Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489
Print version ISSN 0038-2353
S. Afr. j. sci. vol.103 n.11-12 Pretoria Nov./Dec. 2007
RESEARCH ARTICLES
The distribution and spread of the invasive alien common myna, Acridotheres tristis L. (Aves: Sturnidae), in southern Africa
Derick S. PeacockI; Berndt J. van RensburgI; Mark P. RobertsonII, *
IDepartment of Zoology and Entomology, Centre for Invasion Biology, University of Pretoria, Pretoria 0002, South Africa
IICentre for Environmental Studies, Department of Zoology and Entomology, University of Pretoria
ABSTRACT
The common myna is an Asian starling that has become established in many parts of the world outside of its native range due to accidental or deliberate introductions by humans. The South African population of this species originated from captive birds that escaped in Durban in 1902. A century later, the common myna has become abundant throughout much of South Africa and is considered to pose a serious threat to indigenous biodiversity. Preliminary observations suggest that the common myna's distribution is closely tied to that of humans, but empirical evidence for this hypothesis is lacking. We have investigated the relationships between common myna distribution, human population size and land-transformation values at a quarter-degree resolution in South Africa. Common mynas were found more frequently than expected by chance in areas with greater human population numbers and land-transformation values. We also investigated the spatial relationship between the bird's range and the locations of South Africa's protected areas at the quarter-degree scale. These results indicate that, although there is some overlap, the common myna distribution is not closely tied to the spatial arrangement of protected areas. We discuss the original introduction, establishment and rate of spread of the common myna in South Africa and neighbouring countries and contrast the current distribution with that presented in The Atlas of Southern African Birds. We also discuss the factors that affect the common myna's success and the consequences that invasion by this species is likely to have, specifically in protected areas.
Introduction
The continually expanding human population causes massive land-use changes1 that result in the alteration of biological communities in terms of species richness, species evenness, guild structuring, biomass and community composition. This, coupled with the increase in international trade and travel, has led to a massive proliferation of alien organisms in the last 40 years.2 Apart from massive economic costs, the adverse environmental effects of biological invasions (the invasion of habitats by non-native organisms), particularly at the local scale, are immense, and alien organisms now constitute a major obstacle for the continued survival of a large proportion of the world's threatened species.1 Indeed, for island ecosystems where alien introduction is particularly prevalent,3 a strong relationship exists between the number of local extinctions and the number of alien invasions.4 Introductions of alien organisms can either be deliberate (e.g. for the pet trade, agriculture, hunting, biological control, habitat restoration, nostalgia) or accidental (e.g. escapees, ship-assisted transportation). In addition, the geographical ranges of many species are naturally increasing as a result of human-induced habitat modification. The deterioration or fragmentation of habitats is not only conducive to alien establishment, but also works synergistically with the negative effects that the alien species themselves pose to remaining indigenous communities.
Apart from the need to understand the processes and patterns of biological invasions in order to minimize their adverse effects, invasions are also of great academic interest because they provide quantifiable, relatively short-term examples of ecological and biogeographical processes. Owing to the large spatial and long temporal scales required, experimental manipulations of populations and communities for the investigation of, for example, range size constraints, competitive exclusion, differential extinction, establishment success, morphological character release, the relationship between community structure and community stability are expensive and time-consuming. However, the modelling of biological invasions provides a more practical evaluation of such processes, and has formed the focus of a multitude of recent ecological studies.3–8
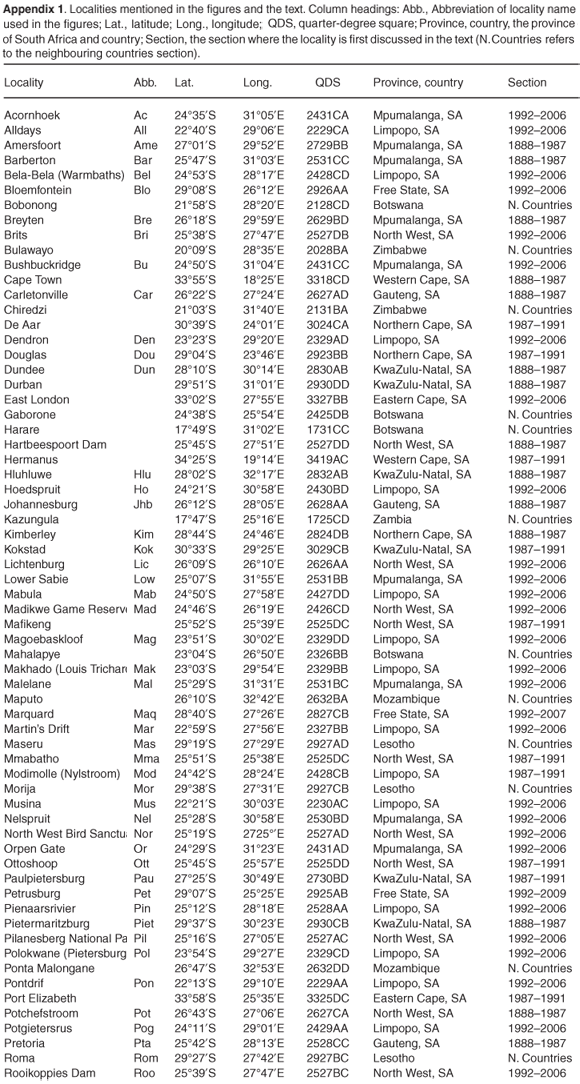
The postulated association between highly successful invasive birds, human distribution and modified habitats in South Africa is exemplified by the invasive alien common myna, Acridotheres tristis, one of only four alien species (out of 48) to have established a viable population and become invasive and widespread.9 Native to Iran, Afghanistan, Turkmenistan, Uzbekistan, Tadzhikistan, southern Kazakhstan, Pakistan, India, Sri Lanka, Nepal, Sikkim, Bhutan, southern China, Burma, Thailand and Vietnam,10 this species has been introduced into many parts of the world outside of its original range (Fig. 1). It now occurs on all continents except South America and Antarctica, and is one of the most common birds on a number of islands in the Indian, Atlantic and Pacific oceans. In addition to deliberate introductions by humans, common mynas are currently also undergoing a natural range expansion at the margins of their native range (e.g. into Singapore, Malaysia and Turkey), probably as a result of extensive clearing of natural vegetation for agriculture11 and increasing human population densities.1
In 2000, the World Conservation Union (IUCN) declared the common myna as one of only three birds among the 'World's 100 worst' invasive species.12 It has been suggested that the common myna poses a threat to indigenous biodiversity through competition for food and nesting resources and aggressive spatial displacement as well as, to a lesser extent, human well-being through the spread of parasites and diseases, agricultural crop damages and noisy and foul communal roosts.10 However, evidence for such effects is generally lacking, particularly in South Africa.
Common mynas are currently undergoing a rapid and extensive range expansion in South Africa. Monitoring the continuing range increase of this species, and determining the relative importance of the influential factors determining its spread and density provides a valuable opportunity for ecologists to manage current, and predict future, avian invasion within the region. In particular, monitoring the common myna's historical and future response to South Africa's large range of climatological and environmental variables, and determining how the localized spatial distribution of major urban centres in South Africa affects its local population densities,13,14 should be prioritized. Ultimately, the results of such studies should be compared with those from other countries suffering similar invasive problems in order to generalize and address the invasion more efficiently.
Throughout its introduced range as well as within its native range,10 the common myna's distribution and population densities seem to be closely tied to that of people. In South Africa, the birds are seldom encountered away from human settlement, even in predominantly natural areas. However, empirical testing and statistical confirmation of this tentative pattern is lacking. In this paper we aim to determine whether the distribution of the common myna is indeed tied to unusually high human population size and other anthropogenic activities such as areas characterized by high land transformation levels. In view of the potential threats that mynas pose to indigenous biodiversity, we also investigate, at the national scale, the relationship between myna distribution patterns and the spatial location of protected areas to understand better the extent to which this species is encroaching upon existing conservation areas. In addition, we provide an updated distribution map of the common myna's range in South Africa, showing the latest published and sight records, and discuss its initial introduction, establishment and spread in South Africa and neighbouring countries.
Methods
Based on the South African Bird Atlas Project (SABAP) data set,15 which captures distributional data for birds (mainly in the period 1987–1991) in 1971 quarter-degree grid (QDG) cells (15' × 15', c. 676 km2), common mynas were found to be present in 256 cells across South Africa. To determine whether cells representing this species are characterized by higher numbers of humans and land transformation than expected by chance, we calculated, at the quarter-degree resolution, the mean human population size (based on the 1996 census data16) and percentage land transformation,13,14 respectively, of these 256 cells. These values were then compared with the mean human population size and percentage land transformation, respectively, found in 10 000 random draws of 256 cells. These grid cells were selected from a pool of all possible grid cells with human population size and land transformation values, i.e. 1971 quarter-degree grid cells covering South Africa. In a similar fashion, we determined whether the cells representing common mynas contained smaller proportions of protected areas than expected by chance.
The 1996, South African population census data supplied by Statistics South Africa (as numbers of humans per quarter-degree grid cell)16 were used for analysis instead of the most recent 2001 population census data17 as the former data set better represents the years during which the SABAP data were compiled. As there is a chance that reporting rates could be higher in areas where human population density is greater, we did not compare the number of individual birds (i.e. common myna abundances) reported per grid cell with human population density. Instead, we selected those grid cells where the species had been observed at least once and compared the population density in these with randomly drawn grid cells. The birds are conspicuous and easily detected due to their distinctive appearance and frequent vocalizations. They are thus likely to have been reported in grid cells where they were present, thus reducing the likelihood of sampling bias. The likelihood of having false negatives, even in areas characterized by relatively low sampling effort, should therefore be low and have little influence on the overall patterns observed.
Data on land transformation were obtained by calculating the percentage of each land-cover class in each quarter-degree cell, based on six transformed land-cover classes provided by Fairbanks et al.14 and Thompson.13 These land-cover values were then summed in each grid cell. These classes were based on seasonally standardized Landsat TM satellite imagery captured primarily during 1994/95 and included cultivated lands, forest plantations, urban/built-up lands, mines and quarries, degraded lands, and water bodies. Finally, the proportion of protected land in each cell was obtained by overlapping the quarter-degree grid with all 637 national level protected areas mapped in the World Database on Protected Areas.18
In order to provide an updated distribution map of the common myna's range in South Africa, dates, locations and details of sight records were collected from a variety of Internet websites (including private birder websites, birding ecotourism sites and online atlassing sites19,20), birding mailing lists and from personal observations. In addition, specific requests for information were posted on South Africa's national birder communication mailing list, SABirdnet (400+ subscribers), and the regionally-based Pretoria BirdNet (380+ subscribers). These records were for the period 1997–2006, spanning almost a decade after the publication of The Atlas of Southern African Birds.15 Sight records were then augmented by the Avian Demography Unit's Birds in Reserves Project data19 and the Pretoria Conservation Challenge electronic bird monitoring data20 and overlaid on the SABAP distributional data.15 Historical records were obtained from the literature, particularly several regional atlases, such as Cyrus & Robson21 and Tarboton et al.22
Results
Association with humans and prevalence in conservation areas
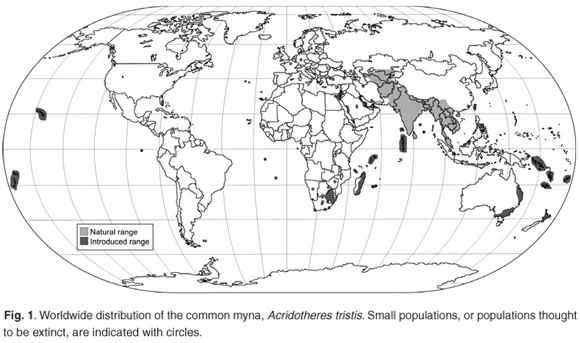
The quarter-degree grid cells representing common myna distribution had significantly greater human population densities and land transformation values, respectively, than expected by chance (P < 0.0001, 10 000 permutations). The total percentage of protected areas contained in the quarter-degree grid cells in which the common myna has been recorded was no different from that of the randomly drawn grid cells (P = 0.155, 10 000 permutations; see also Fig. 2). This indicates that there is some degree of spatial overlap (at this resolution) between the existing protected area network and common myna distribution based on the SABAP data set.15
However, more than a decade has passed since the data on which our analysis is based were collected. During this period, common mynas underwent a considerable range expansion, and it is likely that an analysis of the correlation between human distribution, protected areas and the latest distributional data for this species might yield different results. Given the poor spatial coverage of the latest available distributional data, however, such an analysis is better postponed until a more extensive data set, gathered in a standardized fashion, is available.
Original introductions and early spread (1888–1987)
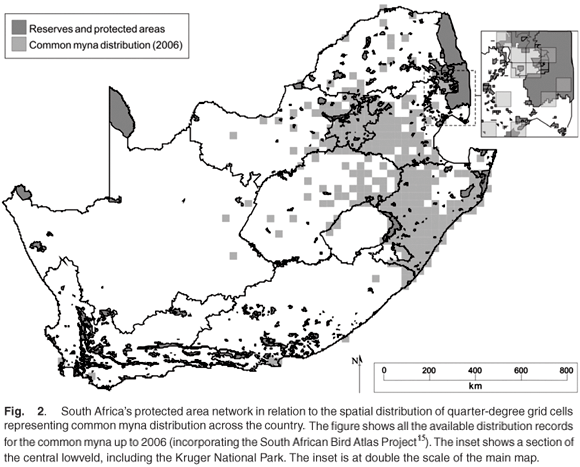
The common myna has been independently introduced into South Africa on at least two, or possibly three, occasions. First, an apparently unsuccessful introduction of birds from Mauritius occurred in Durban, KwaZulu-Natal, in 1888 (Fig. 3).23 This was followed in 1902 by the establishment of escapees from the pet bird trade, primarily of the subspecies A. t. trist oides from Burma and Assam.24 A third introduction, this time of the nominate subspecies, apparently occurred in Johannesburg, Gauteng, in the late 1930s,23 the species first being sighted in Johannesburg's northern suburbs in 1938.25 The closest to Johannesburg that mynas had been recorded at that stage was Dundee, KwaZulu-Natal (approximately 330 km away) and it was thus assumed that the isolated Johannesburg population constituted an independent introduction. This assumption was supported by the fact that the Gauteng birds were of the nominate subspecies A. t. tristis. However, the possibility that a smaller number of the birds introduced to Durban in 1902 were in fact of the nominate subspecies has been raised.26 This, coupled with records of long-distance dispersal of ringed common mynas (e.g. one bird was recovered 381 km from its capture site in little more than a year27) and the uncertain validity of the tristoides subspecies makes the independence of these two introductions doubtful (see Discussion for more details).
The Durban birds reached Pietermaritzburg (some 65 km inland from Durban) in approximately 28 years,23 and continued to invade much of the interior and coastal regions of KwaZulu-Natal in the next 20 years. By 1980, common mynas had invaded as far north as Hluhluwe (Fig. 3), and as far as southern KwaZulu-Natal (where predominantly recorded as a non-breeding visitor23) and westwards over much of the interior regions. Interestingly, they avoided the higher-lying midlands (KwaZulu-Natal) below the Drakensberg escarpment, but extended north-westwards over similar altitudes, perhaps following a major highway linking Gauteng with Durban. The more subtropical northern areas, such as northern KwaZulu-Natal, had not been markedly invaded by 1980.
The spread of mynas in Gauteng (following the postulated 1930s introduction) seems to have progressed at a slower rate than those in KwaZulu-Natal. They were first recorded in Orchards and Bramley in the northern suburbs of Johannesburg in 1938 (where their distribution was estimated to be less than '10 square miles'25). The first sight records in the southern suburbs of Johannesburg (a mere 20 km away) were obtained around 1960. After a considerable initial lag, the birds reached Pretoria (58 km north of Johannesburg) in 1955. However, they did not become widely established there until the 1980s, thus showing a similar lag period to the KwaZulu-Natal populations. Their spread in Pretoria did not seem to follow a linear front or to radiate from a central point, as they were recorded in new locations all around the city (e.g. Hartbeespoort Dam, Pretoria West, Centurion, Swartkops, Wierda Park, East Lynne, Silverton and Hatfield) within six years.25 In the North West province, mynas reached Potchefstroom in 1967 and Rustenburg in 1981. They had also been recorded in Carletonville (Gauteng), Secunda, Breyten, Amersfoort, Volksrust and Wakkerstroom (Mpumalanga) by 1987 and Barberton (the easternmost locality in the former Transvaal province) by 1983.22
The propensity of common mynas to reach isolated locations is also demonstrated by records of breeding attempts in Cape Town in the early 1980s, sight records of single birds near Mahalapye, Botswana in 1975,24 and records of birds breeding, but failing to become established in Kimberley, Northern Cape, in 1975 and 1977. It is unclear, however, if the birds reached Kimberley of their own accord or whether they were introduced.
The Southern African Bird Atlas Project data (1987–1991)
The publication of The Atlas of Southern African Birds15 presented the first detailed map of the entirety of the common myna's distribution range in South Africa (Fig. 4). The 256 quarter-degree grid cells in which the common myna was recorded represents a total of 5.6% of southern Africa's area, and was based on a total of 16 969 records.15 The species was reported on a mean of 42.2% of the atlas cards received for these 256 QDS grids. Despite its distribution being essentially restricted to the eastern half of South Africa and (then) excluding any of the neighbouring countries (Zimbabwe, Botswana, Namibia and Mozambique), the common myna was southern Africa's fourth-most common representative of the 14 southern African starlings (family Sturnidae), being topped by two indigenous species and the alien invasive common starling (Sturnus vulgaris). Out of a list of 48 alien birds introduced into South Africa,9 the common myna is the fourth-most regularly reported species, being eclipsed only by the house sparrow (Passer domesticus) (61 222 records), common starling (21 667 records) and the feral pigeon (Columba livia) (18 449 records).
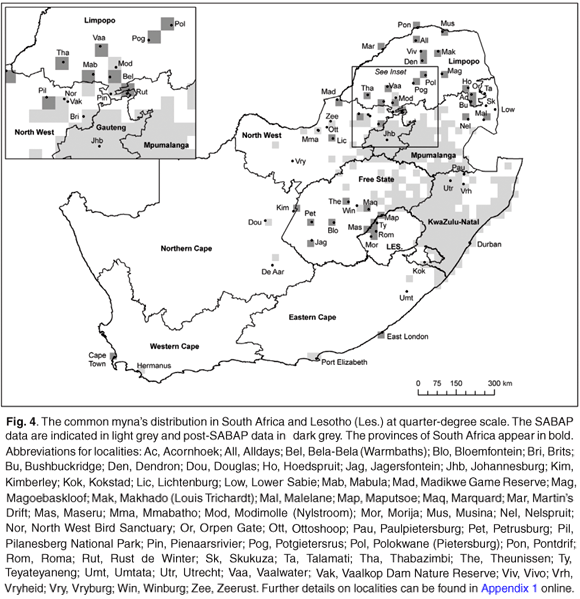
A comparison of the SABAP15 data and maps presented in Cyrus & Robson21 in 1980 reveals little difference in the KwaZulu-Natal distribution, apart from some expansion to the north (now extending to 27°30'S, with individuals recorded on occasion in northern Zululand).15 In addition, higher-lying areas below the escarpment were also invaded, with records right up to the border of Lesotho.15 The notable gap in distribution south of Swaziland during the 1980s21 was also invaded to some extent, the birds having reached Utrecht, Vryheid and Paulpietersburg by 1991.15 There is a remarkably sharp cut-off in the southwestern edge of the KwaZulu-Natal range, along 29°45'E,15 probably reflecting the border of the former Transkei homeland (now the KwaZulu-Natal–Eastern Cape boundary). Apart from a population in Kokstad and an isolated population north of Umtata, the birds were not recorded southwest of this line, and first reappear in Port Elizabeth (approximately 540 km southwest, as the myna flies).15 The only record for the Western Cape province was obtained on the coast near Hermanus.15 This population is at least 590 km from the nearest isolated population (Port Elizabeth) and approximately 1090 km from the nearest edge of the main distribution (southern KwaZulu-Natal).
In the former Transvaal province (Gauteng, Mpumalanga, Limpopo and the eastern North West), a more dramatic range expansion is revealed in the SABAP data (as compared with Tarboton et al.22). In particular, common mynas greatly expanded their range on an east–west axis, showing an essentially continuous distribution from 26°E (Zeerust, Ottoshoop) eastwards to the border of Swaziland. On a north–south axis, common mynas then occurred in continuous band from the northern Free State (27°S) to about 25°S, with the northernmost records obtained at Modimolle (Nylstroom). However, the birds were largely absent from the Limpopo province at that time. Their distribution also extended patchily across much of the northeastern and northern Free State, reaching Winburg in the southwest. Visual inspection of the SABAP map shows an interesting zone of reduced reporting rates in the eastern Free State, southern Mpumalanga and north-western KwaZulu-Natal, which might correspond to a gap between populations derived from the Gauteng and KwaZulu-Natal introductions (Fig. 4). Small outlying populations of common mynas were also recorded in the North West province, e.g. near Mmabatho/Mafikeng and northwest of Vryburg, and in the Northern Cape, e.g. Douglas and north of De Aar.
Post-SABAP spread (1992–2006)
The continuing range expansion of the common myna in South Africa is poorly documented for the 15 years after data gathering for SABAP ended in December 1991. However, a large number of new records have been obtained (see Fig. 4 and Appendix 1 in supplementary material online at www.sajs.co.za) through the advent of birdwatching, e-mail newsgroups, particularly the national SABirdnet. During this period, mynas considerably expanded their range northwards into the Limpopo province and northeastwards into the lowveld of Limpopo and Mpumalanga provinces. In Gauteng, the common myna is now recorded from all the quarter-degree grid cells. In the North West province, common mynas are now well-established in several major urban centres (e.g. Lichtenberg, Brits) and also occur in, or in close proximity to, several conservation areas, such as Vaalkop Dam Nature Reserve, North West Bird Sanctuary, Pilanesberg National Park, Rooikoppies Dam Nature Reserve (near Brits) and Madikwe Game Reserve on the Botswana border.
Records of the common myna in the Limpopo province (in addition to the SABAP data) now exist for Vaalwater, Bela-Bela (Warmbaths), Mabula, Pienaarsrivier, Rust de Winter, Potgietersrus, Thabazimbi, Dendron, Vivo and Alldays (Fig. 4). It is also regularly reported on the Botswana border, e.g. the Pontdrif and Martin's Drift border posts on the Limpopo River. It is probably reasonable to assume that common mynas also occur around human habitation (albeit at low densities or in small, localized populations) in the intervening areas as well. The birds were first recorded breeding in Polokwane (Pietersburg) in 1997,24 and are also recorded from the nearby Magoebaskloof area. From there they extended northwards (probably along the N1 highway), becoming established at Makhado (Louis Trichardt) and reached the border of Zimbabwe at Musina in approximately July 2004.
Many urban centres in the lower-lying areas of Mpumalanga have also been invaded in recent years. Common mynas now seem to be resident in Hoedspruit, Acornhoek, Bushbuckridge, Nelspruit and Malelane, for example, but the extensive Kruger National Park remains largely uninvaded.28
In the Northern Cape, the common myna was first found breeding in Kimberley in 1975 but subsequently disappeared for almost three decades. However, reports of a pair of birds (possibly more) in Kimberley have been received on a regular basis since 2003. Bloemfontein is another large urban centre that is currently being colonized. The common myna has been reported with increasing frequency in Bloemfontein since 1999.27,30 By mid-2006 their numbers had risen to 37 birds observed near a presumed roost site (R.J. Nuttall, pers. obs.). Furthermore, the species seems to be well-established in Marquard (125 km northeast of Bloemfontein30) and isolated sight records exist from Theunissen (February 200131), Petrusburg (November 2001) and towards Jagersfontein (November 2001; R.J. Nuttall, pers. comm.). A small population was also recently discovered in Woodstock, Cape Town, and the bird now seems to be resident at two major cities in the Eastern Cape: Port Elizabeth (up to 10 birds reported in 2003) and East London (a pair, or rarely three birds first recorded in 2001). The remainder of post-SABAP records indicates marginal range expansion on the edge of the known distribution (e.g. Central Free State, Northern Cape and North West province).
Neighbouring countries
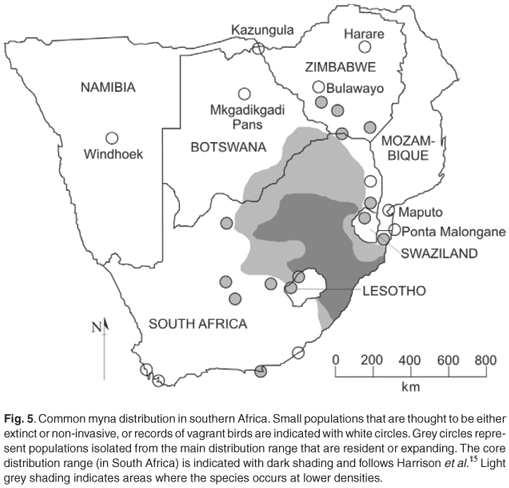
Common mynas were first recorded in Botswana in 1975 near Mahalapye,24 some 60 km from the South African border. Reported in Gaborone in 1991,24 and now common there and in many of the villages in the southeast of the country (e.g. Selebi-Phikwe, Bobonong), especially along major roads (C. Brewster, pers. comm.). They are also established at Sherwood Ranch, 10 km from the Martin's Drift border post (where regularly reported). Sighting was reported from a tourist camp on the edge of the isolated Makgadikgadi Pans National Park in 2005, and recently from Kazungula on the Botswana– Zimbabwe border. In Namibia, it was reported from Windhoek in 2004. In Zimbabwe, it was reportedly 'becoming established' in Harare and Bulawayo in the early 1980s,32 although these occurrences were not reflected in the SABAP map.15 Further records from Zimbabwe (e.g. West Nicholson, Colleen Bawn, near Chiredzi; G. Lowe, pers. comm.) suggest a progressive infiltration of that country from the south, where small numbers of mynas occur in neighbouring northern South Africa and Botswana. In Mozambique, sighting was first recorded in 1997 at Ponta Malongane, some 10 km north of the KwaZulu-Natal border, where a group of six birds was seen around human habitation.33 Sighting was also reported from Maputo in 2005, where much suitable habitat exists and colonization is likely to occur within the next decade. Although mynas are to date completely absent from the highlands of Lesotho, suitable habitat exists in the western lowlands of that country, and large-scale colonization of Maseru is highly likely. The species was first recorded in Maseru in January 1994 (first record for Lesotho), and has now been reported in small numbers from Maputsoe, Roma, Morija and Teyateyaneng34 but is undoubtedly more widespread than currently realized in Lesotho. It is reported to be resident at a single locality in western Swaziland,24 but it is apparently spreading there (see also Fig. 5).
Discussion
Historical distribution and invasion rate
The common myna has been deliberately introduced into many areas outside of its native range as a potential biological control agent of insects, particularly locusts and grasshoppers (the generic name Acridotheres literally means 'grasshopper-hunter'35). Conversely, in terms of accidental introductions, mynas' pleasant, varied and melodic song, ability to mimic human speech, intelligence and attractive plumage make them popular cage birds and a large proportion of their current non-native range was initially colonized by escapees.10 The latter case is also applicable to the South African population (or at least, the Durban introduction24).
Claims that the common myna was independently introduced into Durban (1902) and Johannesburg (1930s) are unsubstantiated. The paler ventral surfaces and less reddish dorsal plumage colouration of the Gauteng birds suggests that northern South Africa is predominantly inhabited by the nominate tristis subspecies, whereas the southern and eastern birds are of the subspecies tristoides.26 However, disagreement exists in the literature about whether the escaped birds in Durban were exclusively tristoides24,26 or if both subspecies were present.32 Furthermore, the subspecific validity of tristoides is not widely recognized (e.g. omitted from Feare & Craig10), and the morphological differences between these two taxa are at best subtle, and possibly clinal. The assumption that the two introductions were independent was apparently largely based on the absence of common mynas between Johannesburg and Dundee (approximately 330 km) at the time. Given that a ringed common myna recently recovered in Groenkloof, Pretoria, moved 381 km in little more than one year,27 this criterion alone cannot be viewed as conclusive evidence for an independent introduction. As has been demonstrated for another invasive alien bird species in southern Africa, the house sparrow (Passer domesticus),36 it is possible that occasional long-distance dispersal (so-called 'leap-frogging') occurs in the common myna.
As has been found in Australia,37 the population size and range of the common myna increased slowly after the initial introduction,9 possibly as a result of a small founder population5 and the Allee effect.36 In the latter case, the initial spread of an introduced organism is initially retarded due to a small population size, reduced fecundity and minimal juvenile dispersal. Once the population increases above a specified threshold level, a major acceleration in the population growth rate, and consequently a rapid range expansion, occurs. The duration of this initial lag period, potentially affected by factors such as propagule size and activity levels of people,38 is expected to vary between regions, as common mynas in Israel took only 'a few years' to start rapidly expanding their range.39
Factors affecting common mynas' success in South Africa
Duncan et al.5 showed that most highly successful introduced alien birds are closely associated with human habitation and modified habitats. For example, of 48 alien bird species introduced to South Africa, only seven have established viable populations and only four of these, all commensals of humans and one of which is the common myna, are considered to be widespread.9 Our study clearly supported this close association with urban habitats at the national scale, the latter areas being characterized by high human population numbers that is indicative of the common myna's distribution range. The significant spatial congruence between high levels of land transformation and myna distribution patterns also comes as no surprise with (i) human population density showing a strong positive relationship with land transformation patterns across South Africa,40 and (ii) most highly transformed parts of South Africa represent urban/built-up areas and permanent cultivated lands,13,14 both being highly suitable habitat conditions for the common myna.10,24 Indeed, at least for Israel, it has been suggested that agricultural fields may act as stepping-stones for mynas to expand their distribution range from one major urban area to another (S. Kark, pers. comm.). Furthermore, its large native range spans a variety of climatic and topographic zones,41 which has also been implicated in its success as an invader throughout the world. However, Brooke et al.23 suggested that cold temperatures in the arid interior of South Africa might arrest the common myna's spread southwestwards of its current distribution. Global climate change could also result in a southward shift of the current range.9
The success of invasive common mynas in South Africa is further explained by a number of biological, physiological and ecological characters. It meets most of the criteria given for successful invasion by Dean:9 the common myna was repeatedly introduced, is strongly associated with humans, is omnivorous and opportunistic, often roosts and feeds in flocks, can utilize alien vegetation and has a wide climatic tolerance.10,23,26,42 In terms of its breeding biology, more beneficial characters exhibited by the common myna include solitary nesting, a high average clutch size, and adaptability in regard to choice of nest site.43 Most of these characteristics are shared by the common starling, another invasive alien sturnid in South Africa that has dramatically increased its range in the last century.44
Consequences for indigenous biodiversity and humans
Despite severe public persecution, the adverse effects of introduced common mynas on indigenous biodiversity in South Africa have not been well documented, and most descriptions are of an anecdotal nature (but see Birkhead45). Elsewhere, mynas are often criticized for competing with indigenous birds46 and mammals47 for nesting cavities, which has been shown to act as a limiting resource for many species throughout the world.48 Predation of chicks and eggs, and direct attacks on adults of indigenous and other alien birds, have been reported in some areas, particularly island ecosystems.10 Interspecific aggression between common mynas and indigenous urban birds (e.g. laughing dove (Streptopelia senegalensis), crested barbet (Trachyphonus vaillantii), red-throated wryneck (Jynx ruficollis) Karoo thrush Turdus smithi), Cape sparrow (Passer melanurus), and southern masked-weaver (Ploceus velatus); R. Geyser, K. Hattingh, pers. comm.; D.S.P., pers. obs.) are occasionally witnessed, but potential displacement of indigenous species is believed to be limited to the local scale.26 The largest proportion of the common myna's diet consists of insects and other invertebrates; ironically, mynas are now believed to destroy beneficial insects in some of the areas where they were originally introduced to reduce the numbers of pest insects.10
When insects are scarce, fruits and seeds make up a more important component of their diet. At such times, common mynas can become agricultural pests.41 Mynas have also been implicated in the spread of invasive alien plants in some parts of the world (e.g. Lantana camara in Hawaii2) by acting as seed dispersal agents. Furthermore, mynas can potentially contribute to the spread of parasites (e.g. the mite Ornithonyssus bursa, which can cause dermatitis in humans47) and diseases (e.g. avian malaria Plasmodium circumflexum10), especially in areas where they congregate in close proximity to humans, e.g. communal roost sites. Such communal roosts (which can be up to several thousand individuals strong) are also exceedingly noisy, cause damage to trees by limb-breaking and accumulation of droppings, and have unpleasant odours.42
If the common myna is indeed a threat to biodiversity conservation in South Africa, its presence in conservation areas is of particular concern. Our results indicate that, although myna distribution is not explicitly tied to the distribution of protected areas, the species does not avoid such areas (see Fig. 2). Moreover, this result is probably quite conservative, seeing that our investigations were based on a scale that is probably too coarse to identify finer-scale patterns (see also ref. 49). Unfortunately, our study was limited to a relatively coarse scale (15' × 15' grid cells) as this was the scale at which the data for the Southern African Bird Atlas Project15 were collected. It is likely that a finer-resolution study, revealing finer-scale effects, will show a higher degree of overlap between myna distribution and protected areas. Indeed, mynas are conspicuous members of the avian communities of many of the smaller, more isolated reserves that are distributed throughout South Africa's moister eastern regions. It is likely, given the common mynas' close association with people, that its presence in protected areas is in part an indirect result of human distribution encroaching onto such sites.40
The common myna's range expansion has been particularly rapid and extensive in South Africa's Limpopo province, and it now occurs in many urban centres adjacent to the extensive Kruger National Park for example (see Fig. 2 inset). Common mynas have been recorded in this reserve on occasion (e.g. at Lower Sabie, November 2000; Talamati camp, January 200128) but these individuals have been controlled or have disappeared soon after first being noticed.28 The park has therefore not been invaded to any considerable extent (at the time of writing). In general, most of the sightings in the KNP are singletons or pairs of birds and have been made mainly along the western border of the park (L.C. Foxcraft, pers. comm.) and one can therefore assume that these individuals are always within a few minutes' flight of some form of human habitation outside the reserve. Nevertheless, at least some suitable habitat exists in the park, particularly in the larger tourist camps (e.g. Skukuza) and the common myna's presence on or just outside the park's borders (e.g. Sabie Sand Game Reserve, Orpen Gate area, Malelane Gate) has prompted the KNP's Alien Biota Section to issue a biohazard warning for the species in the park (L.C. Foxcroft, pers. comm.).
Conclusions
The continuing range expansion of the common myna in South Africa is a popular but contentious topic of debate among ecologists, birders and the general public. It is surprising, however, given the bird's abundance in thousands of suburban gardens throughout the country that very few empirical studies have quantitatively assessed the effects that increasing myna populations are having on indigenous birds. In particular, it is widely believed that they compete with, and eventually displace local birds, but evidence for this is lacking in the literature. Birders should be encouraged to keep notes on any such interspecific interactions and the introduction and spread of alien species in general. From a biogeographical viewpoint at least, the common myna is a fascinating and relevant study subject. We hope that this paper will stimulate further research (from both an ecological and conservation perspective) on invasive species and the common myna in particular.
We would like to thank the all the birders around the country who allowed us a peek into their notebooks: Geoff Lowe, Shaun Peard, Rihann Geyser, Chris Brewster, Pauline Leinberger, Greg Lock, Marius Wheeler, Chris Roche, Les Underhill, Mark Anderson, Peter Oosthuizen, John van Zyl, Etienne Marais, Greg Davies, Hans Schouwink, Oscan van Eyssen, Anthony Cooper, Lester Niss, Mike Bridgeford, Johan Snyman, Stuart Groom, Ross Wanless, David Weaver, Rob Webster, Richard Hurt, Giles Mulholland, Neil Whyte, Richard Rae, Adam Welz, John Bradshaw, Phil Penlington, John McAllister, Kobus Pienaar, Tony Archer, Richard Hurt, Rolf Becker, Ursula Franke and Phil Whittington. B.J.V.R. and D.S.P. acknowledge support from the DST-NRF Centre of Excellence for Invasion Biology.
1. Sala O.E., Chapin F.S., Armesto J.J., Berlow E., Bloomfield J., Dirzo R., Huber-Sanwald E., Huenneke L.F., Jackson B., Kinzig A., Leemans R., Lodge D.M., Mooney H.A., Oesterheld M., Poff N.L., Sykes M.T., Walker B.H., Walker M. and Wall D.H. (2000). Biodiversity—Global biodiversity scenarios for the year 2100. Science 287, 1770–1774. [ Links ]
2. Pimentel D., Lach L., Zuniga R. and Morrison D. (2000). Environmental and economic costs of nonindigenous species in the United States. BioScience 50, 53–65. [ Links ]
3. Blackburn T.M. and Duncan R.P. (2001). Establishment patters of exotic birds are constrained by non-random patterns in introduction. J. Biogeogr. 28, 927–939. [ Links ]
4. Case T.J. (1996). Global patterns in the establishment and distribution of exotic birds. Biol. Conserv. 78, 69–96. [ Links ]
5. Duncan R.P., Blackburn T.M. and Sol D. (2003). The ecology of bird introductions. Annu. Rev. Ecol. Syst. 34, 71–98. [ Links ]
6. Cassey P., Blackburn T.M., Jones K.E. and Lockwood J.L. (2004). Mistakes in the analysis of exotic species establishment: source pool designation and correlates of introduction success among parrots (Aves: Psittaciformes) of the world. J. Biogeogr. 31, 277–284. [ Links ]
7. Cassey P., Blackburn T.M., Russell G.J., Jones K.E. and Lockwood J.L. (2004). Influences on the transport and establishment of exotic bird species: an analysis of the parrots (Psittaciformes) of the world. Glob. Change Biol. 10, 417–426. [ Links ]
8. Cassey P., Blackburn T.M., Duncan R.P. and Lockwood J.L. (2005). Lessons from the establishment of exotic species: a meta-analytical case study using birds. J. Anim. Ecol. 74, 250–258. [ Links ]
9. Dean W.R.J. (2000). Alien birds in southern Africa: what factors determine their success? S. Afr. J. Sci. 96, 9–14. [ Links ]
10. Feare C. and Craig A. (1998). In Starlings and Mynas, pp. 157–161. Christoper Helm, London. [ Links ]
11. Lim H.C., Sodhi N.S., Brook B.W. and Soh M.C.K. (2003). Undesirable aliens: factors determining the distribution of three invasive bird species in Singapore. J. Trop. Ecol. 19, 685–695. [ Links ]
12. Lowe S., Browne M., Boudjelas S. and de Poorter M. (2000). 100 of the World's Worst Invasive Alien Species. A selection from the Global Invasive Species Database. The Invasive Species Specialist Group (ISSG), a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN), Auckland. [ Links ]
13. Thompson M. (1996). The standard land-cover classification scheme for remote-sensing application in South Africa. S. Afr. J. Sci. 92, 34–42. [ Links ]
14. Fairbanks D.H.K., Thompson M.W., Vink D.E., Newby T., van den Berg H.M. and Everard D.A. (2000). The South African land-cover characteristics database: a synopsis of the landscape. S. Afr. J. Sci. 96, 69–86. [ Links ]
15. Harrison J.A., Allan D.G., Underhill L.G., Herremans M., Tree A.J., Parker V. and Brown C.J. (eds). (1997). The Atlas of Southern African Birds. BirdLife South Africa, Johannesburg. [ Links ]
16. Anon. (1996). Population Census 1996. Statistics South Africa, Pretoria. [ Links ]
17. Anon. (2001). Census 2001: Key Results. Statistics South Africa, Pretoria. [ Links ]
18. WDPA (2004). World database on protected areas. IUCN-WCPA and UNEP-WCMC, Washington, D.C. [ Links ]
19. BIRP (2005). Birds in Reserves Project. Online: www.birds.sanbi.org [ Links ]
20. PCC. (2005). Pretoria Conservation Challenge. Online:www.birding.co.za/ challenge [ Links ]
21. Cyrus D. and Robson N. (1980). Bird Atlas of Natal. University of Natal Press, Pietermaritzburg. [ Links ]
22. Tarboton W.R., Kemp M.J. and Kemp A.C. (1987). Birds of the Transvaal. Transvaal Museum, Pretoria. [ Links ]
23. Brooke R.K., Lloyd P.H. and De Villiers A.L. (1986). Alien and translocated terrestrial vertebrates in South Africa. In The Ecology and Management of Biological Invasions in Southern Africa, eds I.A.W. MacDonald, F.J. Kruger and A.A. Ferrar, pp. 63–74. Oxford University Press, Cape Town. [ Links ]
24. Craig A.J.F.K. (1997). Indian myna Acridotheres tristis. In The Atlas of Southern African Birds, vol. 2., eds J.A. Harrison, D.G. Allan, L.G. Underhill, M. Herremans, A.J. Tree, V. Parker and C.J. Brown, pp. 454–455. BirdLife South Africa, Johannesburg. [ Links ]
25. Carr A. (1988). The Indian myna—colonisation of Pretoria. Laniarius 32, 10–14. [ Links ]
26. Hockey P.A.R, Dean W.R.J. and Ryan P.G. (eds) (2005). Roberts – Birds of Southern Africa, 7th edn. The Trustees of the John Voelcker Bird Book Fund, Cape Town. [ Links ]
27. Oschadleus H.D. (2001). Report on recoveries received at SAFRING: July 2000—June 2001. Afring News 30, 74–81. [ Links ]
28. Foxcraft L.C. and Richardson D.M. (2003). Managing alien invasions in the Kruger National Park, South Africa. In Plant Invasions: Ecological Threats and Management Solutions, eds L.E. Child, J.H. Brock, G. Brundu, K. Prach, P.M. PyÓek and M. Williamson, pp. 385–403. Backhuys Publishers, the Netherlands. [ Links ]
29. Nuttall R.J. (1999). Sightings of interest: Free State Region. Mirafra 16(2), 27–28. [ Links ]
30. Colahan B.D. (2000). The European starling in central Bloemfontein—has the invasion begun? Mirafra 17(1&2), 10–11. [ Links ]
31. Nuttall R.J. (2001). Sightings of interest: Free State Region. Mirafra 18(1), 11–15. [ Links ]
32. Fry C.H., Keith S. and Urban E.K. (eds) (2000). The Birds of Africa, vol. 6, pp. 660–661. Academic Press, London. [ Links ]
33. Parker V. (1999). The Atlas of the Birds of Sul do Save, Southern Mozambique, p. 224. Avian Demography Unit and Endangered Wildlife Trust, Cape Town and Johannesburg. [ Links ]
34. Ambrose D. (2002). Sightings of interest in Lesotho: July 2000–June 2002. Mirafra 19(4), 53–56. [ Links ]
35. Clinning C. (1989). Southern African Bird Names Explained, p. 2. The Southern African Ornithological Society, Johannesburg. [ Links ]
36. Msimanga A. and Slotow R. (2000). The house sparrow Passer domesticus (Passeriformes: Ploceidae) invasion of southern Africa: rates and the Allee effect. Durban Mus. Novit. 25, 5–11. [ Links ]
37. Hone J. (1978). Introduction and spread of the common myna in New South Wales. Emu 78, 227–230. [ Links ]
38. Williamson M.H. and Fitter A. (1996). The characters of successful invaders. Biol. Conserv. 78, 163–170. [ Links ]
39. Holzapfel C., Levin N., Hatzofe O. and Kark, S. (2006). Colonisation of the Middle East by the invasive common myna (Acridotheres tristis, L.), with special reference to Israel. Sandgrouse 28, 44–51. [ Links ]
40. Chown S.L., van Rensburg B.J., Gaston K.J., Rodrigues A.S.L. and van Jaarsveld A.S. (2003). Energy, species richness, and human population size: conservation implications at a national scale. Ecol. Appl. 13, 1233–1241. [ Links ]
41. Martin W.K. (1996). The current and potential distribution of the common myna Acridotheres tristis in Australia. Emu 96, 166–173. [ Links ]
42. Yap C.A.M., Sodhi N.S. and Brook B.W. (2002). Roost characteristics of invasive mynas in Singapore. J. Wildl. Manage. 66, 1118–1127. [ Links ]
43. Tarboton W.R. (2001). A Guide to the Nests and Eggs of Southern African Birds. Struik, Cape Town. [ Links ]
44. Richardson D.M. (1992). Starlings: why have they been so successful? African Wildlife 46, 207–210. [ Links ]
45. Birkhead J. (1988). Indian mynas oust redbilled woodhoopoes and other hole nesters. Laniarius 33, 8–9. [ Links ]
46. Pell A.S. and Tidemann C.R. (1997). The impact of two exotic hollow-nesting birds on two native parrots in savannah and woodland in eastern Australia. Biol. Conserv. 79, 145–153. [ Links ]
47. Central Coast Indian Myna Action Group (2003). Research report: impact of the common (Indian) myna on health and the environment and recommendations of the implementation of a control program. Central Coast Indian Myna Action Group, Gosford, Australia. [ Links ]
48. Newton I. (1994). The role of nest sites in limiting the numbers of hole-nesting birds: a review. Biol. Conserv. 70, 265–276. [ Links ]
49. Van Rensburg B.J., Koleff P., Gaston K.J. and Chown S.L. (2004). Spatial congruence of ecological transition at the regional scale in South Africa. J. Biogeogr. 31, 843–854. [ Links ]
Received 10 July 2006. Accepted 5 September 2007.
* Author for correspondence. E-mail: mrobertson@zoology.up.ac.za
Supplementary material to:
Peacock D.S., van Rensburg B. J. and Robertson M. P. (2007). The distribution and spread of the invasive alien common myna, Acridotheres tristis, L. (Aves: Sturnidae), in southern Africa. S. Afr. J. Sci. 103, 465–473.