Pietermaritzburg 3200, South Africa
ABSTRACT
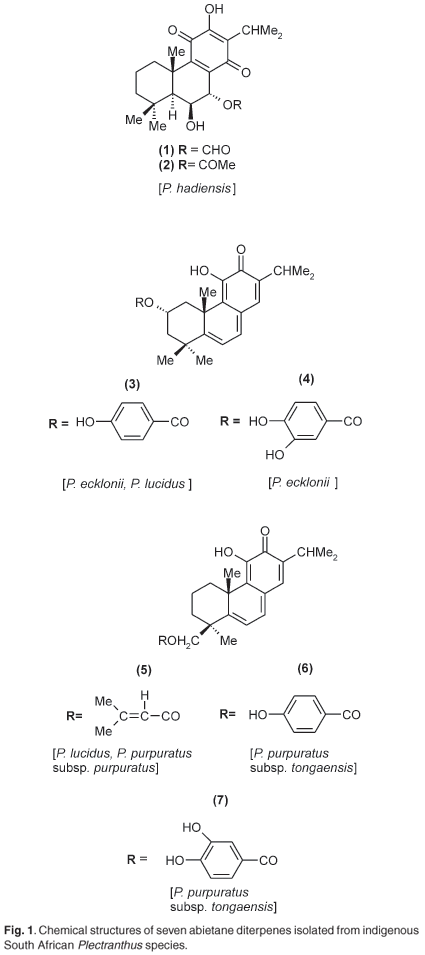
Seven known abietane diterpenes were isolated from five Plectranthus species, namely P. hadiensis, P. lucidus, P. ecklonii, P. purpuratus subsp. purpuratus and P. purpuratus subsp. tongaensis. The seven compounds were tested for their antiplasmodial activity and for their ability to inhibit β-haematin formation. Overall, they showed good activity (IC50 values ranging from 3.11 to 14.65 µM), with one compound being 62% as effective as chloroquine in inhibiting β-haematin formation. However, the cytotoxicity profile indicated a low degree of specificity towards the malaria parasite. When combined with quinine, three compounds interacted in an additive manner whereas one interacted synergistically.
Introduction
It is estimated from recent studies that at least 300 million cases of malaria occur worldwide,1,2 resulting in millions of deaths every year. Our motivation to investigate the genus Plectranthus (Lamiaceae) as a possible source of antiplasmodial compounds has been influenced by three interrelated observations published in recent literature:
(a) Of the known species in the genus (350 in Africa, of which 53 occur in South Africa and Namibia), many have been tested worldwide for a variety of purposes. For example, they are effective against many harmful bacteria,3 have an anti-proliferation effect on human lymphocytes,4 can control insect feeding habits,5 and are good insect repellents.6
(b) In African traditional medicine, 45 species of Plectranthus have received attention, amongst others, for treatment of gastro-intestinal disorders,7 as anti-microbial agents,8 for the treatment of wounds,9 the alleviation of respiratory conditions10 and for malaria.9,11-14
(c) In a recent review,15 the statement is made that several of the compounds from Plectranthus, that have high anti-microbial activities, also possessed good antiplasmodial properties.
These three observations, coupled with recent findings16 that certain species of Plectranthus, notably P. hadiensis, have extremely high anti-fungal and anti-bacterial activity, have been the driving force for the present investigation.
Experimental
See Appendix.
Results and discussion
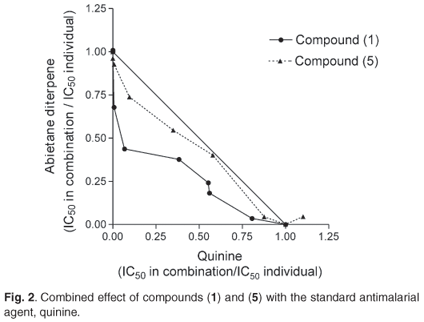
The antiplasmodial activities against a chloroquine-resistant strain of Plasmodium falciparum and the inhibitory effects of the seven abietane diterpenes (1)-(7) (Fig. 1) on β-haematin formation as well as their toxic effects are shown in Table 1. Relative to chloroquine and quinine, the seven compounds were less active, but showed significant activity in inhibiting the formation of β-haematin. These compounds (1), (4) and (7) were more active than quinine. As can be seen from the isobolograms (Fig. 2), when combined with quinine, compounds (4), (5) and (7) interacted in an additive manner, whereas compound (1) interacted synergistically. To ensure clarity in Fig. 2, the results obtained from (4) and (7) were not included as they overlapped the results obtained for (5).
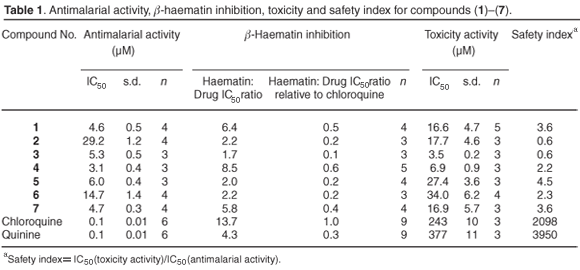
While the activities of our abietane diterpenes (Table 1) may not be as high as previously reported plant metabolites, they display potential pharmacological activities worthy of further investigation. Until this study, the only reported anti-protozoal activity of abietane diterpenes is that of 7-hydroxy-12-methoxy-20-nor-abieta-1,5(10),7,9,12-pentaen-6,14-dione and abieta-8,12-dien-11,14-dione against the amastigote form of Leishmania donovani and L. major.17 The IC50 values for the latter two abietane diterpenes of 170- and 120-nM against L. donovani amastigotes are regarded as 'appreciable' by the authors. When compared to the control drug, sodium stibogluconate (IC50: 10.6 nM) there is a 15-fold decrease in activity with the abietane diterpenes. By comparison, our three best IC50 results [from (4), (1) and (7)] against Plasmodium falciparum were 3.11, 4.59, and 4.74 µM, respectively, and when compared to chloroquine were approximately 35-fold less active (Table 1). The weakest result was for (2) with an IC50 of 29.20 µM.
The lipophilic nature of our abietane diterpenes (1)-(7) enables easy transport across the erythrocytic and parasitic membranes to accumulate in the parasitic food vacuole. The inhibitory effect of our compounds appears to relate to their ability to prevent the formation of β-haematin. This is an important effect, because the malaria parasite degrades haemoglobin and the released haem, which is toxic to the parasite, is normally converted to the inert malaria pigment, β-haematin.18 As can be seen in Table 1, compounds (1), (4) and (7) are more effective than quinine, and compound (4) is 62% as active as chloroquine.
Most of the diterpenes are known to combine high antiprotozoal activity with high toxicity for mammalian cells (such as kidney epithelial cells) (Table 1), hepatoma cells, as well as colonic carcinoma cells.19 The unfavourable safety indices (Table 1) imply that our compounds are poor candidates for development as antimalarial agents. However, the selective antiprotozoal action that inhibits β-haematin formation differs enough from its toxic mode of action (r2 = 0.097) to an extent that appropriate formulations and analogues of compound (4) could retain or enhance antimalarial activity, while reducing toxicity. This particular approach has been successfully applied with the improved potency of the 2,7-dibromo analogue of crytolepine, along with decreased in vivo toxicity.20
Conclusions
These particular findings, coupled with the general statement by the Kew group15 that too little is known about the chemistry of other (excluding forskolin) species of Plectranthus to explain their traditional uses, emphasizes that the results presented here are only a start of a research direction. This involves the approach of using natural products as model compounds for chemical modification,20 ideally increasing anti-parasitic activity, while decreasing toxicity.
A concern is the possibility of interactions between traditional medicines and commonly used antimalarial agents, particularly because these interactions are thinly documented. It is possible that patients taking traditional medicines for the treatment or prevention of malaria could be hospitalized and also administered conventional antimalarial agents such as quinine. Therapeutic failure could be possible if the two treatments interact antagonistically, as has been observed between Aspilia africana and artemisinin.30 However, the interactions observed between quinine and the diterpenoids (4), (7) and (5) are found to be additive; whilst a synergistic interaction has been observed between quinine and compound (1). This latter interaction is of therapeutic importance due to the rapid development of drug resistance, and the necessity to administer two or more agents concurrently, in order to achieve a desired therapeutic goal.
Thanks are due to the University of KwaZulu-Natal Research Committee for the award of a postdoctoral bursary (F.K.) and the provision of financial assistance; to C.-T. Chen for technical assistance; and the University of the Witwatersrand Faculty Research Committee/NRF for financial support.
1. Snow R.W., Guerra C.A., Noor A.M., Myint H.J. and Hay S.I. (2005). The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature 434, 214-217. [ Links ]
2. Willcox M.L. and Bodeker G. (2000). Plant-based malaria control: research initiative on traditional antimalarial methods. Parasitol. Today 16, 220-221. [ Links ]
3. Texeira A.P., Batista O., Simões M.F., Nasimento J., Duarte A., De la Torre M.C. and Rodrigue B. (1997). Phytochemistry 44, 325-327. [ Links ]
4. Cerqueira F., Cordeiro-Da-Silva A., Gaspar-Marques C., Simões F., Pinto M.M. and Nascimento M.S.J. (2004). Effect of abietane diterpenes from Plectranthus grandidentatus on T- and B-lymphocyte proliferation. Bioorg. Med. Chem. 12, 217-223. [ Links ]
5. Wellsow J., Grayer R.J., Veitch N.C., Kokuban T., Lelli R., Kite G.C. and Simmonds M.S.J. (2006). Insect-antifeedant and antibacterial activity of diterpenoids from species of Plectranthus. Phytochemistry 67, 1818-1825. [ Links ]
6. Omolo M.O., Okinyo D., Ndiege I.O., Lwande W. and Hassanali A. (2004). Repellency of essential oils of some Ken yan plants against Anopheles gambiae. Phytochemistry 65, 2797-2802. [ Links ]
7. Kokwaro J.O. (1993). Medicinal Plants of East Africa. Kenya Literature Bureau, Nairobi. [ Links ]
8. Matu E.M. and Van Staden J.H. (2003). Anti-bacterial and anti-inflammatory activities of some plants used for medicinal purposes in Kenya. J. Ethnopharmacol. 87, 35-41. [ Links ]
9. Githinji C.W. and Kokwaro J.O. (1993). Ethnomedicinal study of major species in the family Labiatae from Kenya. J. Ethnopharmacol. 39, 197-203. [ Links ]
10. Boily Y. and Van Puyvelde L.J. (1986). Screening of medicinal plants of Rwanda (Central Africa) for anti-microbial activity. J. Ethnopharmacol. 16, 1-13. [ Links ]
11. Hutchings A., Scott A.H., Lewis G. and Cunningham A. (1996). Zulu Medicinal Plants - An Inventory. University of Natal Press, Pietermaritzburg. [ Links ]
12. Neuwinger H.D. (2000). African Traditional Medicine. A Dictionary of Plant Use and Applications. Medpharma Scientific Publications, Stuttgart. [ Links ]
13. Rwangabo P.C. (1993). La médicine traditionnelle au Rwanda. Editions Karthala, ACCT. [ Links ]
14. Schlage C.H., Mabula C., Mahunnah R.L.A. and Heinrich M. (2000). Medicinal plants of Washamba (Tanzania): documentation and ethnopharmacological evaluation. Plant Biol. 2, 83-92. [ Links ]
15. Lukhoba C.W., Simmonds M.S.J. and Paton A.J. (2006). Plectranthus: a review of ethnobotanical uses. J. Ethnopharmacol. 103, 1-24. [ Links ]
16. Gurlal P. (2005). Isolation and characterization of biologically-active chemicals in selected indigenous plants. M.Sc. thesis, University of KwaZulu-Natal, Pietermaritzburg. [ Links ]
17. Tan N., Kaloga M., Radtke O.A., Kiderlen A.F., Öksüz S., Ulubelen A. and Kolodziej H. (2002). Abietane diterpenoids and triterpenoic acids from Salvia cilicica and their antileishmanial activities. Phytochemistry 61, 881-884. [ Links ]
19. Egan T.J., Ross D.C. and Adams P.A. (1994). Quinoline anti-malarial drugs inhibit spontaneous formation of β-haematin (malaria pigment). FEBS Lett. 352, 54-57. [ Links ]
19. Marques C.G., Pedro M., Simões M.F.A., Nascimento M.S.J., Pinto, M.M.M. and Rodríquez B. (2002). Effect of abietane diterpenes from Plectranthus grandidentatus on the growth of human cancer cell lines. Planta Med. 68, 839-840. [ Links ]
20. Wiesner J., Ortmann R., Jomaa H. and Schlitzer M. (2003). New antimalarial drugs. Angew. Chem. Int. Edit. 42, 5274-5293. [ Links ]
21. Miyase T., Rüedi P. and Eugster C.H. (1977). Diterpenoide Drüsenfarbstoffe aus Labiaten. Helv. Chim. Acta 60, 2770-2779. [ Links ]
22. Hensch M., Rüedi P. and Eugster C.H. (1975). Horminon, Taxochinon und weitere Royleanone aus zwei abessinischen Plectranthus-species. Helv. Chim. Acta 58, 1921-1934. [ Links ]
23. Rüedi P. and Eugster C.H. (1978). Diterpenoide Drüsenfarbstoffe aus Labiaten: Sechs neue p-Chinomethane aus Plectranthus parviflorus Willd. Helv. Chim. Acta 61, 709-715. [ Links ]
24. Katti S.B.and Eugster C.H. (1982). Diterpenoide Chinomethane, vinyloge Chinone und ein Phyllocladen-Derivat aus Plectranthus purpuratus. Helv. Helv. Chim. Acta 65, 2189-2197. [ Links ]
25. Jansen J.B. and Trager W. (1976). Human malaria parasites in continuous culture. Science 193, 674-675. [ Links ]
26. Desjardins R.E., Carfield C.J., Haynes D.J. and Chulay J.D. (1972). Quantitative assessment of antimalarial activity in a semi-automated microdilution technique. Antimicrob. Agents Ch. 16, 710-718. [ Links ]
27. Berenbaum M.C. (1989). A method for testing synergy with any number of agents. J. Infect. Dis. 137, 122-130. [ Links ]
28. Chemaly S.M., Chen C.-T. and Van Zyl R.L. (2007). Naturally occurring cobalamines have antimalarial activity. J. Inorg. Biochem. 101, 764-773. [ Links ]
29. Mosmann T. (1983). Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxic assays. J. Immunolog. Methods 65, 55-63. [ Links ]
30. Waako P.J., Smith P. and Folb P.I. (2005). In vitro interactions of Aspilia africana (Pers.) C.D. Adams, a traditional antimalarial medicinal plant, with artemisinin against Plasmodium falciparum. J. Ethnopharmacol. 102, 262-268. [ Links ]
Received 23 November 2007. Accepted 19 February 2008.
* Author for correspondence. E-mail: drewes@ukzn.ac.za
Experimental
General experimental procedures. The NMR spectra were recorded in CDCl3 on a Varian Gemini spectrometer operating at 500 MHz for 1H and 125 MHz for 13C, respectively. Silica gel was used for flash column chromatography. TLCs were run on Merck Si60 F254 plates and visualized by heating after dipping with anisaldehyde solution in ethanol/H2SO4.
Chemicals and biochemicals. Chloroquine diphosphate, quinine hydrochloride and haemin were from Sigma-Aldrich (South Africa); 3H-hypoxanthine from Amersham Biosciences (South Africa) and 3-[4,5-dimethyl-2-thiazol-yl]-2,5-diphenyl-2H-tetrazolium bromide (MTT) from USB (AEC Amersham, South Africa). Quinine and chloroquine were used as the reference antimalarial agents.
Plant material. Plants were collected from the University Botany Garden on the Life Sciences Campus of the University of KwaZulu-Natal. Plant identification was done by a co-author (T.J.E.). Voucher specimens were deposited at the Bews Herbarium of the university under the name of T.J. Edwards. The five species of Plectranthus used in this study are decorative plants, and are cultivated in many household gardens in South Africa.
Extraction and isolation. Air-died leaves from Plectranthus hadiensis, P. lucidus, P. ecklonii, P. purpuratus subsp. purpuratus, P. purpuratus subsp. tongaensis were powdered and extracted with CH2Cl2 and separated on preparative silica gel columns (Merck silica gel 60, 0.04-0.06 mm) with CH2Cl2-EtOAc 10:1 as eluent. In a typical case, e.g. P. ecklonii, 100 g dry leaf extract yielded 1.1 g of orange powder, which, after separation on the column, yielded 180 mg of (4) and 230 mg of (5). These required one further separation on the Chromatotron (centrifugal TLC) for best purity. All seven compounds isolated in the present investigation are known, and their structures unequivocally determined. Comparison with published 1H and 13C NMR spectra21-24 were used to confirm identities. The X-ray structures of compounds (1), (2) and (3) were elucidated, additionally, to further confirm molecular structural identity.
The two compounds from P. hadiensis, (1) and (2), were first isolated by Eugster and his co-workers21,22 from P. myrianthus (now P. hereroensis) and ascribed the systematic names 7α-formyloxy-6β,12-dihydroxy-abieta-8,12-diene-11,14-dione and 7α-acetoxy-6β,12-dihydroxy-abieta-8,12-diene-11,14-dione, respectively. The two compounds from P. ecklonii (3) and (4) were first isolated in 1978 by Eugster23 from P. parviflorous (now P. strigosus) and designated as parviflorone D and parviflorone F, respectively. Systematically their names are 11-hydroxy-2α-(4-hydroxybenzoyloxy)-abieta-5,7,9(11),13-tetraene-12-one and 11-hydroxy-2α-(3,4-dihydroxybenzoyloxy)abieta-5,7,9(11),13-tetraene-12-one, respectively. From P. lucidus we isolated (3) (above) and (5), 11-hydroxy-19-(methyl-buten-2-oyloxy)-abieta-5,7,9 (11),13-tetraene-12-one.23,24
P. purpuratus subsp. purpuratus afforded (5) and from P. purpuratus subsp. tongaensis (6), 11-hydroxy-19-(4-hydroxy-benzoyloxy)-abieta-5,7,9(11),13-tetraene-12-one, and (7), 11-hydroxy-19-(3,4-dihydroxybenzoyloxy)-abieta-5,7,9(11),13-tetraene-12-one were obtained.23
Antimalarial assay. A chloroquine-resistant strain of Plasmodium falciparum (FCR-3) was continuously maintained in vitro.25 The antimalarial activity of the isolated compounds was determined using the tritiated hypoxanthine incorporation assay.26 Briefly, a parasite suspension, consisting predominantly of the ring stage, was adjusted to 0.5% parasitaemia and 1% haematocrit and plated out with a range of concentrations of the isolated compounds. After 24 h, labelled 3H-hypoxanthine was added and the DNA harvested on glass-fibre filter paper with a semi-automated cell harvester. The concentration that inhibited 50% of the parasite growth (IC50 value) was estimated from the log sigmoid dose-response curve that was generated by the Enzfitter® software.
The antiplasmodial activities of the abietane diterpene and quinine combinations were determined at various concentration ratios based on their individual IC50 values. The concentration range for quinine was between 0.1 and 500 nM, and for the abietane diterpenes between 0.1 and 50 µM. The combination ratio of quinine: abietane diterpenes was arbitrarily set between 0.5: 0.1 and 0.001: 35.26 The IC50 values for the combined drugs were determined using the tritiated hypoxanthine incorporation assay. The nature of the interaction between the two compounds was determined from the isobologram such that points lying above or below the straight diagonal line indicate either an antagonistic or synergistic interaction, respectively. Points lying on the straight line indicate that the combination of the two compounds has an additive interaction.27
The compounds were dissolved in DMSO, which facilitated their dissolution in the aqueous environment. There was no precipitation of the compounds when using 10% DMSO solution; and the final 1% concentration of DMSO was not found to interfere with parasite or cell culture growth.
β-Haematin inhibition assay. A modification of the quantitative β-haematin inhibitory activity (BHIA) assay 28 was used. A 0.767 M haemin solution in DMSO was added to varying concentrations of the test compounds to give final ratios in the range of 1:0.002 to 1:1 (haematin: compound) in triplicate wells; DMSO was used in the control wells. The final percentage of DMSO per well was kept constant at 25%, which does not interfere with the assay. β-Haematin formation was initiated by the addition of 0.5 M acetate buffer (pH 4.4) and the plates were incubated at 37°C for 24 h. After centrifugation at 1750 × g for 20 min, to isolate the DMSO-insoluble β-haematin, it was dissolved in 0.2 M NaOH and diluted fourfold before spectrophotometric absorbance measurement at 405 nm. The haemin: compound molar ratio at which 50% β-haematin formation was inhibited, was calculated from log sigmoidal dose-response curves generated by the Enzfitter® software.
Toxicity analysis. Transformed human kidney epithelial (Graham) cells were maintained continuously in culture at 37°C in 5% CO2. The MTT cellular viability assay was used to determine the toxicity profile of the isolated compounds.29 The trypsinized cell suspension (45 000 cells/well) was plated along with the isolated compounds (in triplicate wells) for 44 h. Thereafter, 20.1 mM MTT was added to the wells and incubated for a further 4 h. DMSO was added to end the reaction and dissolve the formazan crystals. Absorbance was measured at the test wavelength of 540 nm, and at a reference wavelength of 690 nm, and the percentage cellular viability calculated with the appropriate controls taken into account.
Statistical analysis. Statistical calculations were carried out using Graphpad software (San Diego, CA). Results are expressed as the average ± s.d. of at least 3 independent experiments. Student's t-test was used for statistical analysis; P values > 0.95 were taken as significant.

